Effects of Climate Change on the Climatic Niches of Warm-Adapted Evergreen Plants: Expansion or Contraction?

Abstract
:1. Introduction
2. Materials and Methods
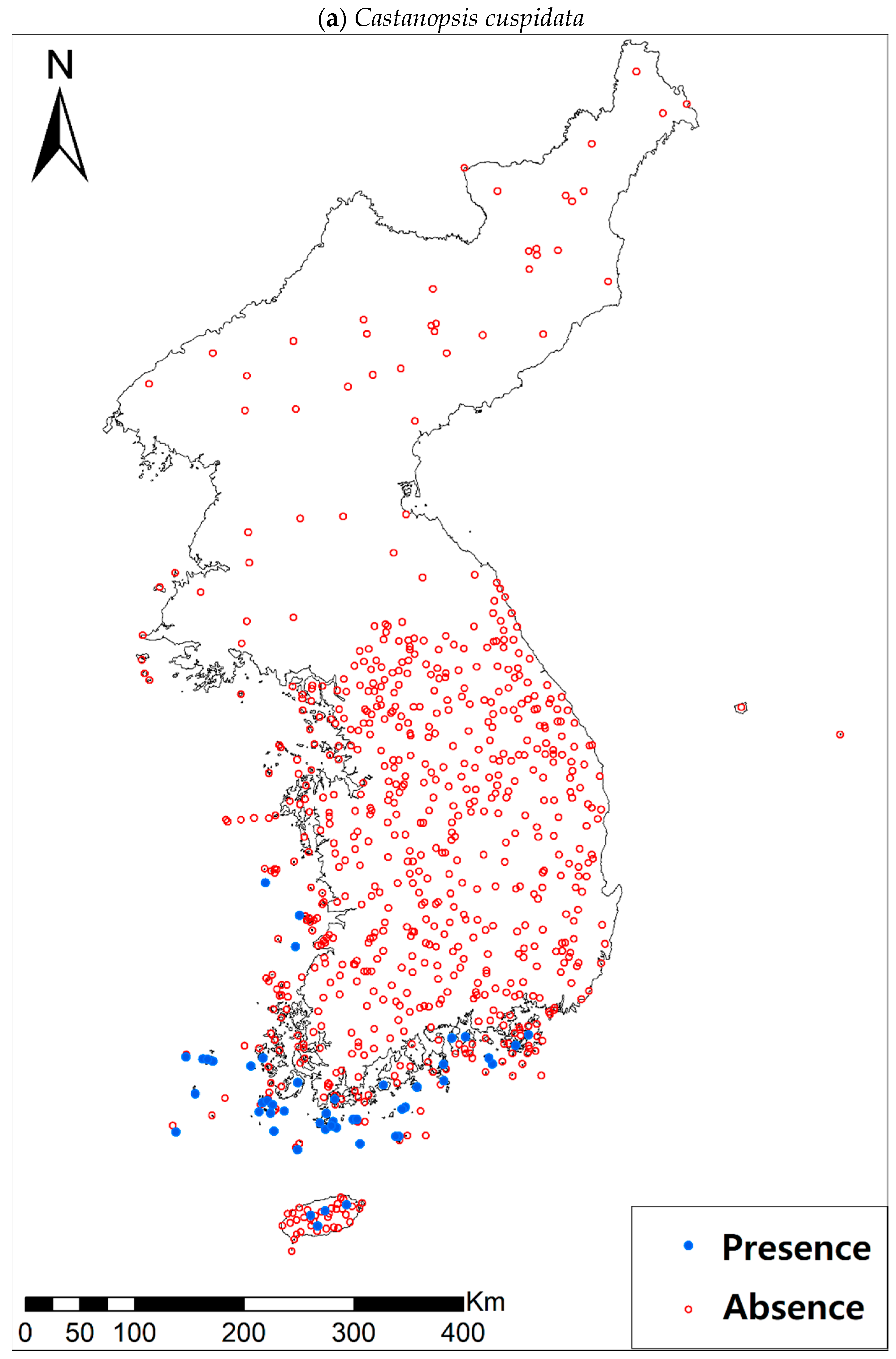
2.1. Study Area and Species
2.2. Climatic Variables
2.3. Species Distribution Modeling and Model Evaluation
2.4. Changes in Habitat Suitability under Climate Change
3. Results
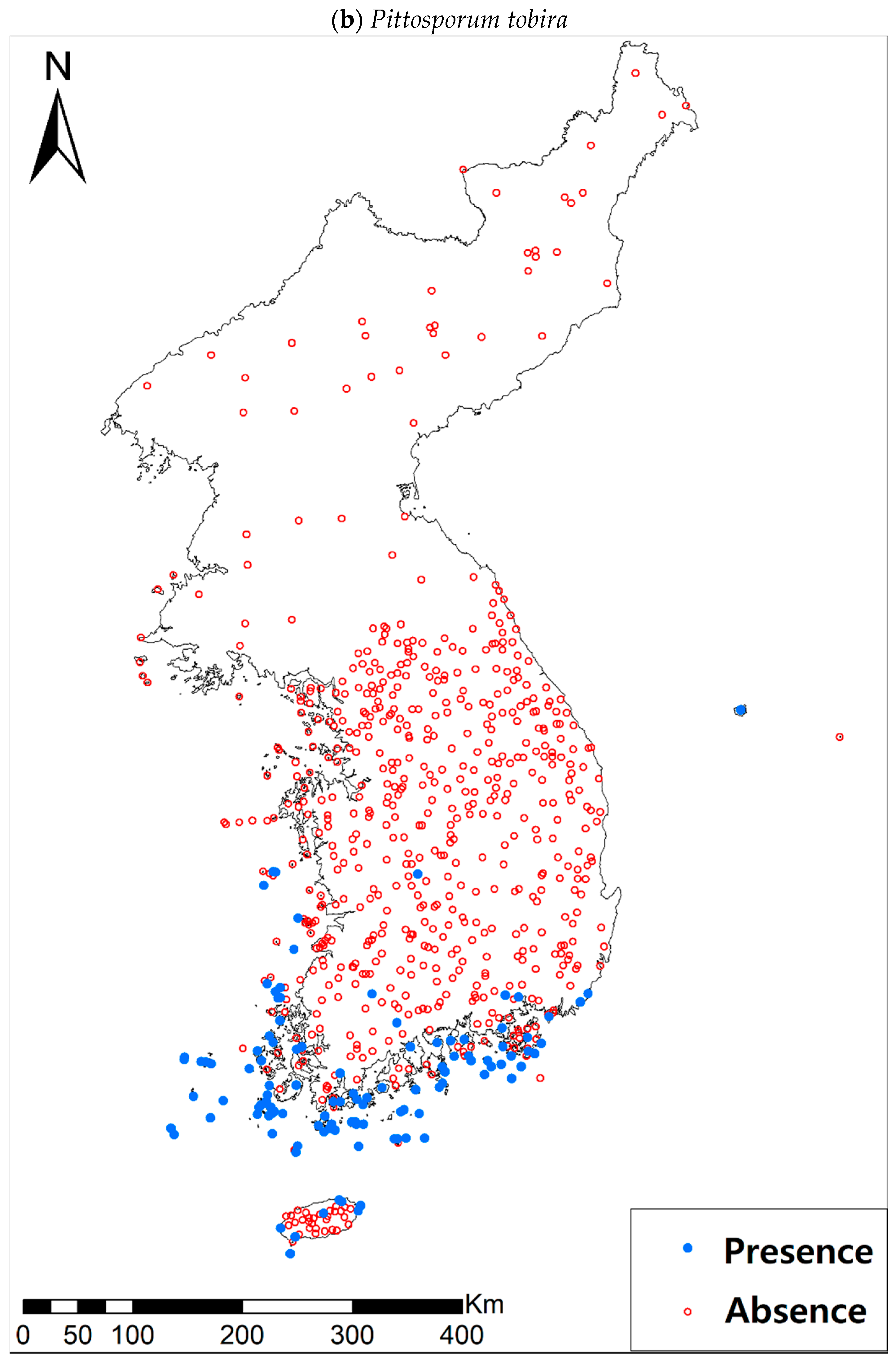
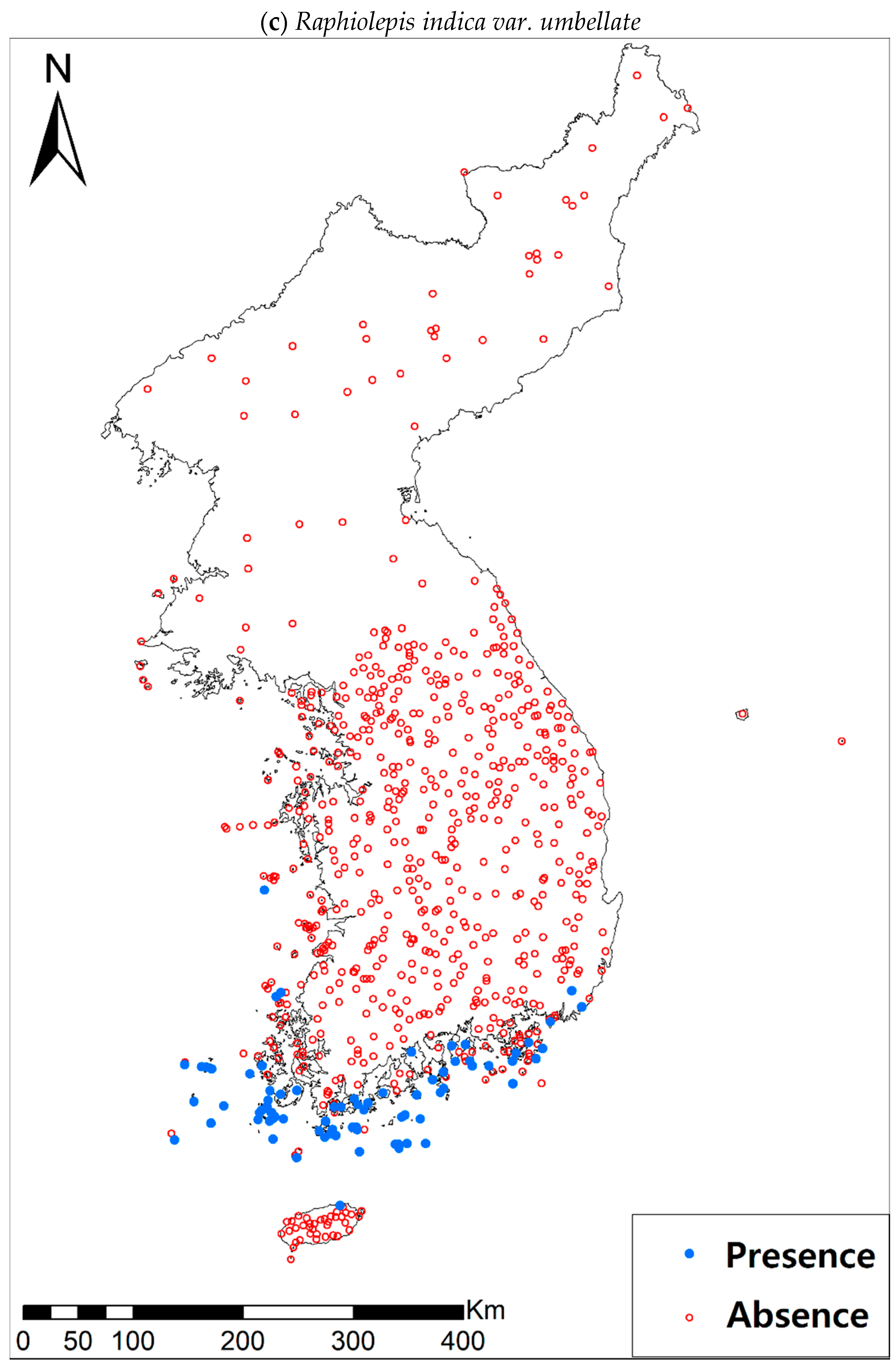
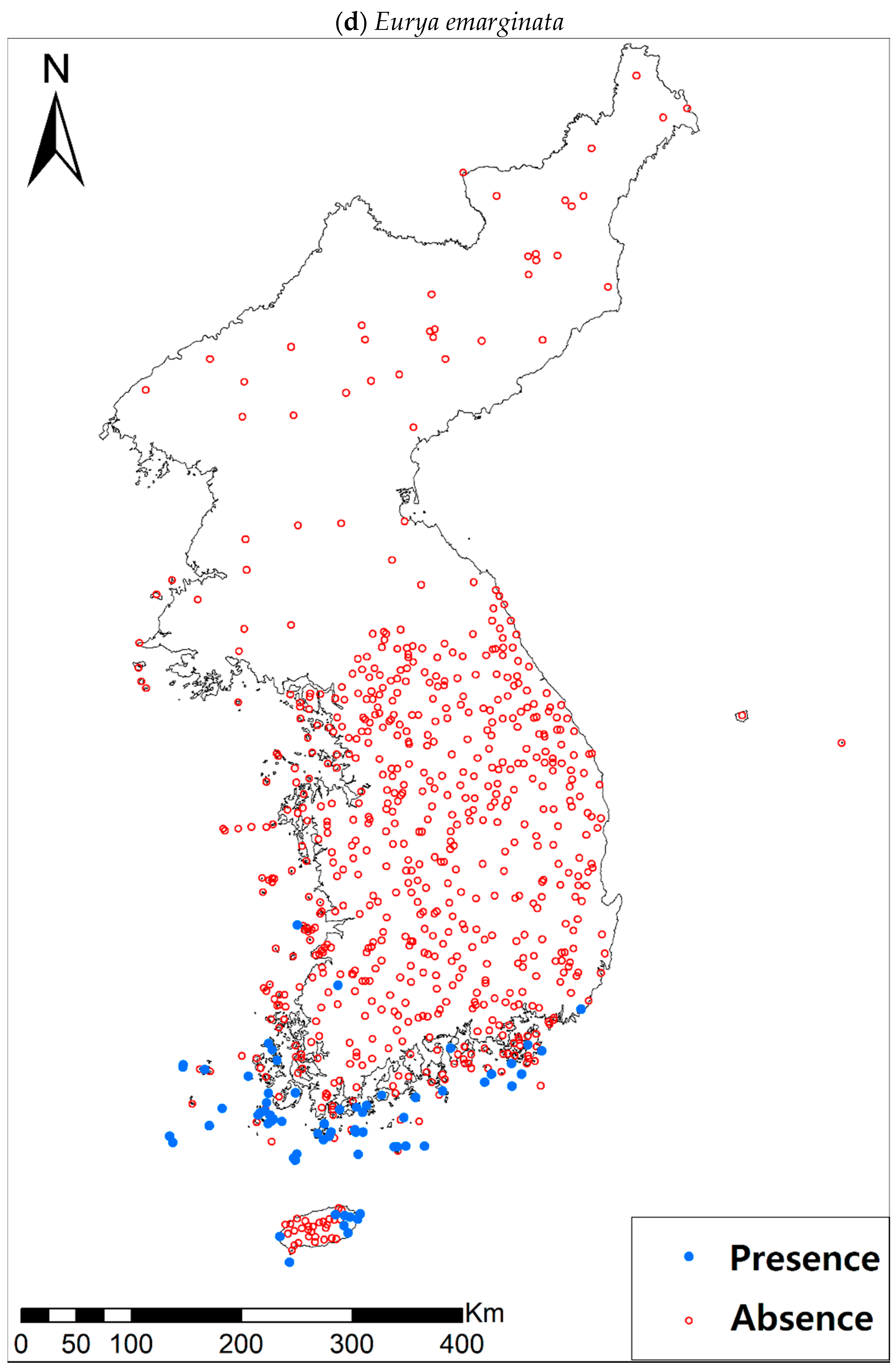
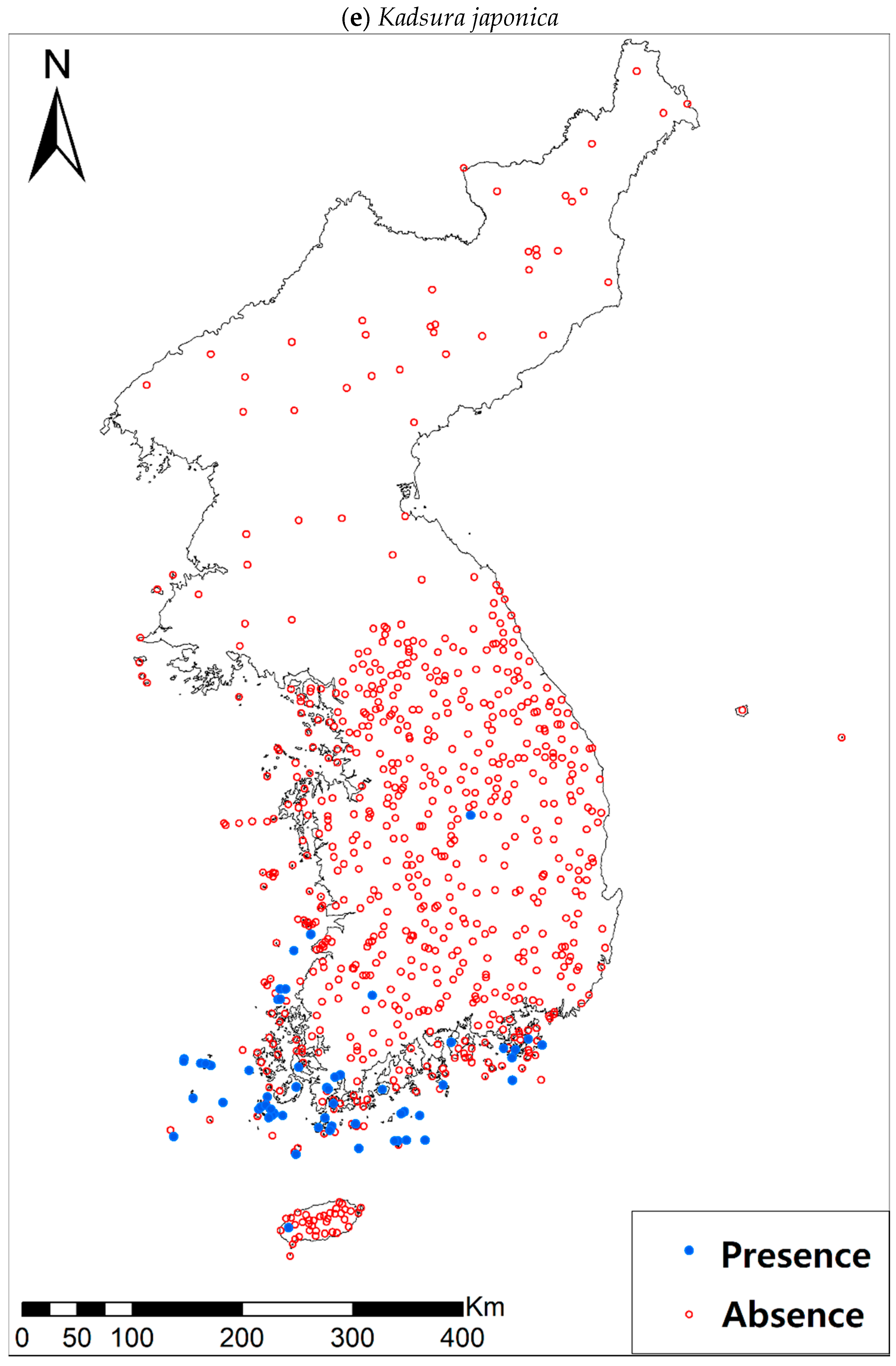
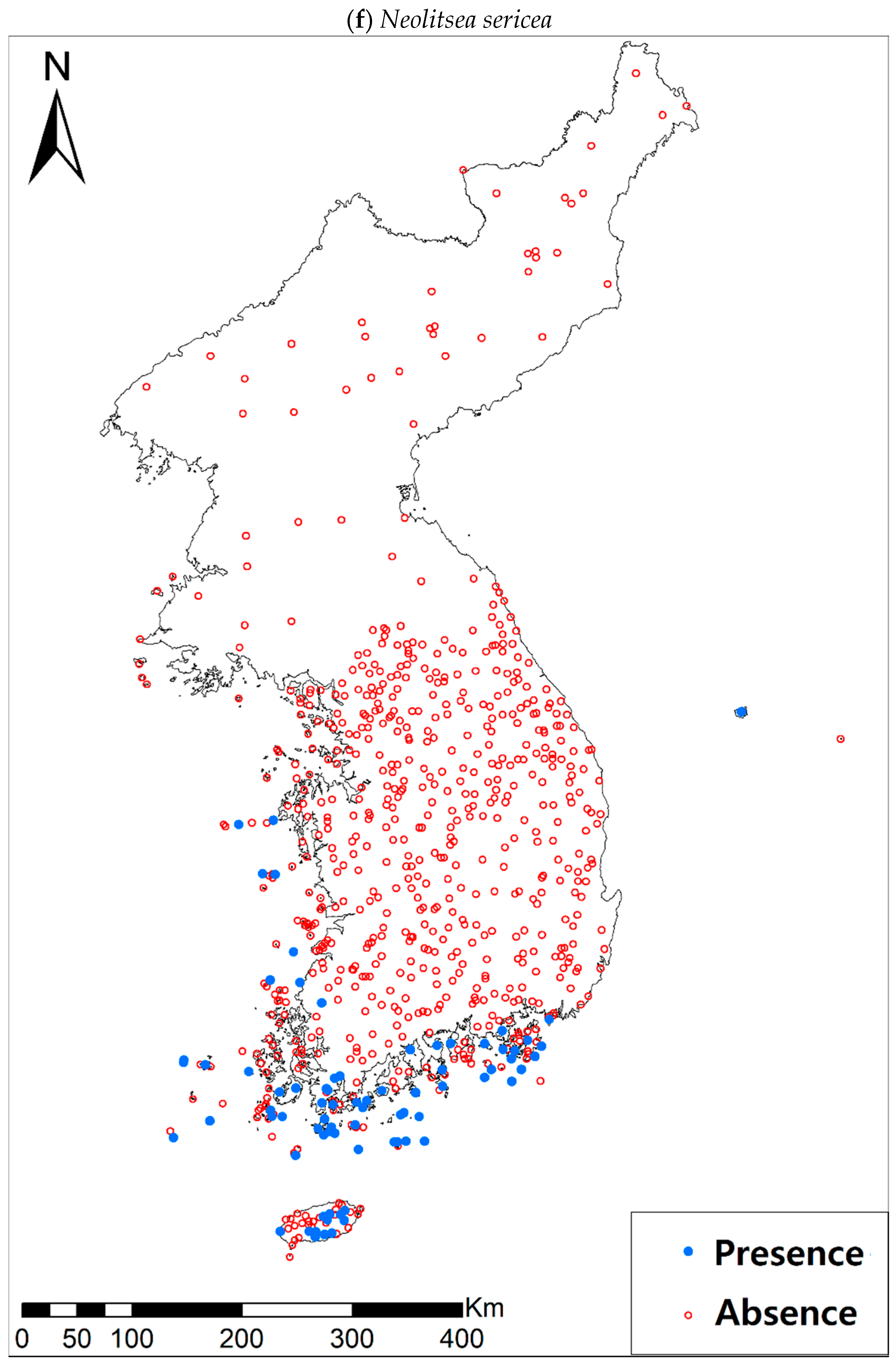
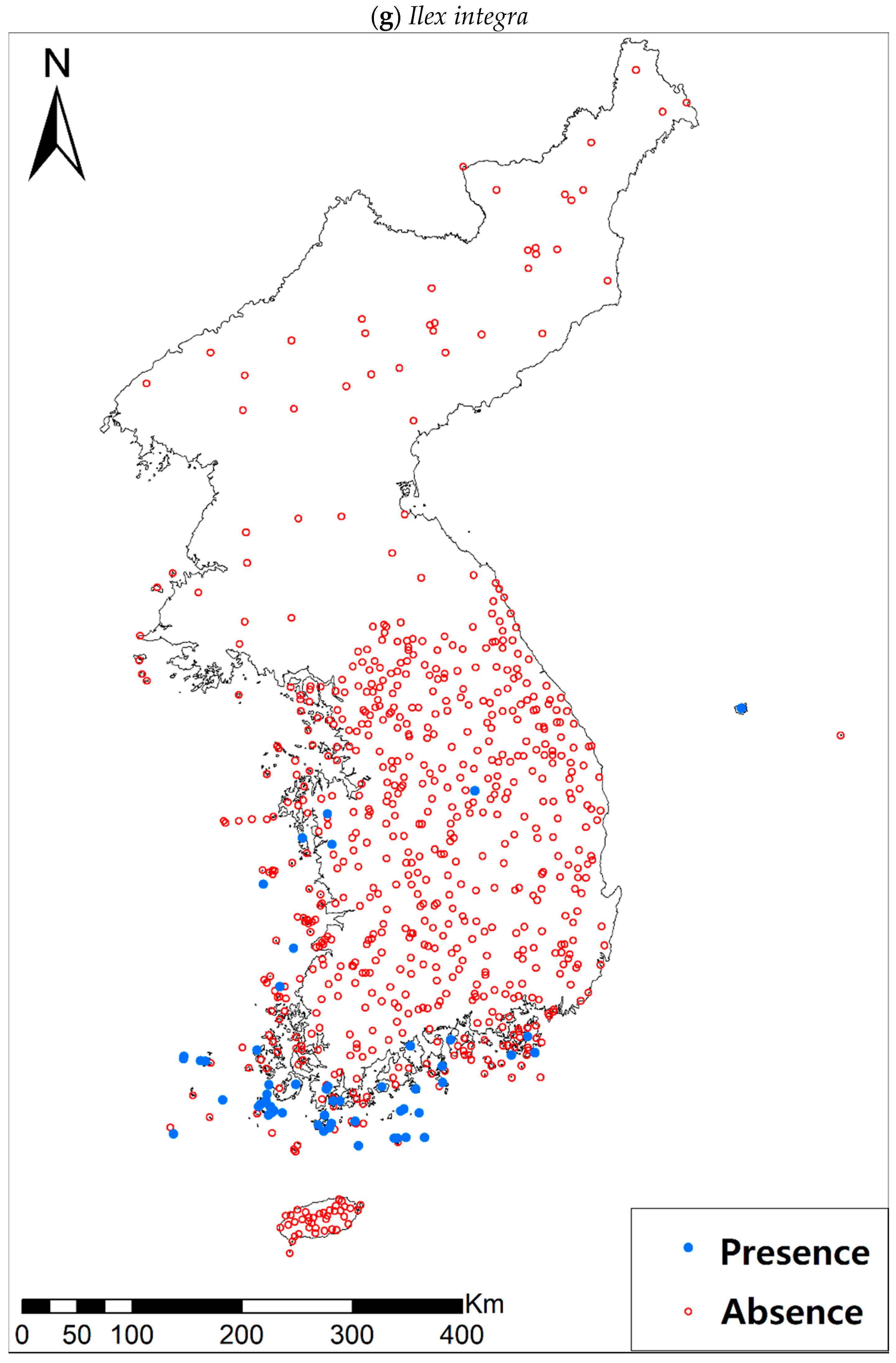
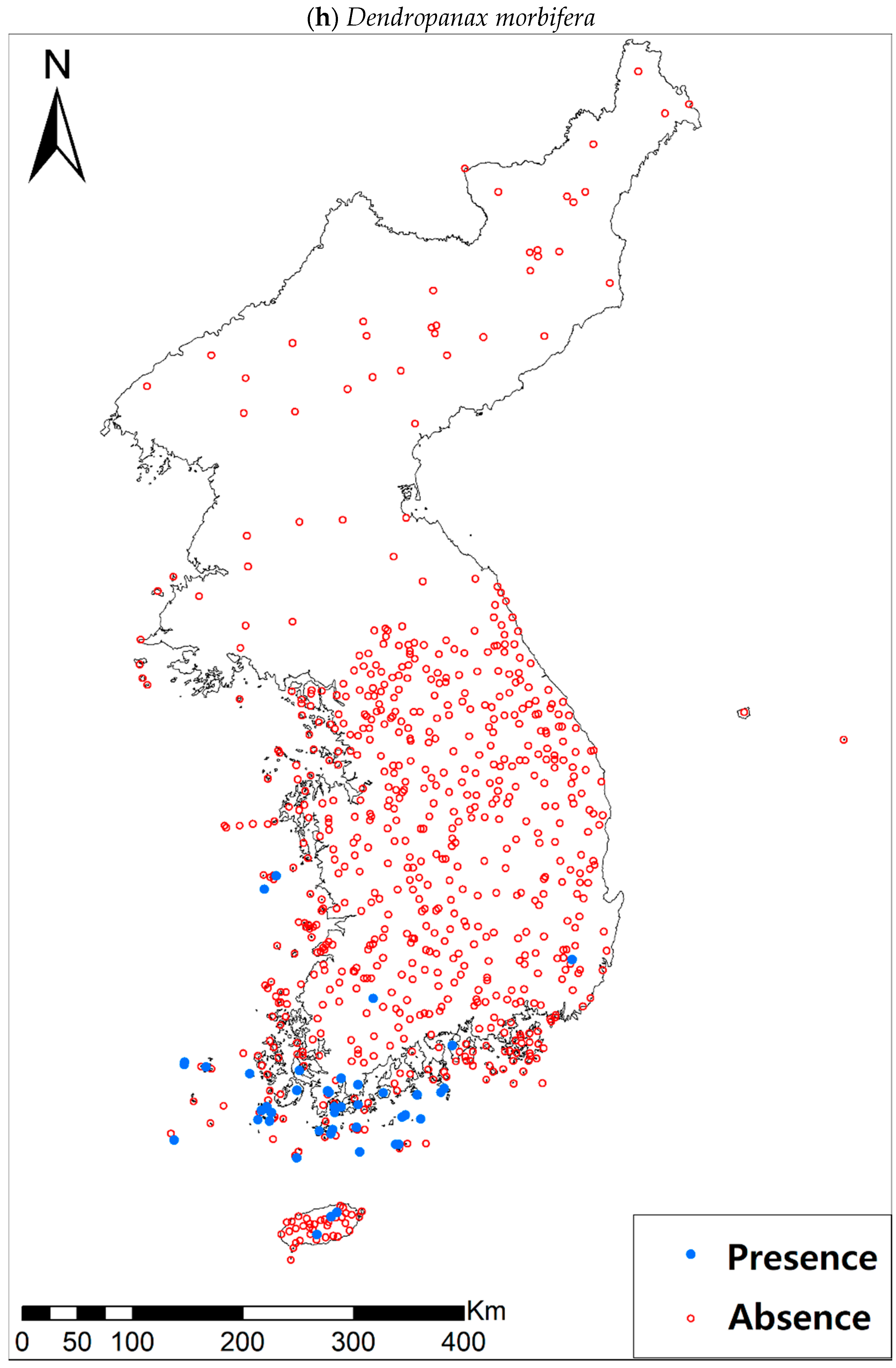
3.1. Model Performance and Current Distribution Patterns
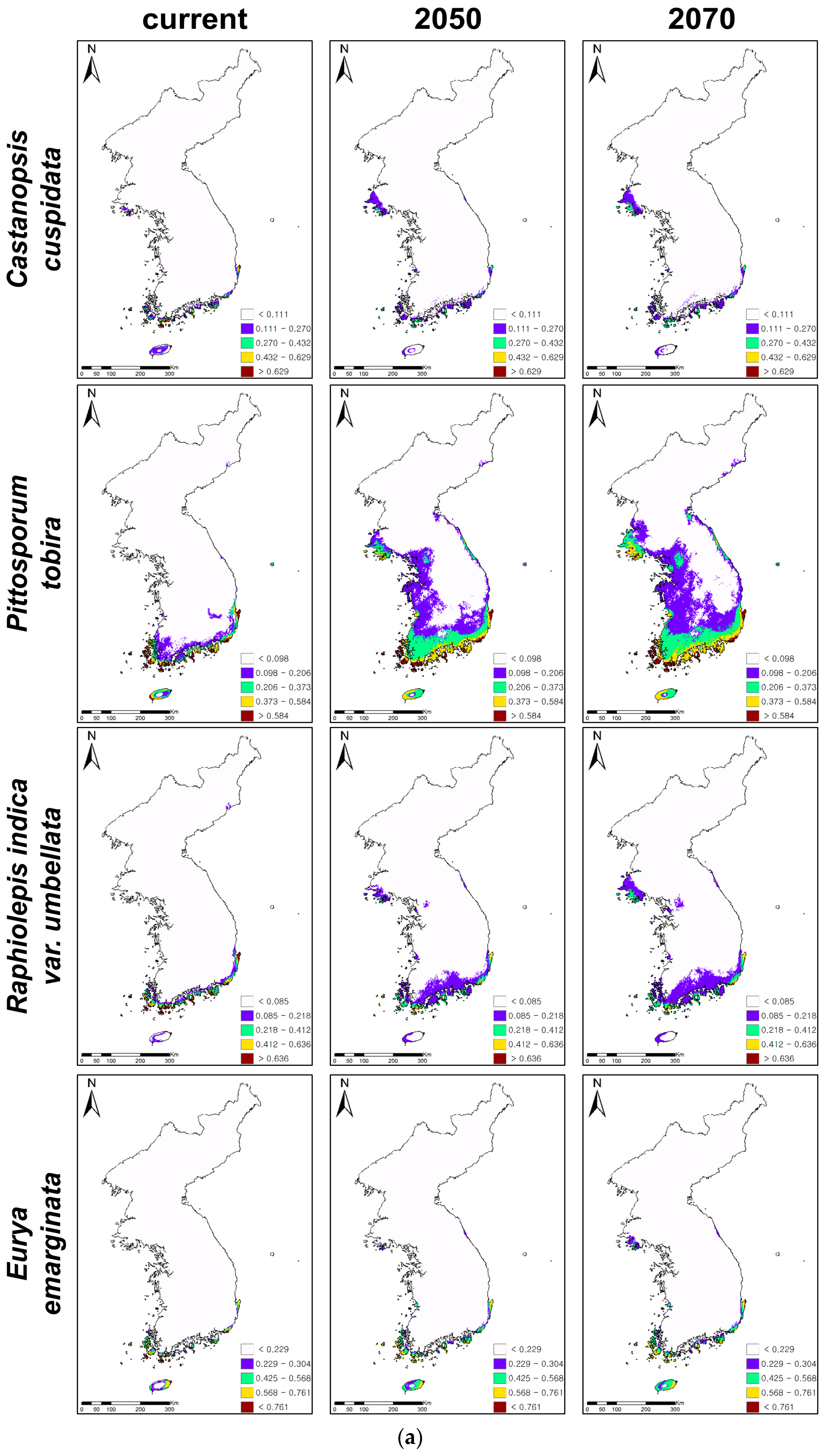
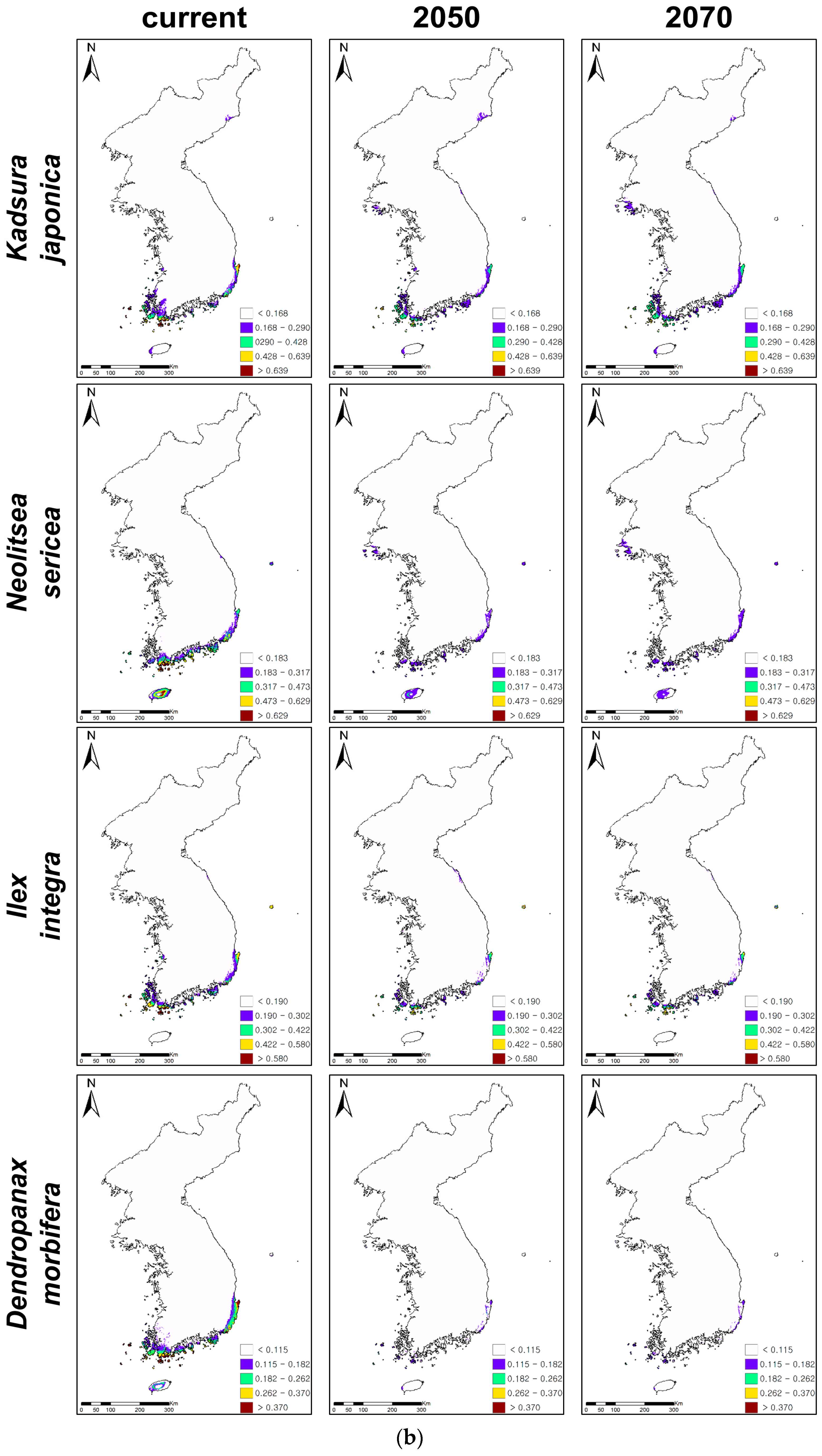
3.2. Future Distributions of Suitable Habitats for Warm-Adapted Evergreens under Climate Changes
4. Discussion
5. Conclusions
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bio1 | Bio2 | Bio3 | Bio4 | Bio5 | Bio6 | Bio7 | Bio8 | Bio9 | Bio10 | Bio11 | Bio12 | Bio13 | Bio14 | Bio15 | Bio16 | Bio17 | Bio18 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bio2 | −0.52 | |||||||||||||||||
| Bio3 | −0.02 | 0.61 | ||||||||||||||||
| Bio4 | −0.72 | 0.79 | 0.02 | |||||||||||||||
| Bio5 | 0.93 | −0.24 | 0.04 | −0.43 | ||||||||||||||
| Bio6 | 0.95 | −0.7 | −0.08 | −0.89 | 0.79 | |||||||||||||
| Bio7 | −0.68 | 0.87 | 0.16 | 0.99 | −0.39 | −0.87 | ||||||||||||
| Bio8 | 0.96 | −0.34 | −0.01 | −0.52 | 0.99 | 0.85 | −0.48 | |||||||||||
| Bio9 | 0.96 | −0.65 | −0.03 | −0.86 | 0.81 | 0.99 | −0.84 | 0.86 | ||||||||||
| Bio10 | 0.96 | −0.36 | 0 | −0.54 | 0.97 | 0.85 | −0.5 | 0.98 | 0.87 | |||||||||
