Traits and Resource Use of Co-Occurring Introduced and Native Trees in a Tropical Novel Forest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Castilla elastica—A Dominant Introduced Tree
2.2. Study Area
2.3. Sampling
2.4. Light Response Curves and Leaf Harvesting Protocol
2.5. Samples Processing
2.6. Statistical Analyses
3. Results
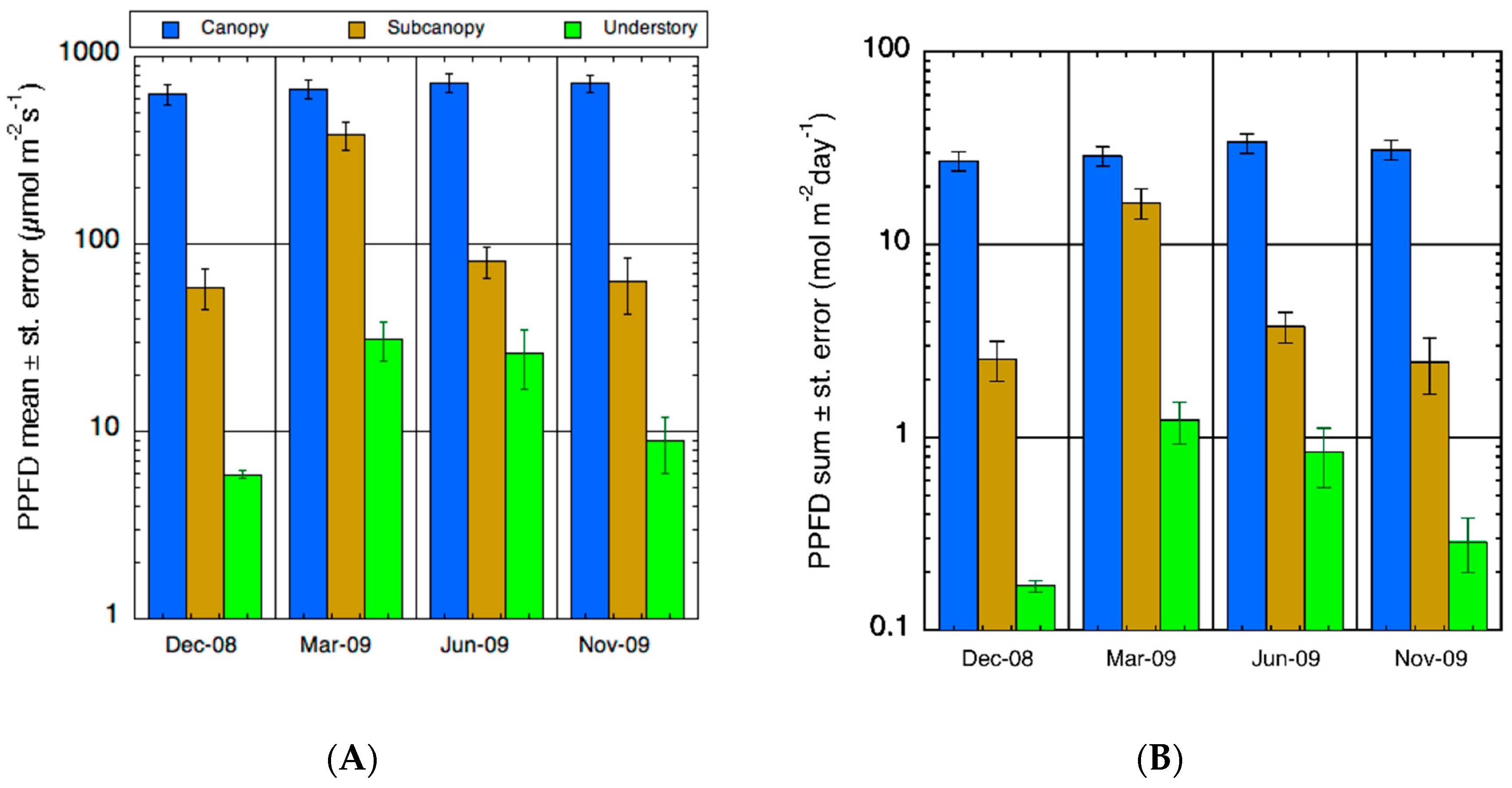
3.1. Environmental Conditions
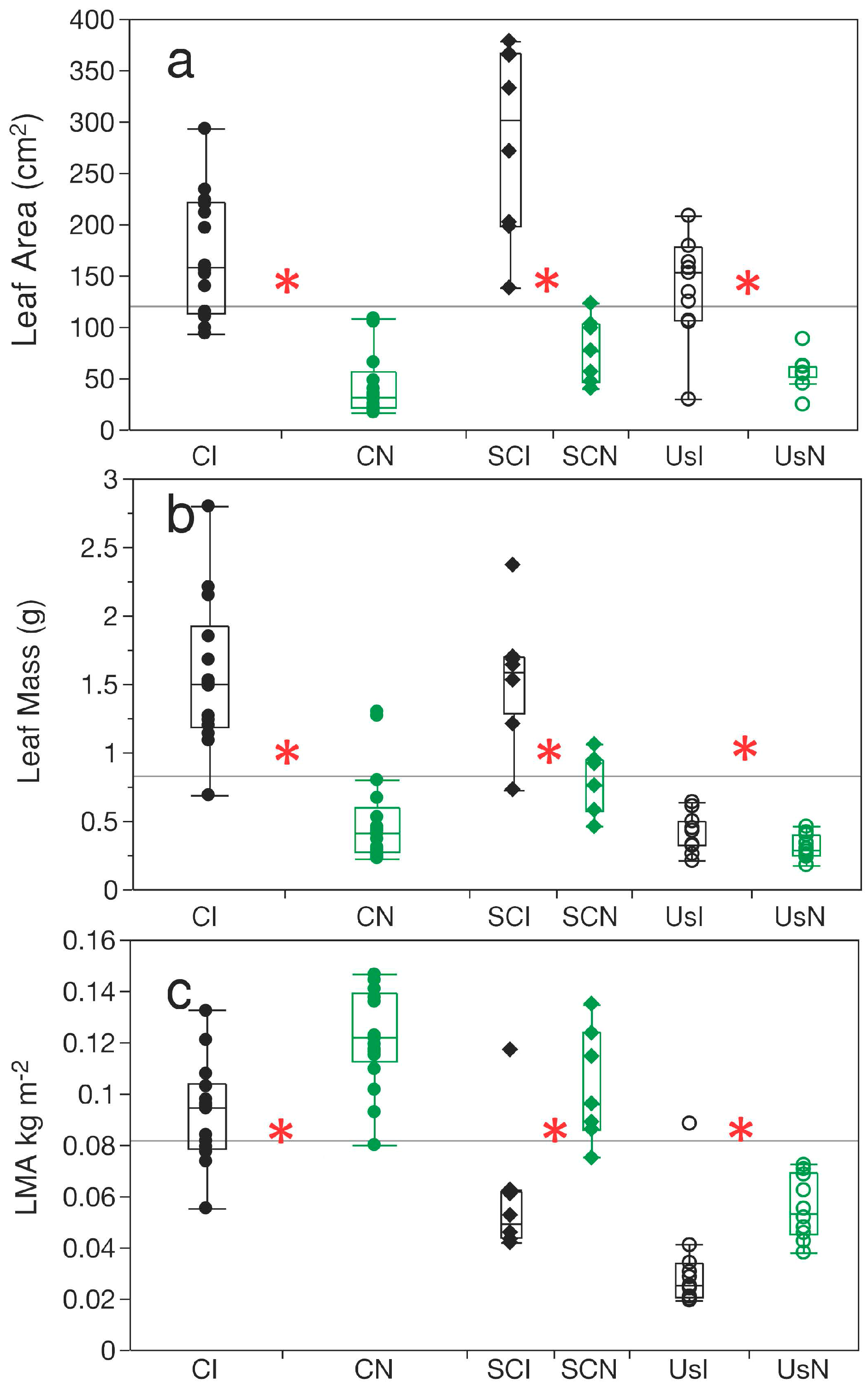
3.2. Leaf Dimensions
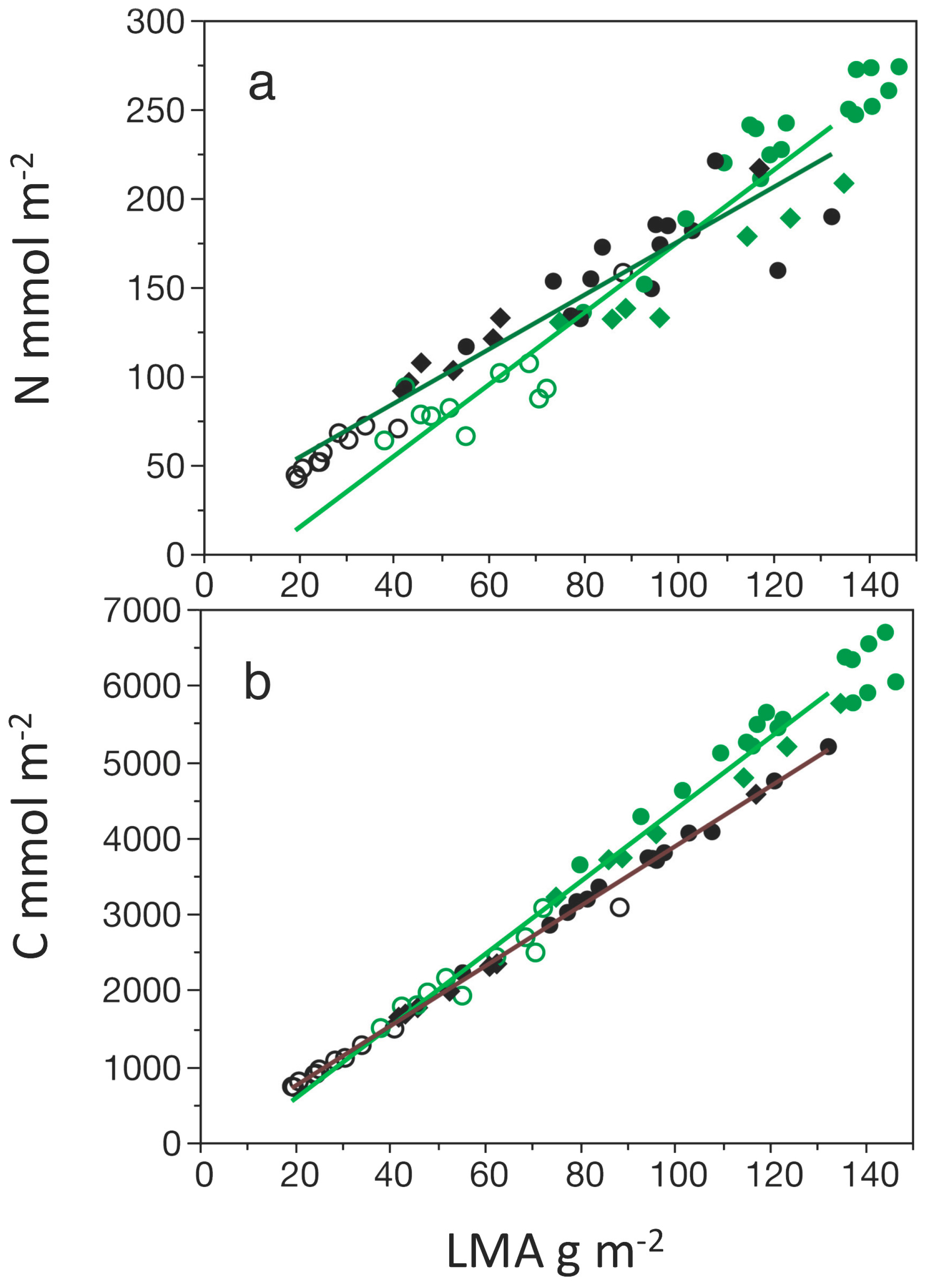
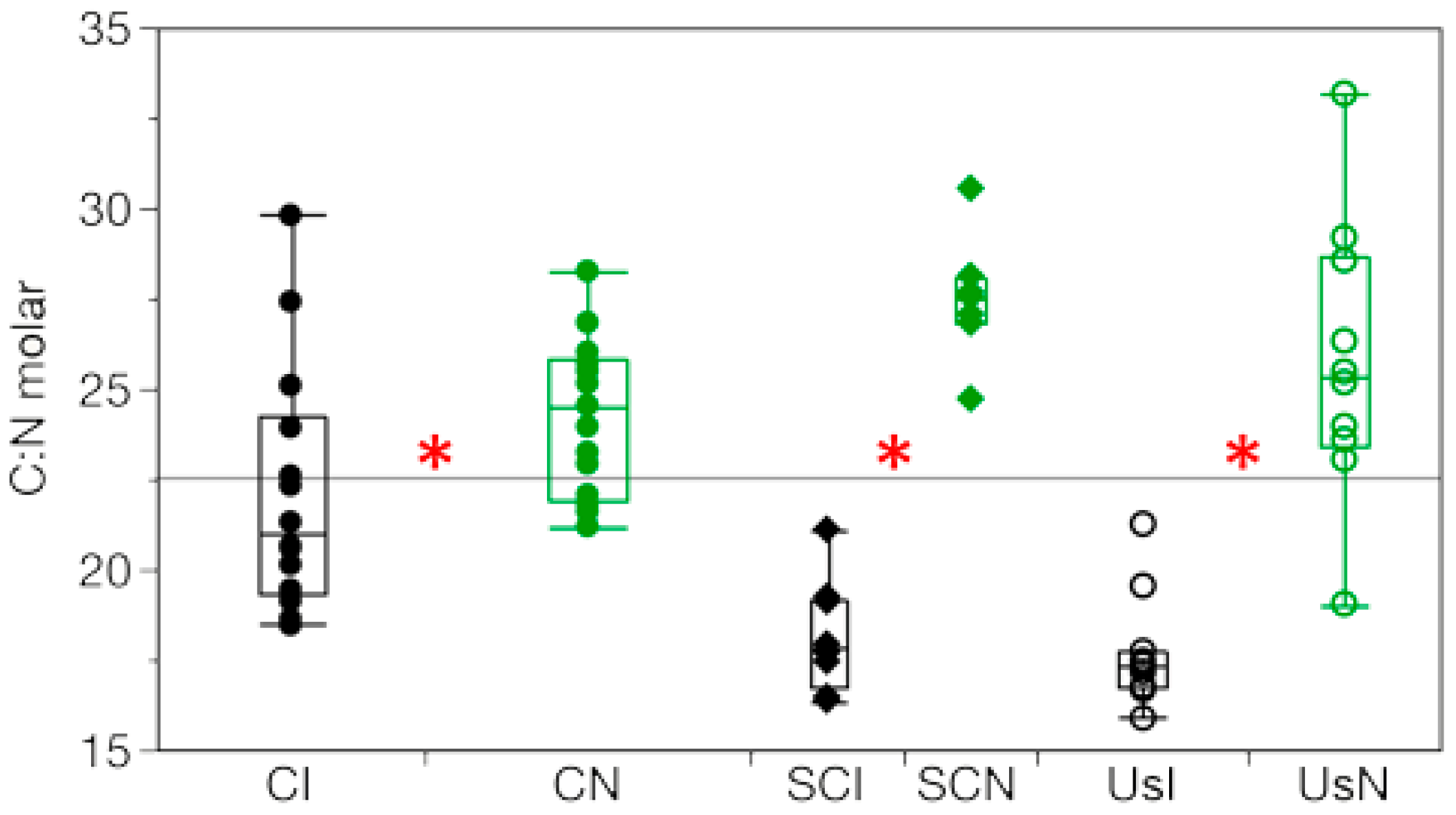
3.3. Concentration of C and N, and C:N Ratios
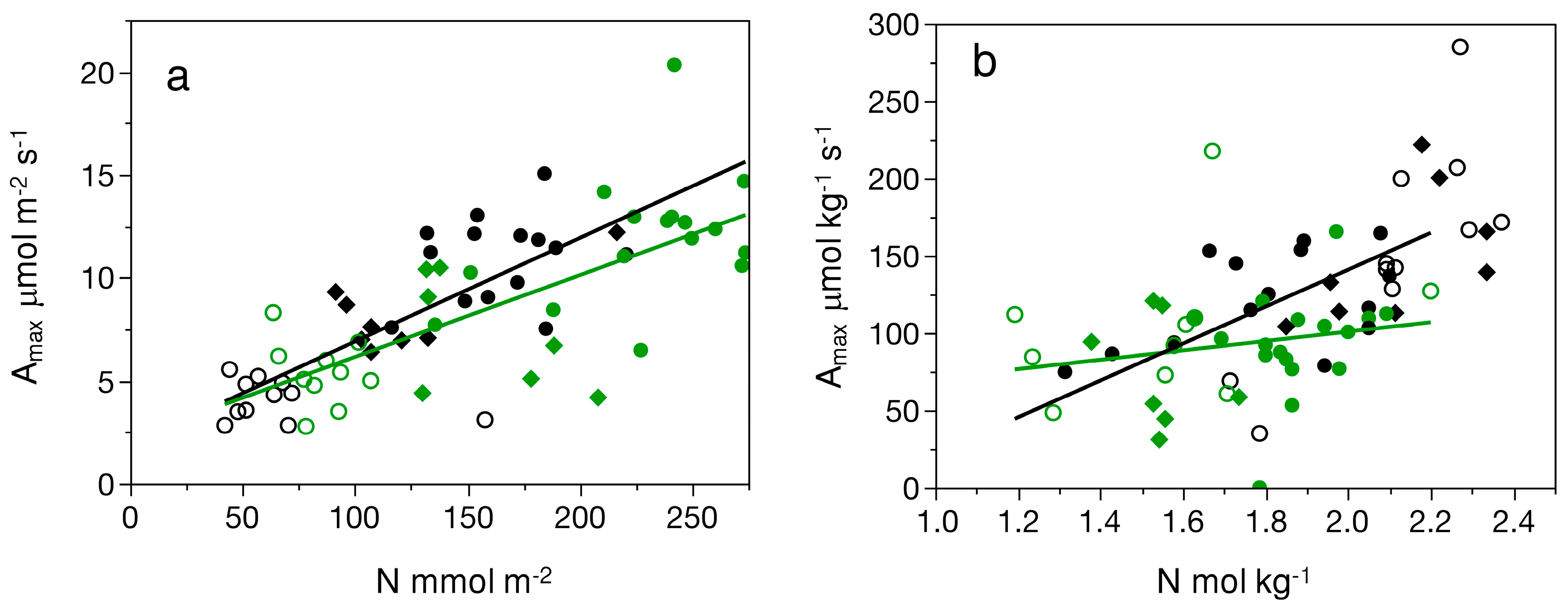
3.4. Comparisons Among Photosynthetic Capacity and Other Physiological Traits
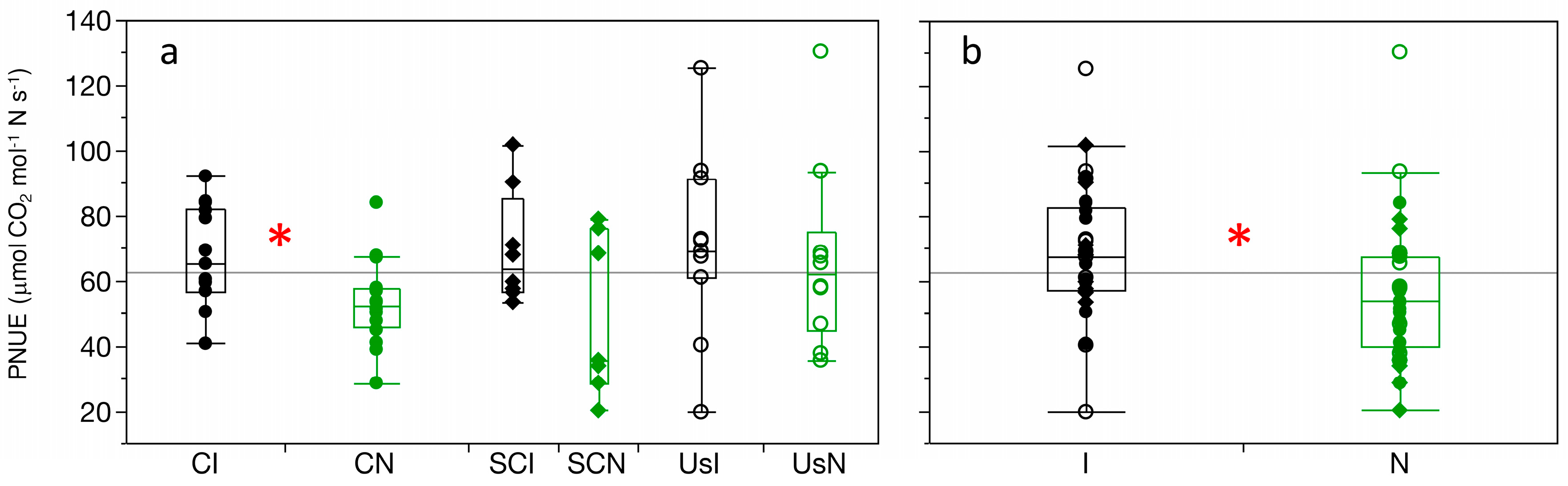
3.5. Photosynthetic Nitrogen Use-Efficiency
4. Discussion
4.1. Castilla elastica Influences the Irradiance Below the Canopy
4.2. Similar Photosynthetic Capacity and Water Use Among Species
4.3. Nitrogen Use-Efficiency and Leaf Area Are Advantages for Castilla elastica
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Malhi, Y.; Gardner, T.A.; Goldsmith, G.R.; Silman, M.R.; Zelazowski, P. Tropical forests in the Anthropocene. Annu. Rev. Environ. Resour. 2014, 39, 125–159. [Google Scholar] [CrossRef]
