Phytoremediation Efficacy of Salix discolor and S. eriocephela on Adjacent Acidic Clay and Shale Overburden on a Former Mine Site: Growth, Soil, and Foliage Traits

Abstract
:1. Introduction
2. Materials and Methods
Statistical Analysis
3. Results
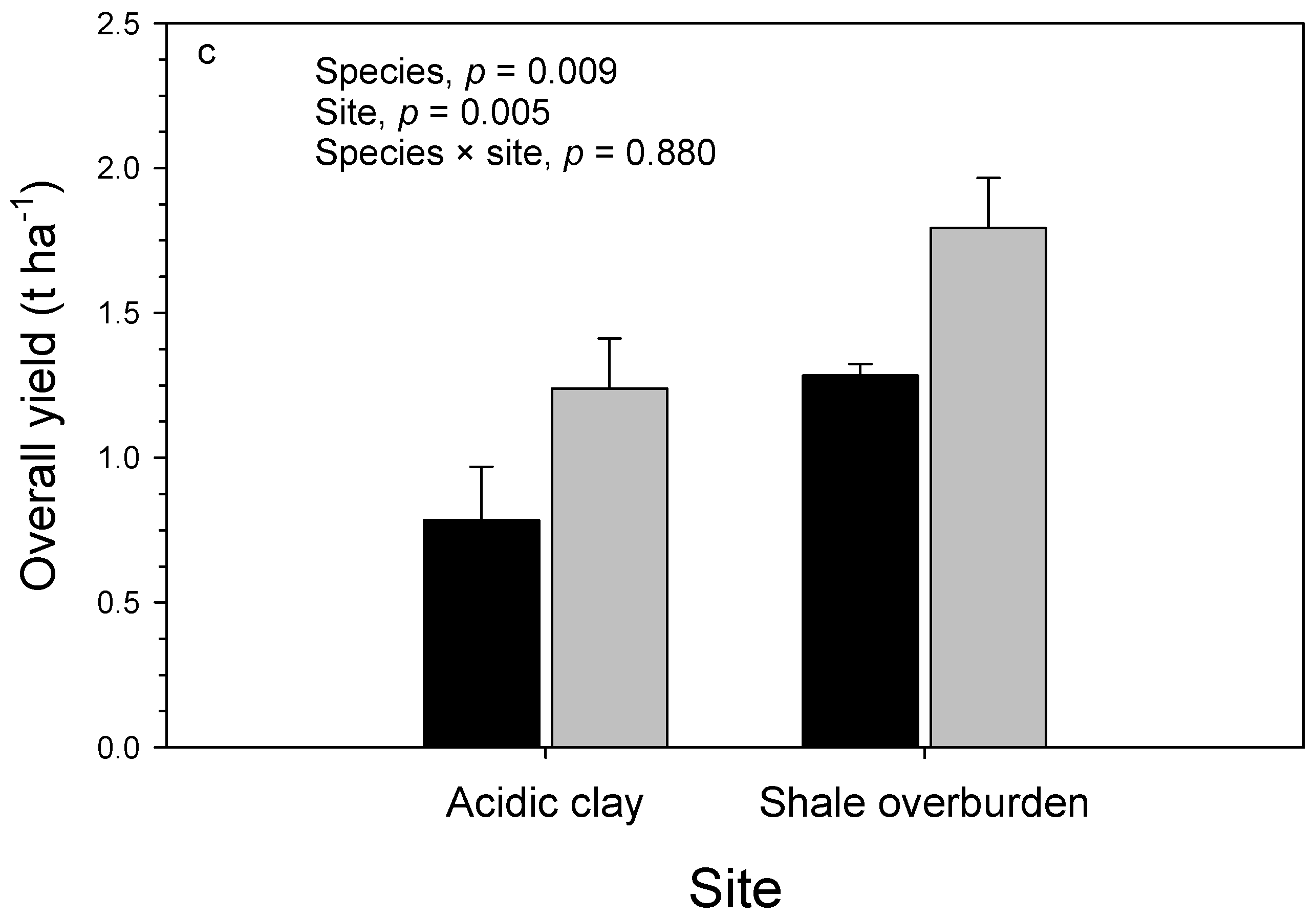
3.1. Growth
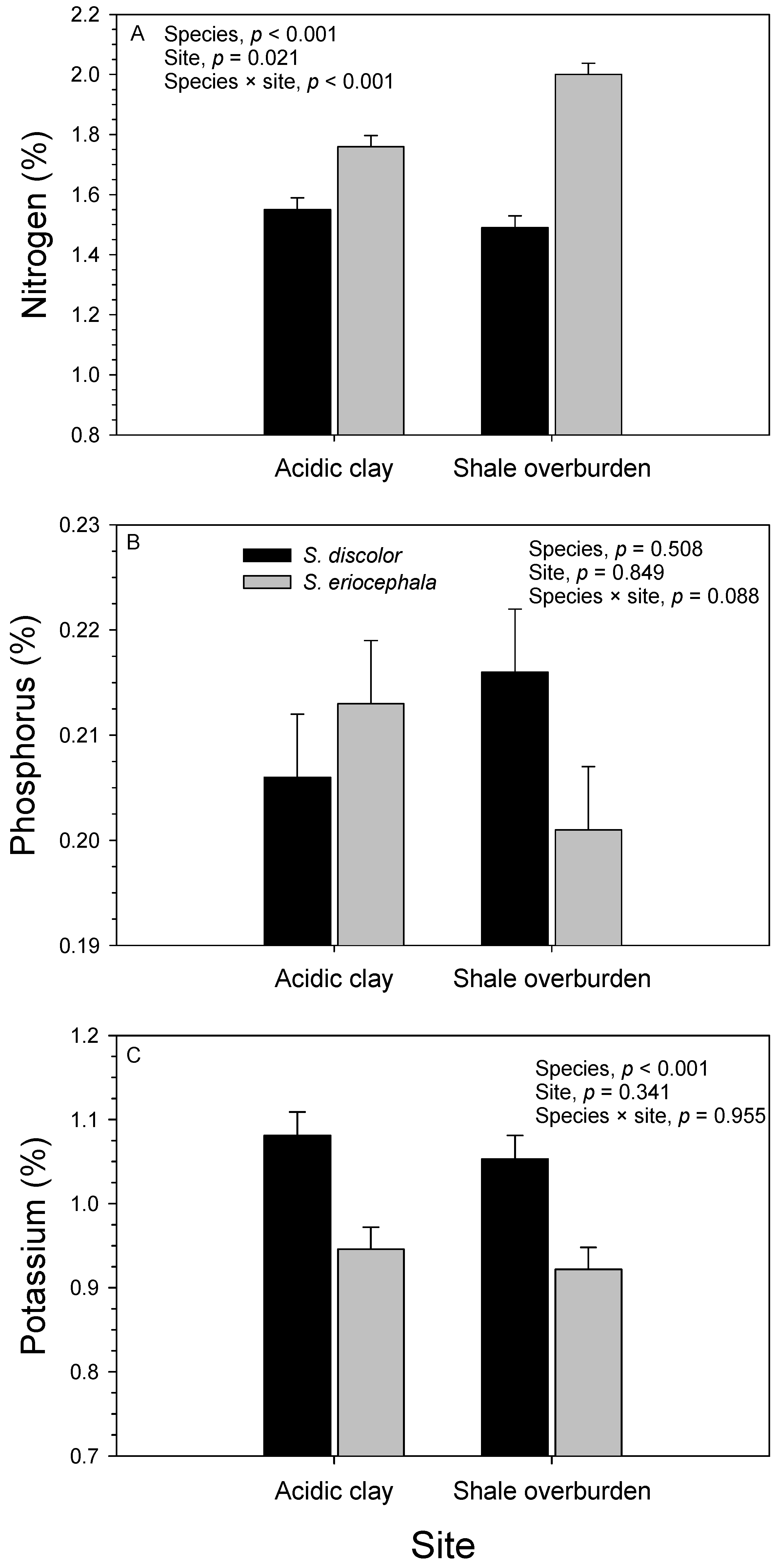
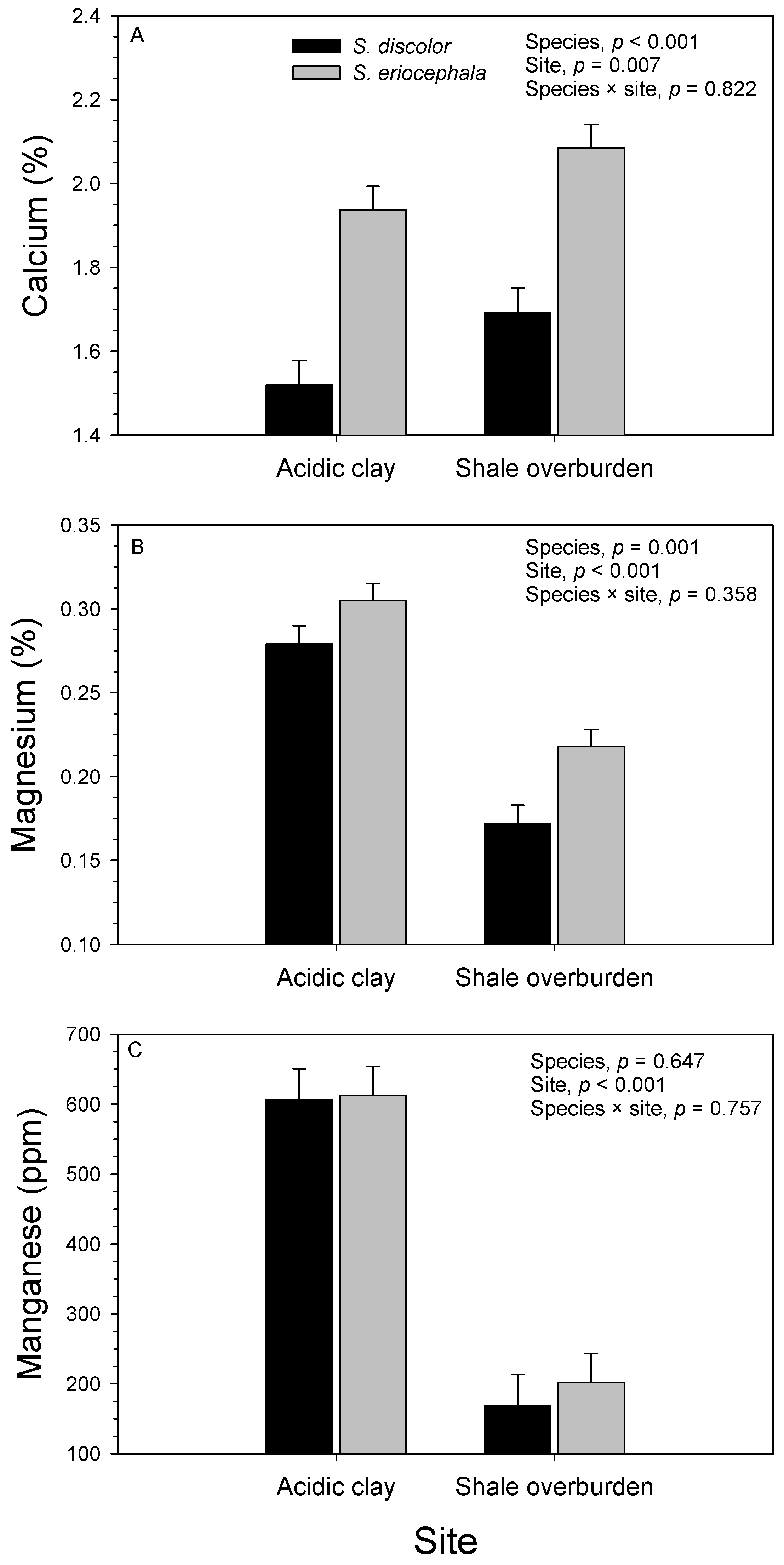
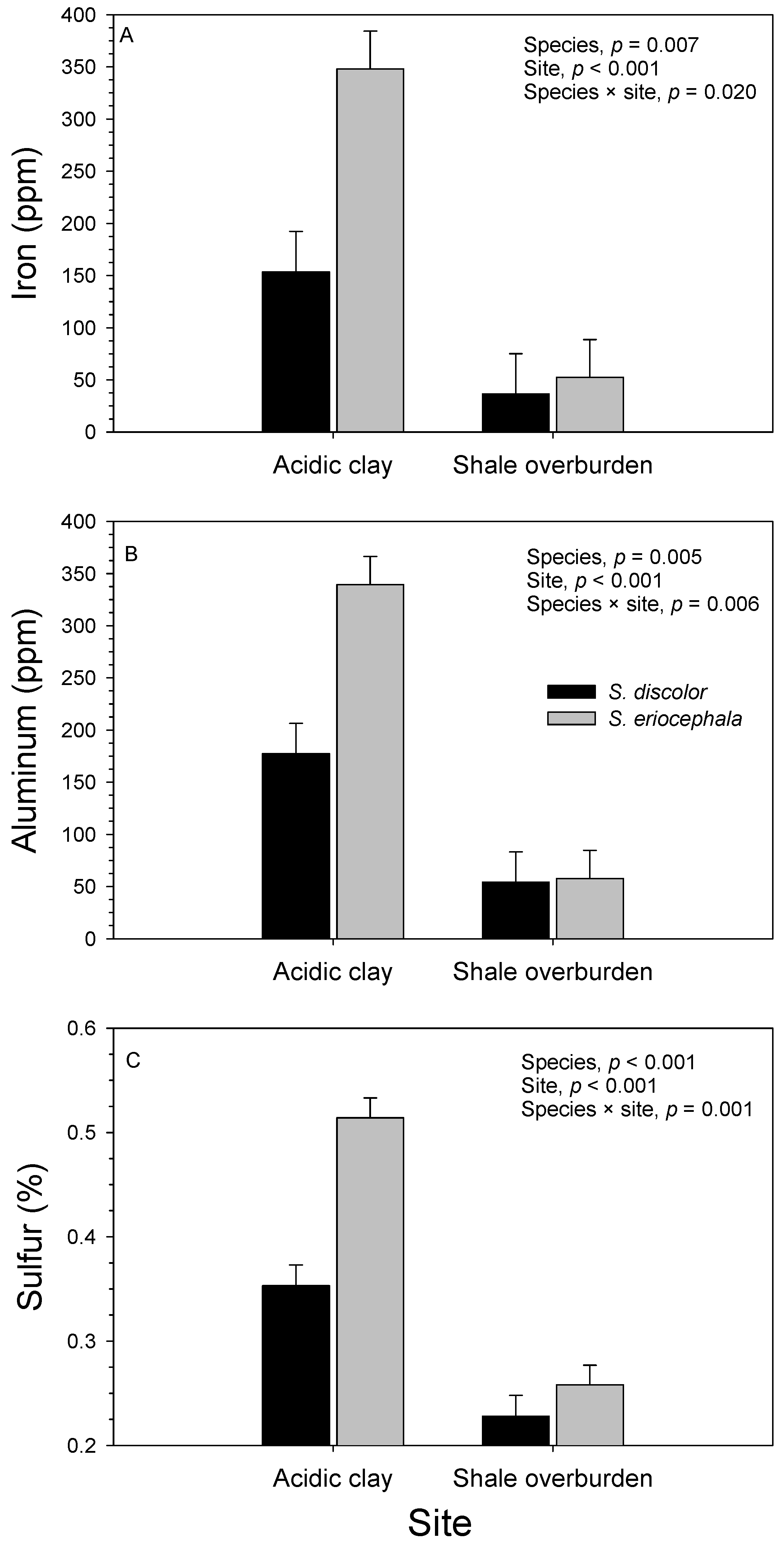
3.2. Soil
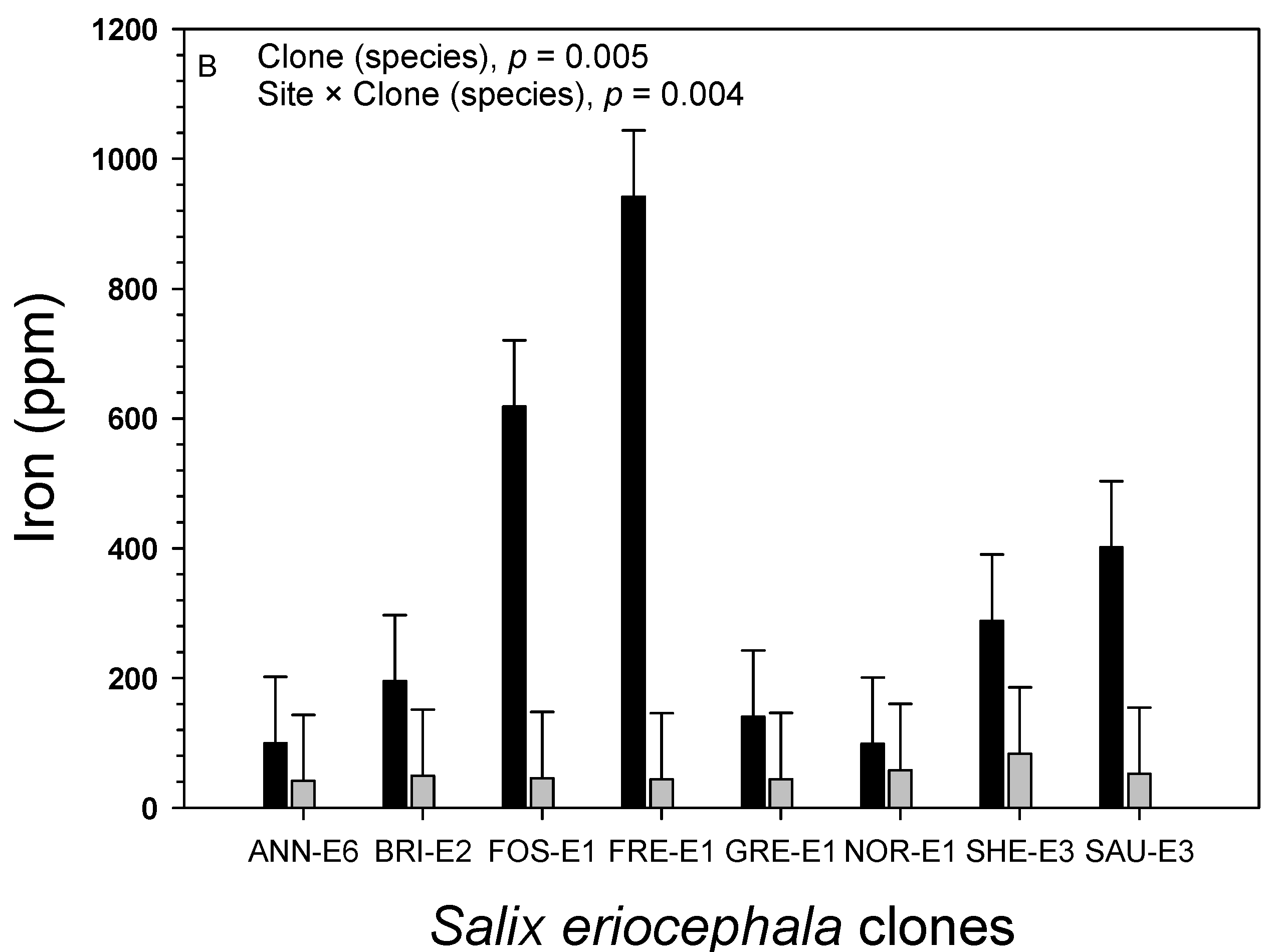
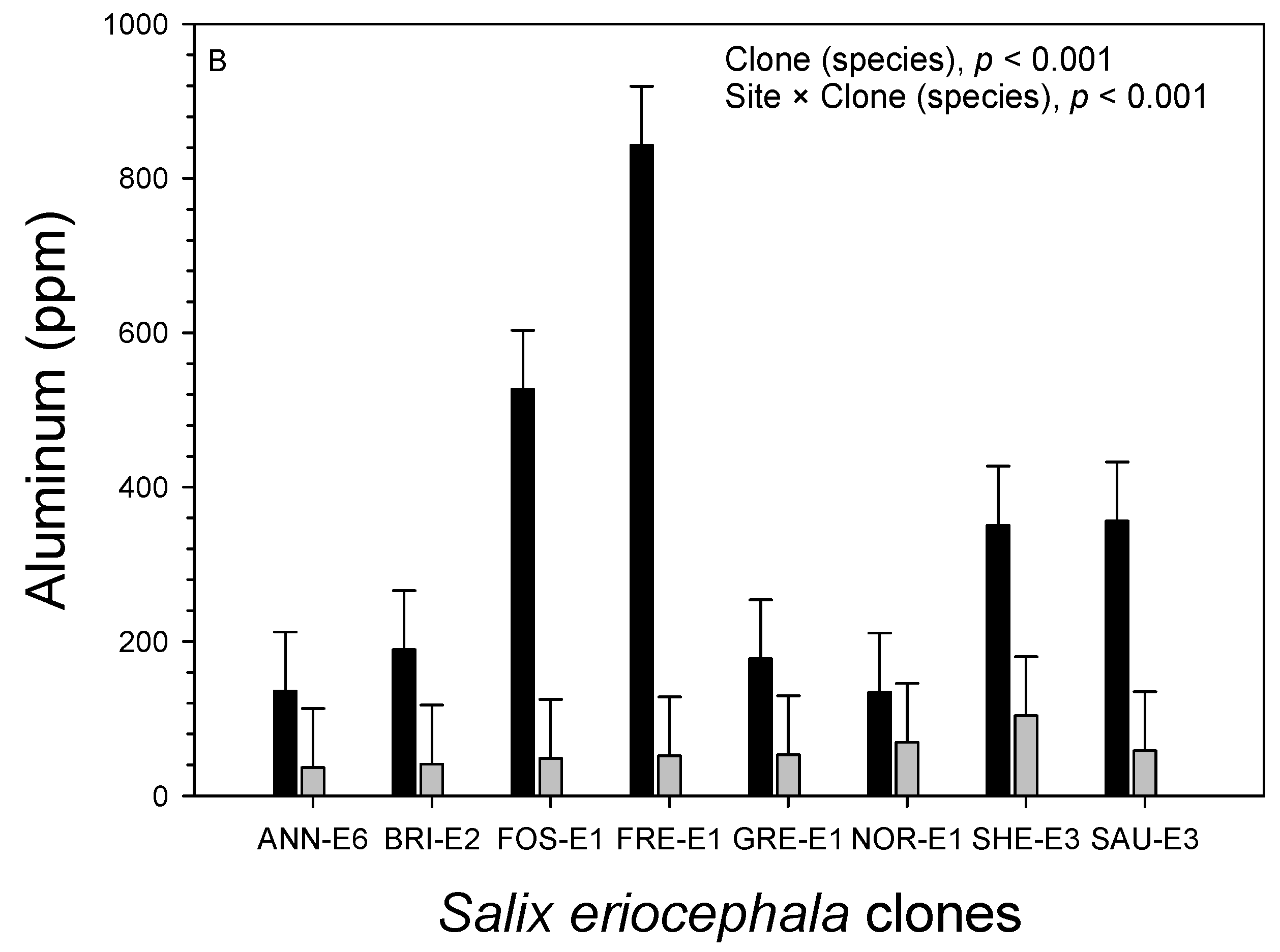
3.3. Foliage
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DIS | Salix discolor |
| EC | electrical conductivity |
| ERI | Salix eriocephala |
| MBG | Montreal Botanical Garden |
| Al | Aluminum |
| C | Carbon |
| Ca | Calcium |
| Fe | Iron |
| INT | Salix interior |
| K | Potassium |
| Mg | Magnesium |
| Mn | Manganese |
| Na | Sodium |
| N | Nitrogen |
| NB | New Brunswick |
| ON | Ontario |
| P | Phosphorous |
| QC | Quebec |
| S | Sulfur |
| VIM | Salix viminalis |
| Zn | Zinc |
References
- Mosseler, A.; Zsuffa, L.; Stoehr, M.U.; Kenney, W.A. Variation in biomass production, moisture content, and specific gravity in some North American willows (Salix L.). Can. J. For. Res. 1988, 18, 1535–1540. [Google Scholar] [CrossRef]
- Zsuffa, L. Genetic improvement of willows for energy plantations. Biomass 1990, 22, 35–47. [Google Scholar] [CrossRef]
- Labrecque, M.; Teodorescu, T.I. Field performance and biomass production of 12 willow and poplars in short-rotation coppice in southern Quebec (Canada). Biomass Bioenergy 2005, 29, 1–9. [Google Scholar]
- Volk, T.A.; Abrahamson, L.P.; Nowak, C.A.; Smart, L.B.; Tharakan, P.J.; White, E.H. The development of short-rotation willow in the northeastern United States for bioenergy and bio-products, agroforestry and phytoremediation. Biomass Bioenergy 2006, 30, 715–727. [Google Scholar] [CrossRef]
- Zalesny, R.S.; Stanturf, J.A.; Gardiner, E.S.; Perdue, J.H.; Young, T.M.; Coyle, D.R.; Headlee, W.L.; Banuellos, G.S.; Hass, A. Ecosystem services of woody crop production systems. Bioenergy Res. 2016, 9, 465–491. [Google Scholar] [CrossRef]
- Labrecque, M.; Teodorescu, T.I.; Daigle, S. Effect of wastewater sludge on growth and heavy metal bioaccumulation of two Salix species. Plant Soil 1994, 171, 303–316. [Google Scholar] [CrossRef]
- Labrecque, M.; Teodorescu, T.I. Influence of plantation site and wastewater sludge fertilization on the performance and foliar nutrient status of two willow species grown under SRIC in southern Quebec (Canada). For. Ecol. Manag. 2001, 150, 223–239. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Knee, M.; Quigley, M.F. Cadmium and copper uptake and translocation in five willow species. Int. J. Phytorem. 2004, 6, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Bourret, M.M.; Brummer, J.E.; Leininger, W.C.; Heil, D.M. Effect of water table on willows grown in amended mine tailing. J. Environ. Qual. 2005, 34, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Kuzovkina, Y.A.; Quigley, M.F. Willows beyond wetlands: Uses of Salix L. species for environmental projects. WASP 2005, 162, 183–204. [Google Scholar] [CrossRef]