| Bio11 | 0.98 | −0.63 | −0.03 | −0.85 | 0.84 | 0.99 | −0.82 | 0.89 | 0.99 | 0.89 | ||||||||
| Bio12 | 0.07 | −0.09 | 0.34 | −0.35 | −0.1 | 0.14 | −0.29 | −0.07 | 0.19 | 0.19 | 0.15 | |||||||
| Bio13 | −0.24 | 0.5 | 0.27 | 0.43 | −0.12 | −0.38 | 0.48 | −0.18 | −0.33 | −0.33 | −0.32 | 0.57 | ||||||
| Bio14 | 0.18 | −0.43 | 0.06 | −0.58 | −0.07 | 0.36 | −0.59 | 0 | 0.32 | 0.32 | 0.31 | 0.53 | −0.09 | |||||
| Bio15 | −0.45 | 0.72 | 0.05 | 0.88 | −0.17 | −0.68 | 0.88 | −0.26 | −0.63 | −0.63 | −0.61 | −0.16 | 0.67 | −0.69 | ||||
| Bio16 | −0.18 | 0.32 | 0.37 | 0.13 | −0.19 | −0.23 | 0.19 | −0.21 | −0.16 | −0.16 | −0.19 | 0.85 | 0.89 | 0.16 | 0.37 | |||
| Bio17 | 0.34 | −0.55 | 0.07 | −0.75 | 0.06 | 0.53 | −0.75 | 0.14 | 1 | 0.5 | 0.48 | 0.58 | −0.16 | 0.95 | −0.8 | 0.14 | ||
| Bio18 | −0.24 | 0.41 | 0.4 | 0.24 | −0.21 | −0.31 | 0.3 | −0.24 | −0.25 | −0.25 | −0.26 | 0.79 | 0.93 | 0.12 | 0.45 | 0.98 | 0.07 | |
| Bio19 | 0.27 | −0.54 | 0.06 | −0.72 | −0.01 | 0.47 | −0.72 | 0.07 | 0.45 | 0.45 | 0.42 | 0.6 | −0.14 | 0.95 | −0.78 | 0.16 | 0.98 | 0.1 |
References
- Alamgir, M.; Mukul, S.A.; Turton, S.M. Modelling spatial distribution of critically endangered asian elephant and hoolock gibbon in Bangladesh forest ecosystems under a changing climate. Appl. Geogr. 2015, 60, 10–19. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Svenning, J.C. Climate-related range shifts—A global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Costion, C.M.; Simpson, L.; Pert, P.L.; Carlsen, M.M.; Kress, W.J.; Crayn, D. Will tropical mountaintop plant species survive climate change? Identifying key knowledge gaps using species distribution modelling in Australia. Biol. Conserv. 2015, 191, 322–330. [Google Scholar] [CrossRef]