- Lugo, A.E.; Helmer, E. Emerging forests on abandoned land: Puerto Rico’s new forests. For. Ecol. Manag. 2004, 190, 145–161. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Arico, S.; Aronson, J.; Baron, J.S.; Bridgewater, P.; Cramer, V.A.; Epstein, P.R.; Ewel, J.J.; Klink, C.A.; Lugo, A.E.; et al. Novel ecosystems: Theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 2006, 15, 1–7. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.S.; Hall, C. Novel Ecosystems: Intervening in the New Ecological World Order, 1st ed.; John Wiley & Sons Ltd.: Chichester, UK, 2013. [Google Scholar]
- Higgs, E. Novel and designed ecosystems. Restor. Ecol. 2017, 25, 8–13. [Google Scholar] [CrossRef]
- Corlett, R.T. The Anthropocene concept in ecology and conservation. Trends Ecol. Evol. 2015, 30, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Alpert, P. The advantages and disadvantages of being introduced. Biol. Invasions 2006, 8, 1523–1534. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Pattison, R.R.; Goldstein, G.; Ares, A. Growth, biomass allocation and photosynthesis of invasive and native Hawaiian rainforest species. Oecologia 1998, 117, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Baruch, Z.; Goldstein, G. Leaf construction cost, nutrient concentration, and net CO2 assimilation of native and invasive species in Hawaii. Oecologia 1999, 121, 183–192. [Google Scholar] [CrossRef] [PubMed]