- Meers, E.; Lamsal, S.; Vervaeke, P.; Hopgood, M.; Lust, N.; Tack, F.M.G. Availability of heavy metals for uptake by Salix viminalis on a moderately contaminated dredged sediment disposal site. Environ. Pollut. 2005, 137, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Meers, E.; Vandecasteele, B.; Ruttens, A.; Vangronsveld, J.; Tack, F.M.G. Potential of five willow species (Salix spp.) for phytoextraction of heavy metals. Environ. Exp. Bot. 2007, 60, 57–68. [Google Scholar] [CrossRef]
- Shanahan, J.O.; Brummer, J.E.; Leininger, W.C.; Paschke, M.W. Manganese and zinc toxicity thresholds for mountain and Geyer willow. Int. J. Phytorem. 2007, 9, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Boyter, M.J.; Brummer, J.E.; Leininger, W.C. Growth and metal accumulation of Geyer and mountain willow in topsoil versus amended mine tailings. WASP 2009, 198, 17–29. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Volk, T.A. The characteristics of willow (Salix L.) varieties for use in ecological engineering applications: Co-ordination of structure, function and autecology. Ecol. Eng. 2009, 35, 1178–1189. [Google Scholar] [CrossRef]
- Witters, N.; van Slycken, S.; Ruttens, A.; Adriaensen, K.; Meers, E.; Meiresonne, L.; Tack, F.M.G.; Thewys, T.; Laes, E.; Vangronsveld, J. Short-rotation coppice of willow for phytoremediation of a metal-contaminated agricultural area: A sustainability assessment. Bioenergy Res. 2009, 2, 144–152. [Google Scholar] [CrossRef]
- Dimitriou, I.; Mola-Yudego, B.; Aronsson, P.; Eriksson, J. Changes in organic carbon and trace elements in the soil of willow short-rotation coppice plantations. Bioenergy Res. 2012, 5, 563–572. [Google Scholar] [CrossRef]
- Tordoff, G.M.; Baker, A.J.M.; Willis, A.J. Current approaches to the revegetation and reclamation of metalliferous mine wastes. Chemosphere 2000, 41, 219–228. [Google Scholar] [CrossRef]
- Landberg, T.; Greger, M. Differences in uptake and tolerance to heavy metals in Salix from unpolluted and polluted areas. Appl. Geochem. 1996, 11, 175–180. [Google Scholar] [CrossRef]
- Landberg, T.; Greger, M. Differences in oxidative stress in heavy metal resistant and sensitive clones of Salix viminalis. J. Plant Physiol. 2002, 159, 69–75. [Google Scholar] [CrossRef]
- Nissen, L.R.; Lepp, N.W. Baseline concentrations of copper and zinc in shoot tissues of a range of Salix species. Biomass Bioenergy 1997, 12, 115–120. [Google Scholar] [CrossRef]