- Feeley, K.J.; Davies, S.J.; Perez, R.; Hubbell, S.P.; Foster, R.B. Directional changes in the species composition of a tropical forest. Ecology 2011, 92, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J.; Silman, M.R.; Bush, M.B.; Farfan, W.; Cabrera, K.G.; Malhi, Y.; Meir, P.; Revilla, N.S.; Quisiyupanqui, M.N.R.; Saatchi, S. Upslope migration of Andean trees. J. Biogeogr. 2011, 38, 783–791. [Google Scholar] [CrossRef]
- Nakao, K.; Matsui, T.; Horikawa, M.; Tsuyama, I.; Tanaka, N. Assessing the impact of land use and climate change on the evergreen broad-leaved species of quercus acuta in japan. Plant Ecol. 2011, 212, 229–243. [Google Scholar] [CrossRef]
- Klötzli, F.; Walther, G.-R. Recent Vegetation Shifts in Switzerland. In Conference on Recent Shifts in Vegetation Boundaries of Deciduous Forests, Especially Due to General Global Warming; Springer: Berlin, Germany, 1999; pp. 15–29. [Google Scholar]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.J. Distributional migrations, expansions, and contractions of tropical plant species as revealed in dated herbarium records. Glob. Chang. Biol. 2012, 18, 1335–1341. [Google Scholar] [CrossRef]
- Feeley, K.J.; Malhi, Y.; Zelazowski, P.; Silman, M.R. The relative importance of deforestation, precipitation change, and temperature sensitivity in determining the future distributions and diversity of amazonian plant species. Glob. Chang. Biol. 2012, 18, 2636–2647. [Google Scholar] [CrossRef]
- Nakao, K.; Higa, M.; Tsuyama, I.; Lin, C.-T.; Sun, S.-T.; Lin, J.-R.; Chiou, C.-R.; Chen, T.-Y.; Matsui, T.; Tanaka, N. Changes in the potential habitats of 10 dominant evergreen broad-leaved tree species in the Taiwan-Japan archipelago. Plant Ecol. 2014, 215, 639–650. [Google Scholar] [CrossRef]