- McDowell, S.C.L. Photosynthetic characteristics of invasive and noninvasive species of Rubus (Rosaceae). Am. J. Bot. 2002, 89, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.H. A comparison of invasive and noninvasive dayflowers (Commelinaceae) across experimental nutrient and water gradients. Divers. Distrib. 2004, 10, 387–397. [Google Scholar] [CrossRef]
- Leicht Young, S.A.; Silander, J.A., Jr.; Latimer, A.M. Comparative performance of invasive and native Celastrus species across environmental gradients. Oecologia 2007, 154, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L. Differences in plasticity between invasive and native plants from a low resource environment. J. Ecol. 2008, 96, 1162–1173. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Llusià, J.; Owen, S.M.; Carnicer, J.; Giambelluca, T.W.; Rezende, E.L.; Waite, M.; Niinemets, Ü. Faster returns on “leaf economics” and different biogeochemical niche in invasive compared with native plant species. Glob. Chang. Biol. 2010, 16, 2171–2185. [Google Scholar] [CrossRef]
- Cavaleri, M.A.; Sack, L. Comparative water use of native and invasive plants at multiple scales: A global meta-analysis. Ecology 2010, 91, 2705–2715. [Google Scholar] [CrossRef] [PubMed]
- Lamarque, L.J.; Delzon, S.; Lortie, C.J. Tree invasions: A comparative test of the dominant hypotheses and functional traits. Biol. Invasions 2011, 13, 1969–1989. [Google Scholar] [CrossRef]