- Perttu, K.L.; Kowalik, P.J. Salix vegetation filters for purification of water and soils. Biomass Bioenergy 1997, 12, 9–19. [Google Scholar] [CrossRef]
- Punshon, T.; Dickinson, N.M. Acclimation of Salix to metal stress. New Phytol. 1997, 137, 303–314. [Google Scholar] [CrossRef]
- Punshon, T.; Dickinson, N.M. Mobilization of heavy metals using short-rotation coppice. Asp. Appl. Biol. 1997, 49, 285–292. [Google Scholar]
- Punshon, T.; Dickinson, N.M. Heavy metal resistance and accumulation characteristics in willows. Int. J. Phytorem. 1999, 4, 361–385. [Google Scholar] [CrossRef]
- Aronsson, P.; Perttu, K. Willow vegetation filters for wastewater treatment and soil remediation combined with biomass production. For. Chron. 2001, 77, 293–298. [Google Scholar] [CrossRef]
- Pulford, I.D.; Riddell-Black, D.M.; Stewart, C. Heavy metal uptake by willow clones from sewage sludge-treated soil: The potential for phytoremediation. Int. J. Phytorem. 2002, 4, 59–72. [Google Scholar] [CrossRef]
- Pulford, I.D.; Watson, C. Phytoremediation of heavy metal-contaminated land by trees—A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef]
- Hammer, D.; Kayser, A.; Keller, C. Phytoextraction of Cd and Zn with Salix viminalis in field trials. Soil Use Manag. 2002, 19, 187–192. [Google Scholar] [CrossRef]
- Klang-Westin, E.; Eriksson, J. Potential of Salix as phytoextractor for Cd on moderately contaminated soils. Plant Soil 2003, 249, 127–137. [Google Scholar] [CrossRef]
- Keller, C.; Hammer, D.; Kayser, A.; Richner, W.; Brodbeck, M.; Sennhauser, M. Root development and heavy metal phytoextraction efficiency: Comparison of different species in the field. Plant Soil 2003, 249, 67–81. [Google Scholar] [CrossRef]
- Vervaeke, P.; Luyssaert, S.; Mertens, J.; Meers, E.; Tack, F.M.G.; Lust, N. Phytoremediation prospects of willow stands on contaminated sediments: A field trial. Environ. Pollut. 2003, 126, 275–282. [Google Scholar] [CrossRef]
- Bagatto, G.; Shorthouse, J.D. Biotic and abiotic characteristics of ecosystems on acid metalliferous mine tailings near Sudbury, Ontario. Can. J. Bot. 1999, 77, 410–425. [Google Scholar]
- Berti, W.R.; Cunningham, S.D. Phytostabilization of metals. In Phytoremediation of Toxic Metals—Using Plants to Clean Up the Environment; Raskin, I., Ensley, B.D., Eds.; John Wiley and Sons: New York, NY, USA, 2000; pp. 71–88. [Google Scholar]
- Green, C.H.; Heil, D.M.; Cardon, G.E.; Butters, G.L.; Kelly, E.F. Solubilization of manganese and trace metals in soils affected by acid mine runoff. J. Environ. Qual. 2003, 32, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, W.; Keller, C.; Boschi, K. Phytoextraction capacity of trees growing on a metal contaminated soil. Plant Soil 2003, 256, 265–272. [Google Scholar] [CrossRef]