- Wright, S.J.; Muller-Landau, H.C.; Schipper, J. The future of tropical species on a warmer planet. Conserv. Biol. 2009, 23, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.A.; Kong, W.-S.; Park, S.U.; Lee, J.H.; Kim, J.; Jung, H. Sensitivity of Korean fir (Abies koreana wils.), a threatened climate relict species, to increasing temperature at an island subalpine area. Ecol. Model. 2017, 353, 5–16. [Google Scholar] [CrossRef]
- Osland, M.J.; Enwright, N.M.; Day, R.H.; Gabler, C.A.; Stagg, C.L.; Grace, J.B. Beyond just sea-level rise: Considering macroclimatic drivers within coastal wetland vulnerability assessments to climate change. Glob. Chang. Biol. 2016, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.M. The impact of climate change on mangrove forests. Curr. Clim. Chang. Rep. 2015, 1, 30–39. [Google Scholar] [CrossRef]
- Valle, M.; Chust, G.; del Campo, A.; Wisz, M.S.; Olsen, S.M.; Garmendia, J.M.; Borja, Á. Projecting future distribution of the seagrass Zostera noltii under global warming and sea level rise. Biol. Conserv. 2014, 170, 74–85. [Google Scholar] [CrossRef]
- McKee, K.; Rogers, K.; Saintilan, N. Response of salt marsh and mangrove wetlands to changes in atmospheric CO2, climate, and sea level. In Global Change and the Function and Distribution of Wetlands; Springer: Berlin, Germany, 2012; pp. 63–96. [Google Scholar]
- Di Nitto, D.; Neukermans, G.; Koedam, N.; Defever, H.; Pattyn, F.; Kairo, J.G.; Dahdouh-Guebas, F. Mangroves facing climate change: Landward migration potential in response to projected scenarios of sea level rise. Biogeosciences 2014, 11, 857. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.; Burnside, N.; Joyce, C.; Sepp, K.; Teasdale, P. Improved modelling of the impacts of sea level rise on coastal wetland plant communities. Hydrobiologia 2016, 774, 203–216. [Google Scholar] [CrossRef]