- Whitmore, T.C. Canopy gaps and the two major groups of forest trees. Ecology 1989, 70, 536–538. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Gemmell, N.J.; Rand, T.A.; Ewers, R.M. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol. Evol. 2007, 22, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Fonseca da Silva, J. Dynamics of novel forests of Castilla elastica in Puerto Rico: From species to ecosystems. Ecol. Evol. 2015, 5, 3299–3311. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M. Biological invasions and ecosystem processes: Towards an integration of population biology and ecosystem studies. Oikos 1990, 57, 7–13. [Google Scholar] [CrossRef]
- Funk, J.L.; Vitousek, P.M. Resource-use efficiency and plant invasion in low-resource systems. Nature 2007, 446, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Heard, M.J.; Sax, D.F. Coexistence between native and exotic species is facilitated by asymmetries in competitive ability and susceptibility to herbivores. Ecol. Lett. 2013, 16, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Fonseca da Silva, J. Species Composition, Diversity and Structure of Novel Forests of Castilla elastica in Puerto Rico. Trop. Ecol. 2014, 55, 231–244. [Google Scholar]
- Francis, J.K.; Liogier, H.A. Naturalized Exotic Tree Species in Puerto Rico; General Technical Report SO-82; USDA Forest Service: New Orleans, LA, USA, 1991.
- Sautu, A.; Baskin, J.M.; Baskin, C.C.; Condit, R. Studies on the seed biology of 100 native species of trees in a seasonal moist tropical forest, Panama, Central America. For. Ecol. Manag. 2006, 234, 245–263. [Google Scholar] [CrossRef]