- Chaney, R.L. Zinc phytotoxicity. In Proceedings of the International Symposium on Zinc in Soils and Plants, Perth, Australia, 27–28 September 1993; Robson, A.D., Ed.; Kluwer Academic: London, UK, 1993; pp. 135–149. [Google Scholar]
- Kahle, H. Response of roots of trees to heavy metals. Environ. Exp. Bot. 1993, 33, 99–119. [Google Scholar] [CrossRef]
- Tack, F.M.G.; Callewaert, O.W.; Verloo, M.G. Metal solubility as a function of pH in contaminated, dredged sediment affected by oxidation. Environ. Pollut. 1996, 91, 199–208. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T. Use of willow in phytoextraction. Int. J. Phytorem. 1999, 1, 115–123. [Google Scholar] [CrossRef]
- Rout, G.R.; Samantaray, S.; Das, P. Aluminium toxicity in plants: A review. Agronomie 2001, 21, 3–21. [Google Scholar] [CrossRef]
- Mariano, E.D.; Pinheiro, A.S.; Garcia, E.F.; Keltjens, W.G.; Jorge, R.A.; Menossi, M. Differential aluminium-impaired nutrient uptake along the root axis of two maize genotypes contrasting in resistance to aluminium. Plant Soil 2015, 388, 323–335. [Google Scholar] [CrossRef]
- Rehmus, A.; Bigalke, M.; Valarezo, C.; Castillo, J.M.; Wilcke, W. Aluminum toxicity to tropical montane forest tree seedlings in southern Ecuador: Response of nutrient status to elevated Al concentrations. Plant Soil 2015, 388, 87–97. [Google Scholar] [CrossRef]
- Andersson, M. Toxicity and tolerance of aluminium in vascular plants. WASP 1988, 39, 439–462. [Google Scholar]
- Mosseler, A.; Major, J.E. Coppice growth responses of two North American willows (Salix spp.) in acidic clay deposits on coal mine overburden. Can. J. Plant Sci. 2014, 94, 1269–1279. [Google Scholar] [CrossRef]
- Argus, G.W. Salix L. In Flora of North America North of Mexico, Volume 7: Magnoliophyta: Salicaceae to Brassicaceae; Editorial Committee, Ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2010; pp. 23–162. [Google Scholar]
- Russell, W.B.; La Roi, G.H. Natural vegetation and ecology of abandoned coal-mined land, Rocky Mountain Foothills, Alberta, Canada. Can. J. Bot. 1986, 64, 1286–1298. [Google Scholar] [CrossRef]
- Strong, W.L. Vegetation development on reclaimed lands in the Coal Valley Mine of western Alberta, Canada. Can. J. Bot. 2000, 78, 110–118. [Google Scholar]
- Mosseler, A.; Major, J.E.; Labrecque, M. Growth and survival of seven native willow species on highly disturbed coal mine sites in eastern Canada. Can. J. For. Res. 2014, 44, 1–10. [Google Scholar] [CrossRef]
- McKeague, J.A. (Ed.) Manual on Soil Sampling and Methods of Analysis, 2nd ed.; Canadian Society of Soil Science: Ottawa, ON, Canada, 1978; 212p. [Google Scholar]
- Gullberg, U. Towards making willows pilot species for coppicing production. For. Chron. 1993, 69, 721–726. [Google Scholar] [CrossRef]