- Armitage, A.R.; Highfield, W.E.; Brody, S.D.; Louchouarn, P. The contribution of mangrove expansion to salt marsh loss on the Texas Gulf Coast. PLoS ONE 2015, 10, e0125404. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.T.; Sundareshwar, P.; Nietch, C.T.; Kjerfve, B.; Cahoon, D.R. Responses of coastal wetlands to rising sea level. Ecology 2002, 83, 2869–2877. [Google Scholar] [CrossRef]
- Garner, K.L.; Chang, M.Y.; Fulda, M.T.; Berlin, J.A.; Freed, R.E.; Soo-Hoo, M.M.; Revell, D.L.; Ikegami, M.; Flint, L.E.; Flint, A.L. Impacts of sea level rise and climate change on coastal plant species in the central California Coast. PeerJ 2015, 3, e958. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.; Hartley, A.; Butchart, S.; Willis, S. Choice of baseline climate data impacts projected species’ responses to climate change. Glob. Chang. Biol. 2016, 22, 2392–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearney, M.R.; Wintle, B.A.; Porter, W.P. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conserv. Lett. 2010, 3, 203–213. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. Biomod—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Hartley, S.; Harris, R.; Lester, P.J. Quantifying uncertainty in the potential distribution of an invasive species: Climate and the argentine ant. Ecol. Lett. 2006, 9, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Wenger, S.J.; Som, N.A.; Dauwalter, D.C.; Isaak, D.J.; Neville, H.M.; Luce, C.H.; Dunham, J.B.; Young, M.K.; Fausch, K.D.; Rieman, B.E. Probabilistic accounting of uncertainty in forecasts of species distributions under climate change. Glob. Chang. Biol. 2013, 19, 3343–3354. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Barry, S.; Elith, J. Error and uncertainty in habitat models. J. Appl. Ecol. 2006, 43, 413–423. [Google Scholar] [CrossRef]