- Ewel, J.; Whitmore, J.L. The Ecological life Zones of Puerto Rico and the U.S. Virgin Islands; Forest Service Research Paper ITF-18; USDA Forest Service, International Institute of Tropical Forestry: Río Piedras, Puerto Rico, 1973.
- Viera-Martínez, C.A.; Abelleira Martínez, O.J.M.; Lugo, A.E. Estructura y química del suelo en un bosque de Castilla elastica en el Carso del norte de Puerto Rico: Resultados de una calicata. Acta Cient. 2008, 22, 29–35. [Google Scholar]
- Axelrod, F.S. A Systematic Vademecum to the Vascular Plants of Puerto Rico; Botanical Research Institute of Texas: Fort Worth, TX, USA, 2011. [Google Scholar]
- Peek, M.S.; Russek-Cohen, E.; Wait, D.A.; Forseth, I.N. Physiological response curve analysis using nonlinear mixed models. Oecologia 2002, 132, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.C.; Hughes, A.P. Plant growth and the aerial environment. I. Effect of artificial shading on Impatiens parviflora. New Phytol. 1961, 60, 150–180. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A.; Rich, P.M.; Weiss, S.; Oberbauer, S.F. Landscape-scale evaluation of understory light and canopy structures: Methods and application in a Neotropical lowland rain forest. Can. J. For. Res. 1996, 26, 747–757. [Google Scholar] [CrossRef]