- Guidi-Nissim, W.G.; Pitre, F.E.; Teodorescu, T.I.; Labrecque, M. Long-term biomass productivity of willow bioenergy plantations maintained in southern Quebec, Canada. Biomass Bioenergy 2013, 56, 361–369. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E.; Labrecque, M. Genetic by environment interactions of two North American Salix species assessed for coppice yield and components of growth on three sites of varying quality. Trees 2014, 28, 1401–1411. [Google Scholar] [CrossRef]
- Klang-Westin, E.; Perttu, K. Effects of nutrient supply and soil cadmium concentration on cadmium removal by willow. Biomass Bioenergy 2002, 23, 415–426. [Google Scholar] [CrossRef]
- Mleczek, M.; Rutkowski, P.; Rissmann, I.; Kaczmarek, Z.; Golinski, P.; Szentner, K.; Strazynska, K.; Stachowiak, A. Biomass productivity and phytoremediation potential of Salix alba and Salix viminalis. Biomass Bioenergy 2010, 34, 1410–1418. [Google Scholar] [CrossRef]
- Syc, M.; Pohorely, M.; Kamenikova, P.; Habart, J.; Svoboda, K.; Puncochar, M. Willow trees from heavy metals phytoextraction as energy crops. Biomass Bioenergy 2012, 37, 106–113. [Google Scholar] [CrossRef]
- Utmazian, M.N.; Wieshammer, G.; Vega, R.; Wenzel, W.W. Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars. Environ. Pollut. 2007, 148, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Riddell-Black, D.M. Heavy metal uptake by fast growing willow species. In Willow Vegetation Filters for Municipal Wastewaters and Sludges: A Biological Purification System; Aronsson, P., Perttu, K., Eds.; Sveriges Lantbruksuniversiteit: Uppsala, Sweden, 1994; pp. 133–144. [Google Scholar]
- Landberg, T.; Greger, M. Interclonal variation of heavy metal interactions in Salix viminalis. Environ. Toxicol. Chem. 2002, 21, 2669–2674. [Google Scholar] [CrossRef] [PubMed]
- Vyslouzilova, M.; Tlustos, P.; Szakova, J. Cadmium and zinc phytoextraction of seven clones of Salix spp. planted on heavy metal contaminated soils. Plant Soil Environ. 2003, 49, 542–547. [Google Scholar]
- Zottl, H.W. Heavy metal levels and cycling in forest ecosystems. Experientia 1985, 41, 1104–1113. [Google Scholar] [CrossRef]
- Domsch, H.; Giebel, A. Estimation of soil textural features from soil electrical conductivity recorded using the EM38. Precis. Agric. 2004, 5, 389–409. [Google Scholar] [CrossRef]
- Sheldon, A.R.; Dalal, R.C.; Kirchhof, G.; Kopittke, P.M.; Menzies, N.W. The effect of salinity on plant-available water. Plant Soil 2017, 418, 477–491. [Google Scholar] [CrossRef]