- Leitão, P.J.; Moreira, F.; Osborne, P.E. Effects of geographical data sampling bias on habitat models of species distributions: A case study with steppe birds in Southern Portugal. Int. J. Geogr. Inf. Sci. 2011, 25, 439–454. [Google Scholar] [CrossRef]
- Bystriakova, N.; Peregrym, M.; Erkens, R.H.; Bezsmertna, O.; Schneider, H. Sampling bias in geographic and environmental space and its effect on the predictive power of species distribution models. Syst. Biodivers. 2012, 10, 305–315. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with maxent using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evolut. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Hannemann, H.; Willis, K.J.; Macias-Fauria, M. The devil is in the detail: Unstable response functions in species distribution models challenge bulk ensemble modelling. Glob. Ecol. Biogeogr. 2016, 25, 26–35. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.E.; DiTommaso, A.; Wang, R.L.; Liang, K.M. Predicting the potential distribution of lantana camara l. Under rcp scenarios using isi-mip models. Clim. Chang. 2015, 134, 193–208. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Innes, J.; Nitschke, C.; Kang, H. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia–pacific region. For. Ecol. Manag. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F. Package ‘Biomod2’: Ensemble Platform for Species Distribution Mdoeling, R Package Version 3.3-7. Available online: https://cran.r-project.org/web/packages/biomod2/index.html (accessed on 1 March 2016).