- Sakai, S. Thrips pollination of androdioecious Castilla elastica (Moraceae) in a seasonal tropical forest. Am. J. Bot. 2001, 88, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Mozdzer, T.J.; Zieman, J.C. Ecophysiological differences between genetic lineages facilitate the invasion of non-native Phragmites australis in North American Atlantic coast wetlands. J. Ecol. 2010, 98, 451–458. [Google Scholar] [CrossRef]
- Kitajima, K.; Mulkey, S.S.; Wright, S.J. Seasonal leaf phenotypes in the canopy of a tropical dry forest: Photosynthetic characteristics and associated traits. Oecologia 1997, 109, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S.; Uhl, C. Photosynthesis-nitrogen relations in Amazonian tree species. Oecologia 1994, 97, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Palacio López, K.; Gianoli, E. Invasive plants do not display greater phenotypic plasticity than their native or non-invasive counterparts: A meta-analysis. Oikos 2011, 120, 1393–1401. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating resources in plant communities: A general theory of invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Grotkopp, E.; Rejmánek, M.; Rost, T.L. Toward a causal explanation of plant invasiveness: Seedling growth and life-history strategies of 29 pine (Pinus) species. Am. Nat. 2002, 159, 396–419. [Google Scholar] [CrossRef] [PubMed]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Lake, J.C.; Leishman, M.R. Invasion success of exotic plants in natural ecosystems: The role of disturbance, plant attributes and freedom from herbivores. Biol. Conserv. 2004, 117, 215–226. [Google Scholar] [CrossRef]