- Major, J.E.; Mosseler, A.; Malcolm, J.W.; Heartz, S. Salinity tolerance of three Salix species: Survival, biomass yield and allocation, and biochemical efficiencies. Biomass Bioenergy 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Major, J.E.; Mosseler, A.; Malcolm, J.W. Salix species variation in leaf gas exchange, sodium, and nutrient parameters at three levels of salinity. Can. J. For. Res. 2017, 47, 1045–1055. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Cassaniti, C.; Romano, D.; Hop, M.E.C.M.; Flowers, T.J. Growing floricultural crops with brackish water. Environ. Exp. Bot. 2013, 92, 165–175. [Google Scholar] [CrossRef]
- Dong, Y.; Ma, Y.; Wang, H.; Zhang, J.; Zhang, G.; Yang, M.-S. Assessment of tolerance of willows to saline soils through electrical impedance measurements. For. Sci. Pract. 2013, 15, 32–40. [Google Scholar]
- Alloway, B.J. (Ed.) Heavy Metals in Soils, 2nd ed.; Blaikie Academic and Professional: London, UK, 1995; p. 368. [Google Scholar]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity in plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Larcheveque, M.; Desrochers, A.; Bussiere, B.; Cartier, H.; David, J.-S. Re-vegetation of non-acid-generating, thickened tailings with boreal trees: A greenhouse study. J. Environ. Qual. 2013, 42, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, H.; Wang, A.; Lu, M.; Shen, Z.; Lian, C. Phenotypic plasticity accounts for most of the variation in leaf manganese concentrations in Phytolacca americana growing in manganese-contaminated environments. Plant Soil 2015, 396, 215–227. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press Inc.: London, UK, 1986; 674p. [Google Scholar]
- Hansen, E.A.; McLaughlin, R.A.; Pope, P.E. Biomass and nitrogen dynamics of hybrid poplar on two different soils: Implications for fertilization strategy. Can. J. For. Res. 1988, 18, 223–230. [Google Scholar] [CrossRef]
- Doty, S.L.; Oakley, B.; Xin, G.; Kang, J.W.; Singleton, G.; Khan, Z.; Vajzovic, A.; Staley, J.T. Diazotrophic endophytes of native black cottonwood and willow. Symbiosis 2009, 47, 23–33. [Google Scholar] [CrossRef]
- Knoth, J.L.; Kim, S.-H.; Ettl, G.J.; Doty, S.L. Biological nitrogen fixation and biomass accumulation within poplar clones as a result of inoculation with diazotrophic consortia. New Phytol. 2013, 201, 599–609. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Population Location * | Latitude North | Longitude West | Selected Clones |
|---|---|---|---|---|
| S. discolor | Levis, QC | 46°78’ | 71°18’ | LEV-D3, LEV-D6 |
| Lower Anfield, NB | 46°92’ | 67°49’ | ANF-D1 | |
| Hawkesbury, ON | 45°39’ | 74°75’ | HAW-D5 | |
| Montmagny, QC | 46°94’ | 70°60’ | MON-D1 | |
| Mud Lake, ON | 45°88’ | 76°78’ | MUD-D4 | |
| Richmond Fen, ON | 45°13’ | 75°82’ | RIC-D2 | |
| S. eriocephala | Ste. Anne de la Perade, QC | 46°56’ | 72°20’ | ANN-E6 |
| Bristol, NB | 46°47’ | 67°58’ | BRI-E2 | |
| Fosterville, NB | 45°78’ | 67°76’ | FOS-E1 | |
| Fredericton, NB | 45°94’ | 66°62’ | FRE-E1 | |
| Green River, NB | 47°34’ | 68°19’ | GRE-E1 | |