- Kong, W.-S.; Watts, D. The Plant Geography of Korea with an Emphasis on the Alpine Zones; Kluwer Academic: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Kim, K.; Jeong, G.Y. Factors influencing natural occurrence of fluoride-rich groundwaters: A case study in the southeastern part of the korean peninsula. Chemosphere 2005, 58, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.; Kong, W.-S.; Kim, C.-K. Distribution of evergreen broad-leaved plants and climate factors. J. Korean Geogr. Soc. 2001, 36, 247–257. [Google Scholar]
- Yun, J.-H.; Nakao, K.; Kim, J.-H.; Kim, S.-Y.; Park, C.-H.; Lee, B.-Y. Habitat prediction and impact assessment of Neolitsea sericea (Blume) Koidz. under Climate Change in Korea. J. Environ. Impact Assess. 2014, 23, 101–111. [Google Scholar] [CrossRef]
- Yun, J.-H.K.; Jung, H.; Oh, K.-H.; Lee, B.-Y. Distributional change and climate condition of warm-temperate evergreen broad-leaved trees in Korea. Korean J. Environ. Ecol. 2011, 25, 47–56. [Google Scholar]
- Pachauri, R.K.; Allen, M.R.; Barros, V.; Broome, J.; Cramer, W.; Christ, R.; Church, J.; Clarke, L.; Dahe, Q.; Dasgupta, P. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Botts, E.A.; Erasmus, B.F.; Alexander, G.J. Small range size and narrow niche breadth predict range contractions in South African frogs. Glob. Ecol. Biogeogr. 2013, 22, 567–576. [Google Scholar] [CrossRef]
- Koo, K. Distribution of Evergreen Broad-Leaved Trees and Climate Factors in the Korean Peninsula. Master’s Thesis, Kyung Hee University, Seoul, Korea, 2000. [Google Scholar]
- Lee, W.C.Y.; Yang, J. Plant Geography with Special Reference to Korea; Kangwon National University Press: Chooncheon, Korea, 2002; p. 412. [Google Scholar]
- Koo, K.A.; Kong, W.-S.; Nibbelink, N.P.; Hopkinson, C.S.; Lee, J.H. Potential effects of climate change on the distribution of cold-tolerant evergreen broadleaved woody plants in the Korean peninsula. PLoS ONE 2015, 10, e0134043. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic Predictors for Supporting Ecological Applications in the Conterminous United States; US Geological Survey Data Series; USGS: Reston, VA, USA, 2012; Volume 691.
- Araújo, M.B.; Whittaker, R.J.; Ladle, R.J.; Erhard, M. Reducing uncertainty in projections of extinction risk from climate change. Glob. Ecol. Biogeogr. 2005, 14, 529–538. [Google Scholar] [CrossRef]
- Diniz-Filho, J.A.F.; Mauricio, B.L.; Fernando, R.T.; Loyola, R.D.; Hof, C.; Nogués-Bravo, D.; Araújo, M.B. Partitioning and mapping uncertainties in ensembles of forecasts of species turnover under climate change. Ecography 2009, 32, 897–906. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; CRC Press: Boca Raton, FL, USA, 1989; Volume 37. [Google Scholar]
- Ridgeway, G. The state of boosting. Comput. Sci. Stat. 1999, 31, 172–181. [Google Scholar]
- Hastie, T.J.; Tibshirani, R.J. Generalized Additive Models; CRC Press: Boca Raton, FL, USA, 1990; Volume 43. [Google Scholar]
- Breiman, L.; Friedman, J.; Stone, C.J.; Olshen, R.A. Classification and Regression Trees; CRC Press: Boca Raton, FL, USA, 1984. [Google Scholar]
- Ripley, B.D. Pattern Recognition and Neural Networks; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Busby, J. Bioclim—A bioclimate analysis and prediction system. In Nature Conservation: Cost Effective Biological Surveys and Data Analysis; Margules, C.R., Austin, M.P., Eds.; CSIRO: Melbourne, Australia, 1991; pp. 64–68. [Google Scholar]
- Hastie, T.; Tibshirani, R.; Buja, A. Flexible discriminant analysis by optimal scoring. J. Am. Stat. Assoc. 1994, 89, 1255–1270. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Friedman, J.H. Multivariate adaptive regression splines. Ann. Stat. 1991, 19, 1–67. [Google Scholar] [CrossRef]
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. In Synthesis. American Museum of Natural History; American Museum of Natural History: New York, NY, USA, 2007; Volume 3, pp. 54–89. [Google Scholar]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Jones, A.W.; Dalton, C.M.; Stowe, E.S.; Post, D.M. Contribution of declining anadromous fishes to the reproductive investment of a common piscivorous seabird, the double-crested cormorant (Phalacrocorax auritus). Auk 2010, 127, 696–703. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Ruete, A.; Leynaud, G.C. Goal-Oriented Evaluation of Species Distribution Models’ Accuracy and Precision: True Skill Statistic Profile and Uncertainty Maps. PeerJ PrePrints 2015, 3, e1208v1. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Hanberry, B.; He, H. Prevalence, statistical thresholds, and accuracy assessment for species distribution models. Web Ecol. 2013, 13, 13–19. [Google Scholar] [CrossRef]
- Moon, H.-I. Antidiabetic effects of dendropanoxide from leaves of Dendropanax morbifera Leveille in normal and streptozotocin-induced diabetic rats. Hum. Exp. Toxicol. 2011, 30, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hon, J.; Yi, J.; Chun, Y.; Le, J. Characteristic of seed germination of Dendropanax morifera according to temperaure and ligh factors for estiamting change of habitat area in global warming. Clim. Res. 2013, 8, 143–151. [Google Scholar] [CrossRef]
- Kim, S.; Jang, Y.; Han, J.; Chung, H.; Lee, S.; Cho, K. Genetic variation and population structure of dendropanax morbifera lev.(araliaceae) in korea. Silvae Genet. 2006, 55, 7–13. [Google Scholar] [CrossRef]
- Choi, S.; Yoon, J.; Shin, H. Germination and growth characteristics of dendropanax morbifera lev. By sowing methods and shading conditions. Planta Med. 2013, 79, PN122. [Google Scholar] [CrossRef]
- Sakai, A.; Weiser, C. Freezing resistance of trees in North America with reference to tree regions. Ecology 1973, 54, 118–126. [Google Scholar] [CrossRef]
- Wang, X.P.; Tang, Z.Y.; Fang, J.Y. Climatic control on forests and tree species distribution in the forest region of northeast china. J. Integr. Plant Biol. 2006, 48, 778–789. [Google Scholar] [CrossRef]
- Kira, T. Forest ecosystems of east and Southeast Asia in a global perspective. Ecol. Res. 1991, 6, 185–200. [Google Scholar] [CrossRef]
- Da, L.-J.; Kang, M.-M.; Song, K.; Shang, K.-K.; Yang, Y.-C.; Xia, A.-M.; Qi, Y.-F. Altitudinal zonation of human-disturbed vegetation on mt. Tianmu, Eastern China. Ecol. Res. 2009, 24, 1287. [Google Scholar] [CrossRef]
- Chiu, C.-A.; Chiou, C.-R.; Lin, J.-R.; Lin, P.-H.; Lin, C.-T. Coldness index does not indicate the upper limit of evergreen broad-leaved forest on a subtropical island. J. For. Res. 2014, 19, 115–124. [Google Scholar] [CrossRef]