- Hamilton, M.A.; Murray, B.R.; Cadotte, M.W.; Hose, G.C.; Baker, A.C.; Harris, C.J.; Licari, D. Life-history correlates of plant invasiveness at regional and continental scales. Ecol. Lett. 2005, 8, 1066–1074. [Google Scholar] [CrossRef]
- Leishman, M.R.; Thomson, V.P. Experimental evidence for the effects of additional water, nutrients and physical disturbance on invasive plants in low fertility Hawkesbury Sandstone soils, Sydney, Australia. J. Ecol. 2005, 93, 38–49. [Google Scholar] [CrossRef]
- Grotkopp, E.; Rejmánek, M. High seedling relative growth rate and specific leaf area are traits of invasive species: Phylogenetically independent contrasts of woody angiosperms. Ecology 2007, 94, 526–532. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Forest Stratum |
|---|---|---|
| Casearia guianensis (Aubl.) Urban N | Flacourtiaceae | Us |
| Casearia sylvestris Sw. N | Flacourtiaceae | SC |
| Castilla elastica Sessé In | Moraceae | C, SC, Us |
| Chrysophyllum argenteum Jacques N | Sapotaceae | Us |
| Cordia alliodora (Ruiz & Pav.) N | Boraginaceae | C |
| Faramea occidentalis (L.) A. Rich N | Rubiaceae | Us |
| Guarea guidonia (L.) Sleumer N | Meliaceae | Us |
| Ocotea floribunda (Sw.) Mez N | Lauraceae | C |