| Norton, NB | 45°67’ | 65°81’ | NOR-E10 | |
| Shepody Creek, NB | 45°71’ | 64°77’ | SHE-E3 | |
| Rivière au Saumon, QC | 47°21’ | 70°35’ | SAU-E3 |
| Site | Carbon (g kg−1) | Nitrogen (g kg−1) | Potassium (ppm) | Calcium (ppm) | Magnesium (ppm) | Phosphorus (ppm) | Sodium (%) |
|---|---|---|---|---|---|---|---|
| Acidic clay | 53.1 ± 2.3a | 2.07 ± 0.17a | 116.1 ± 14.5a | 2968 ± 116a | 352.4 ± 14.6a | 10.75 ± 0.6a | 0.443 ± 0.031a |
| Shale overburden | 4.6 ± 2.3b | 1.02 ± 0.11b | 91.1 ± 14.5a | 1466 ± 116b | 80.2 ± 14.6b | 3.98 ± 0.65b | 0.022 ± 0.031b |
| Site | Sand (%) | Silt (%) | Clay (%) | pH | C:N ratio | Sulfur (%) | EC 1 (mS cm−1) | WC at Sat. 2 (%) |
|---|---|---|---|---|---|---|---|---|
| Acidic clay | 12.9 ± 2.4b | 44.9 ± 2.5a | 42.3 ± 1.9a | 3.6 ± 0.2b | 25.9 ± 1.4b | 0.079 ± 0.012a | 3.89 ± 0.12a | 64.0 ± 1.4a |
| Shale overburden | 67.2 ± 2.4a | 23.4 ± 2.5b | 9.4 ± 1.9b | 6.8 ± 0.2a | 4.6 ± 1.4a | 0.008 ± 0.012b | 0.88 ± 0.12b | 22.9 ± 1.4b |
| Site | Carbon (%) | Nitrogen (%) | Carbon: Nitrogen Ratio | Phosphorus (%) | Potassium (%) | Calcium (%) | Magnesium (%) |
|---|---|---|---|---|---|---|---|
| Acidic clay | 49.95 ±0.06b | 1.66 ±0.03b | 30.8 ± 0.45a | 0.210 ± 0.004a | 1.01 ± 0.02a | 1.73 ± 0.04b | 0.292 ± 0.007a |
| Shale overburden | 50.26 ±0.06a | 1.75 ± 0.03a | 29.9 ± 0.45a | 0.208 ± 0.004a | 0.99 ± 0.02a | 1.89 ± 0.04a | 0.195 ± 0.007b |
| Site | Manganese (ppm) | Iron (ppm) | Sodium (%) | Zinc (ppm) | Aluminum (ppm) | Sulfur (%) |
|---|---|---|---|---|---|---|
| Acidic clay | 609.5 ± 30.3a | 250.8 ± 26.4a | 0.89 ± 0.04a | 243.3 ± 8.9a | 258.4 ± 19.8a | 0.433 ± 0.014a |
| Shale overburden | 185.4 ± 30.3b | 44.5 ± 26.4b | 0.90 ± 0.04a | 257.3 ± 8.9a | 56.1 ± 19.8b | 0.243 ± 0.014b |
| Source of Variation | df | Carbon (%) | Nitrogen (%) | C:N ratio | Phosphorus (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| MS | p Value | MS | p Value | MS | p Value | MS | p Value | ||
| Block (site) | 4 | 0.98 | 0.001 | 0.18 | <0.001 | 56.3 | <0.001 | 1.8 × 10−3 | 0.096 |
| Species | 1 | 2.20 | 0.001 | 2.87 | <0.001 | 899.3 | <0.001 | 0.4 × 10−3 | 0.508 |
| Site | 1 | 2.20 | 0.001 | 0.18 | 0.021 | 19.5 | 0.149 | 0.1 × 10−3 | 0.849 |
| Species × site | 1 | 1.16 | 0.017 | 0.54 | <0.001 | 142.0 | <0.001 | 2.6 × 10−3 | 0.088 |
| Genotype (species) | 13 | 0.06 | 0.991 | 0.12 | <0.001 | 39.5 | <0.001 | 2.5 × 10−3 | 0.002 |
| Genotype (species) × site | 13 | 0.17 | 0.567 | 0.02 | 0.816 | 7.5 | 0.636 | 1.3 × 10−3 | 0.155 |
| Error | 56 | 0.19 | 0.03 | 9.1 | 0.8 × 10−3 | ||||
| R2 | 0.541 | 0.773 | 0.789 | 0.554 | |||||
| Source of Variation | df | Iron (ppm) | Zinc (ppm) | Aluminum (ppm) | Sulfur (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| MS | p Value | MS | p Value | MS | p Value | MS | p Value | ||