- Chen, X.; Wang, L.; Inouye, D. Delayed response of spring phenology to global warming in subtropics and tropics. Agric. For. Meteorol. 2017, 234, 222–235. [Google Scholar] [CrossRef]
- Luedeling, E.; Girvetz, E.H.; Semenov, M.A.; Brown, P.H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 2011, 6, e20155. [Google Scholar] [CrossRef] [PubMed]
- Moyes, A.B.; Germino, M.J.; Kueppers, L.M. Moisture rivals temperature in limiting photosynthesis by trees establishing beyond their cold-edge range limit under ambient and warmed conditions. New Phytol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Niu, S.; Wang, S. Precipitation regulates plant gas exchange and its long-term response to climate change in a temperate grassland. J. Plant Ecol. 2016, 9, 531–541. [Google Scholar] [CrossRef]
- Niu, S.; Luo, Y.; Li, D.; Cao, S.; Xia, J.; Li, J.; Smith, M.D. Plant growth and mortality under climatic extremes: An overview. Environ. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D.; Walters, S.M. Plant Variation and Evolution; Cambridge University: Cambridge, UK, 1997. [Google Scholar]
- Hamrick, J.L. Response of forest trees to global environmental changes. For. Ecol. Manag. 2004, 197, 323–335. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Sork, V.; Aitken, S.; Dyer, R.; Eckert, A.; Legendre, P.; Neale, D. Putting the landscape into the genomics of trees: Approaches for understanding local adaptation and population responses to changing climate. Tree Genet. Genomes 2013, 9, 901–911. [Google Scholar] [CrossRef]
- Graham, C.H.; Ron, S.R.; Santos, J.C.; Schneider, C.J.; Moritz, C. Integrating phylogenetics and environmental niche models to explore speciation mechanisms in dendrobatid frogs. Evolution 2004, 58, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Hereford, J. A quantitative survey of local adaptation and fitness trade-offs. Am. Nat. 2009, 173, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Grassein, F.; Lavorel, S.; Till-Bottraud, I. The importance of biotic interactions and local adaptation for plant response to environmental changes: Field evidence along an elevational gradient. Glob. Chang. Biol. 2014, 20, 1452–1460. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, R.J.; Wong, P.P.; Burkett, V.; Codignotto, J.; Hay, J.; McLean, R.; Ragoonaden, S.; Woodroffe, C.D.; Abuodha, P.; Arblaster, J. Coastal Systems and Low-Lying Areas; IPCC: Geneva, Switzerland, 2007. [Google Scholar]
- Mendoza-González, G.; Martínez, M.L.; Rojas-Soto, O.R.; Vázquez, G.; Gallego-Fernández, J.B. Ecological niche modeling of coastal dune plants and future potential distribution in response to climate change and sea level rise. Glob. Chang. Biol. 2013, 19, 2524–2535. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.R. Modernization, development and underdevelopment: Reclamation of Korean tidal flats, 1950s–2000s. Ocean Coast. Manag. 2014, 102, 426–436. [Google Scholar] [CrossRef]
- Kim, Y.; Cho, K. Sea level rise around Korea: Analysis of tide gauge station data with the ensemble empirical mode decomposition method. J. Hydro-Environ. Res. 2016, 11, 138–145. [Google Scholar] [CrossRef]
- Park, S.U.; Koo, K.A.; Kong, W.-S. Potential impact of climate change on distribution of warm temperate evergreen broad-leaved trees in the Korean peninsula. J. Korean Geogr. Soc. 2016, 51, 201–217. [Google Scholar]



| (a) | ||
| Scientific Name | # of Presence (Absence) Data | |
| Castanopsis cuspidata (Thunb.) Schottky | 53 (669) | |
| Kadsura japonica (L.) Dunal | 63 (659) | |
| Neolitsea sericea (Blume) Koidz. | 86 (636) | |
| Pittosporum tobira (Thunb.) W.T. Aiton | 120 (602) | |
| Raphiolepis indica var. umbellata (Thunb.) Ohashi | 80 (642) | |
| Ilex integra Thunb. | 59 (663) | |
| Eurya emarginata (Thunb.) Makino | 71 (651) | |
| Dendropanax morbifera H. Lév. | 48 (674) | |
| (b) | ||
| Bioclimatic Variables (BIO) | Definition of Variables | |
| BIO1 | Annual Mean Temperature | |
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp-min temp)) | |
| BIO12 | Annual Precipitation | |
| BIO13 | Precipitation of Wettest Month | |
| (c) | ||
| GCM | Code | Institution |
| GISS-E2-R | GSt | NASA Goddard Institute for Space Studies USA |
| HadGEM2-AO | HD | UK Met Office Hadley Centre |
| HadGEM2-ES | HE | UK Met Office Hadley Centre |
| MIROC-ESM-CHEM | MI | University of Tokyo, National Institute for Environmental Studies, and Japan Agency for Marine-Earth Science and Technology |
| MRI-CGCM3 | MG | University of Tokyo, National Institute for Environmental Studies, and Japan Agency for Marine-Earth Science and Technology |
| Species Name | TSS | Threshold | Sensitivity | Specificity |
|---|---|---|---|---|
| Castanopsis cuspidata | 0.923 | 0.111 | 1.000 | 0.921 |
| Kadsura japonica | 0.866 | 0.168 | 0.917 | 0.949 |
| Neolitsea sericea | 0.898 | 0.183 | 0.980 | 0.917 |
| Pittoporum tobira | 0.860 | 0.098 | 0.968 | 0.890 |
| Raphiolepis indica var. umbellata | 0.932 | 0.085 | 1.000 | 0.932 |
| Ilex integra | 0.787 | 0.190 | 0.875 | 0.912 |
| Eurya emarginata | 0.880 | 0.229 | 0.921 | 0.959 |
| Dendropanax morbifera | 0.868 | 0.115 | 0.968 | 0.900 |
| Species Name | 2050 | 2070 |
|---|---|---|
| Castanopsis cuspidata | 41 | 76 |
| Pittosporum tobira | 261 | 390 |
| Raphiolepis indica var. umbellata | 147 | 232 |
| Eurya emarginata | 62 | 106 |
| Kadsura japonica | −12 | −7 |
| Neolitsea sericea | −52 | −47 |
| Ilex integra | −51 | −53 |
| Dendropanax morbifera | −90 | −89 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koo, K.A.; Park, S.U.; Seo, C. Effects of Climate Change on the Climatic Niches of Warm-Adapted Evergreen Plants: Expansion or Contraction? Forests 2017, 8, 500. https://doi.org/10.3390/f8120500
Koo KA, Park SU, Seo C. Effects of Climate Change on the Climatic Niches of Warm-Adapted Evergreen Plants: Expansion or Contraction? Forests. 2017; 8(12):500. https://doi.org/10.3390/f8120500
Chicago/Turabian StyleKoo, Kyung Ah, Seon Uk Park, and Changwan Seo. 2017. "Effects of Climate Change on the Climatic Niches of Warm-Adapted Evergreen Plants: Expansion or Contraction?" Forests 8, no. 12: 500. https://doi.org/10.3390/f8120500
APA StyleKoo, K. A., Park, S. U., & Seo, C. (2017). Effects of Climate Change on the Climatic Niches of Warm-Adapted Evergreen Plants: Expansion or Contraction? Forests, 8(12), 500. https://doi.org/10.3390/f8120500