| Ocotea leucoxylon (Sw.) Mez N | Lauraceae | C |
| Thouinia striata Radlk Ne | Sapindaceae | Us |
| Trichilia pallida Sw N | Meliaceae | Us |
| Amax | LCP | LSP | ||
|---|---|---|---|---|
| Stratum | µmol kg−1 s−1 | µmol m−2 s−1 | µmol m−2 s−1 | |
| Canopy | 104.4 ± 18.8 (31) | 11.3 ± 1.6 (34) | 22.6 ± 3.2 | 927 ± 57 |
| Subcanopy | 113.8 ± 25.4 (15) | 7.6 ± 1.7 (19) | 15.2 ± 3.5 | 775 ± 122 |
| Understory | 127.1 ± 42.6 (21) | 4.9 ± 1.0 (26) | 9.6 ± 1.9 | 275 ± 57 |
| χ2 Wilcoxon/Kruskal-Wallis | p = 0.462 | p < 0.0001 | p < 0.0001 | p < 0.0001 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca da Silva, J.; Medina, E.; Lugo, A.E. Traits and Resource Use of Co-Occurring Introduced and Native Trees in a Tropical Novel Forest. Forests 2017, 8, 339. https://doi.org/10.3390/f8090339
Fonseca da Silva J, Medina E, Lugo AE. Traits and Resource Use of Co-Occurring Introduced and Native Trees in a Tropical Novel Forest. Forests. 2017; 8(9):339. https://doi.org/10.3390/f8090339
Chicago/Turabian StyleFonseca da Silva, Jéssica, Ernesto Medina, and Ariel E. Lugo. 2017. "Traits and Resource Use of Co-Occurring Introduced and Native Trees in a Tropical Novel Forest" Forests 8, no. 9: 339. https://doi.org/10.3390/f8090339
APA StyleFonseca da Silva, J., Medina, E., & Lugo, A. E. (2017). Traits and Resource Use of Co-Occurring Introduced and Native Trees in a Tropical Novel Forest. Forests, 8(9), 339. https://doi.org/10.3390/f8090339