| Block (site) | 4 | 10.3 × 104 | 0.017 | 17.0 × 103 | 0.002 | 10.3 × 104 | 0.039 | 0.014 | 0.164 |
| Species | 1 | 24.8 × 104 | 0.007 | 2.1 × 103 | 0.443 | 24.8 × 104 | 0.005 | 0.202 | <0.001 |
| Site | 1 | 95.4 × 104 | <0.001 | 4.4 × 103 | 0.270 | 95.4 × 104 | <0.001 | 0.812 | <0.001 |
| Species × site | 1 | 17.9 × 104 | 0.020 | 1.4 × 103 | 0.533 | 17.9 × 104 | 0.006 | 0.095 | 0.001 |
| Genotype (species) | 13 | 8.5 × 104 | 0.005 | 24.5 × 103 | <0.001 | 8.5 × 104 | <0.001 | 0.011 | 0.268 |
| Genotype (species) × site | 13 | 8.8 × 104 | 0.004 | 5.5 × 103 | 0.120 | 8.8 × 104 | <0.001 | 0.015 | 0.079 |
| Error | 56 | 3.1 × 104 | 3.5 × 103 | 3.1 × 104 | 0.008 | ||||
| R2 | 0.700 | 0.703 | 0.752 | 0.764 | |||||
| Source of Variation | df | Potassium (%) | Sodium (%) | Calcium (%) | Magnesium (%) | Manganese (ppm) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | p Value | MS | p Value | MS | p Value | MS | p Value | MS | p Value | ||
| Block (site) | 4 | 0.060 | 0.010 | 0.021 | 0.880 | 0.682 | <0.001 | 0.014 | <0.001 | 25.5 × 104 | <0.001 |
| Species | 1 | 0.396 | <0.001 | 0.082 | 0.288 | 3.679 | <0.001 | 0.029 | 0.001 | 0.9 × 104 | 0.647 |
| Site | 1 | 0.015 | 0.341 | 0.002 | 0.878 | 0.575 | 0.007 | 0.210 | <0.001 | 402.8 × 104 | <0.001 |
| Species × site | 1 | <0.001 | 0.955 | 0.134 | 0.176 | 0.004 | 0.822 | 0.002 | 0.358 | 0.4 × 104 | 0.757 |
| Genotype (species) | 13 | 0.109 | <0.001 | 0.143 | 0.038 | 0.242 | 0.001 | 0.012 | <0.001 | 7.2 × 104 | 0.074 |
| Genotype (species) × site | 13 | 0.029 | 0.067 | 0.131 | 0.061 | 0.109 | 0.159 | 0.002 | 0.463 | 1.9 × 104 | 0.938 |
| Error | 56 | 0.016 | 0.072 | 0.074 | 0.002 | 4.1 × 104 | |||||
| R2 | 0.728 | 0.491 | 0.767 | 0.783 | 0.731 | ||||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosseler, A.; Major, J.E. Phytoremediation Efficacy of Salix discolor and S. eriocephela on Adjacent Acidic Clay and Shale Overburden on a Former Mine Site: Growth, Soil, and Foliage Traits. Forests 2017, 8, 475. https://doi.org/10.3390/f8120475
Mosseler A, Major JE. Phytoremediation Efficacy of Salix discolor and S. eriocephela on Adjacent Acidic Clay and Shale Overburden on a Former Mine Site: Growth, Soil, and Foliage Traits. Forests. 2017; 8(12):475. https://doi.org/10.3390/f8120475
Chicago/Turabian StyleMosseler, Alex, and John E. Major. 2017. "Phytoremediation Efficacy of Salix discolor and S. eriocephela on Adjacent Acidic Clay and Shale Overburden on a Former Mine Site: Growth, Soil, and Foliage Traits" Forests 8, no. 12: 475. https://doi.org/10.3390/f8120475
APA StyleMosseler, A., & Major, J. E. (2017). Phytoremediation Efficacy of Salix discolor and S. eriocephela on Adjacent Acidic Clay and Shale Overburden on a Former Mine Site: Growth, Soil, and Foliage Traits. Forests, 8(12), 475. https://doi.org/10.3390/f8120475