Regeneration of Native Forest Species in Mainland Portugal: Identifying Main Drivers



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Base Data
2.2. Data Pre-Treatments
2.3. Zero-Inflated Count Data Models
2.4. Model Fitting
- On the count component: G, N, GRAZI, UnderClear, SeedAvail (2 km), DensPp (10 km), DensEg (10 km), FireFreq, OMBRO, THERM, TRASP, SLOPE, SoilMoist (summer) and SPMType;
- On the zero inflation component: SoilMoist (summer), OMBRO, THERM and SeedAvail (10 km).
2.5. Variables Importance Value
3. Results
4. Discussion
4.1. Zero Inflation Component
4.2. Count Component
4.2.1. Q. robur
4.2.2. Q. pyrenaica
4.2.3. Q. broteroi
4.2.4. Q. suber
4.2.5. Q. rotundifolia
4.3. Selected Neighbourhoods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
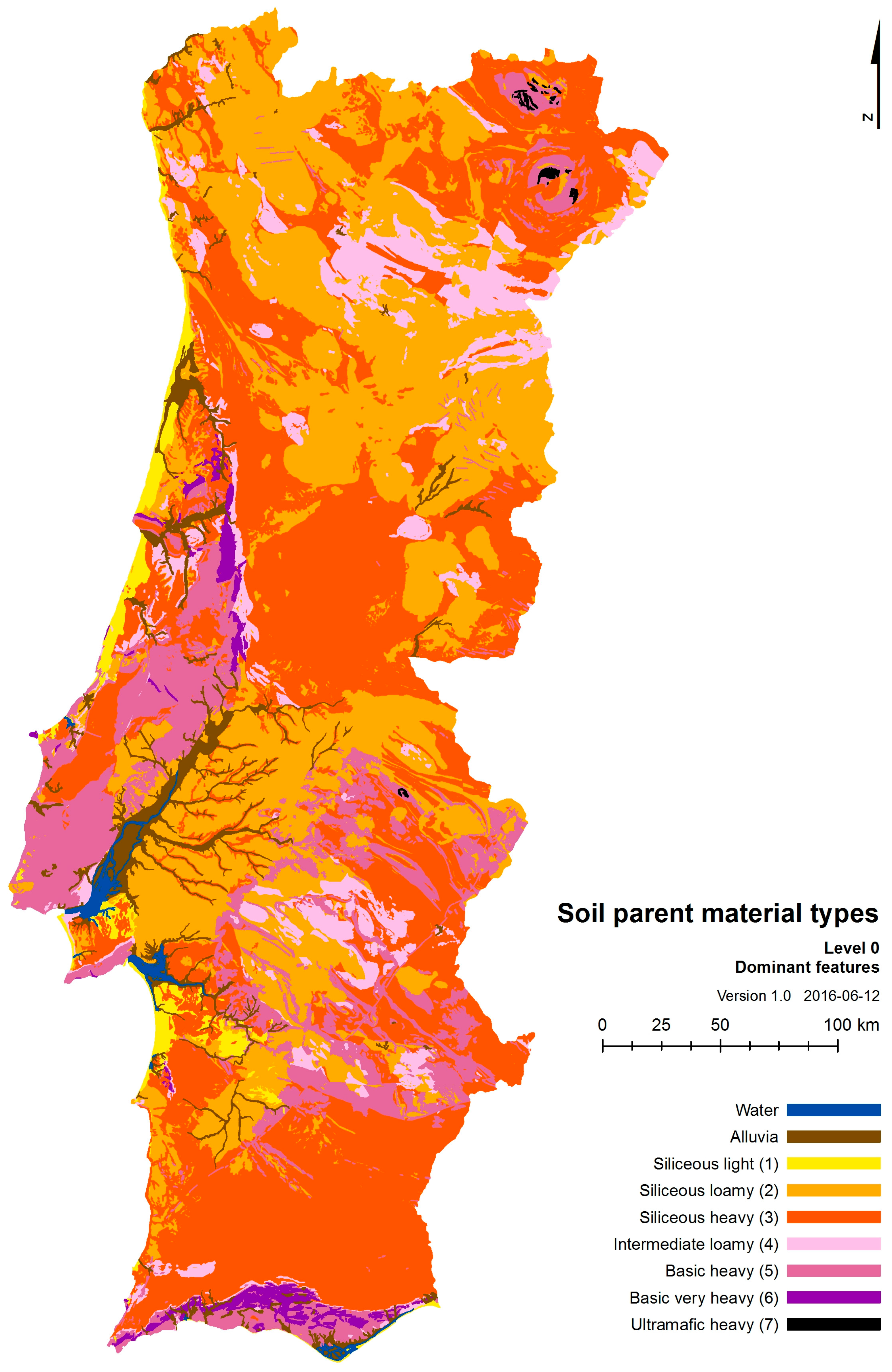
| Textural Classes | ||||
|---|---|---|---|---|
| Light (Sandy) | Loamy | Heavy (Silty or Clayish) | ||
| Base richness | Siliceous | siliceous light | siliceous loamy | siliceous heavy |
| Intermediate | intermediate loamy | |||
| Basic | basic heavy | |||
| Ultramafic | ultramafic heavy | |||
| Class | Relative Content in | Central Soil Texture | Rock Types/Formations | ||
|---|---|---|---|---|---|
| SiO2 | K | Ca | |||
| siliceous light | ++++ | - | - | very light (sandy) | Siliceous sand formations. |
| siliceous loamy | +++ | ++ | - | (light to) loamy | Chert (flint, radiolarite), siliceous/ferric sandstone, consolidated dunes, siliceous conglomerate, quartz syenite, quartz trachyte, acid tuff, granite, rhyolite, syenite, trachyte, siliceous gneiss, siliceous paragneiss, arkose, quartz, quartz monzonite, quartz latite, quartzite. |
| siliceous heavy | ++ | + | + | (loamy to) heavy | Siliceous schist, clayish conglomerate, greywacke, siliceous phyllite, clayish sandstone, turbidite, mica schist, clayish schist, slate, lydite rich metasediments, claystone, clay, mudstone (shale, argilite), metapelite, silt, silty phyllite, siltstone, ampelite, silty arkose, expansive clays. |
| intermediate loamy | + | - | ++ | (light to) loamy (to heavy) | Granodiorite, monzonite, latite, foid syenite, phonolite, tonalite, tonalitic gneiss, quartz monzodiorite, monzodiorite, siliceous schist/phyllite with carbonated or calc-silicate layers, calciturbidite, quartz diorite, diorite, quartz monzogabbro. |
| basic heavy | - | - | +++ | (loamy to) heavy | Carbonate sandstone, carbonate breccia/conglomerate, sandstone + marlstone, sandstone + limestone, andesite, quartz gabbro, monzogabbro, basaltic andesite, mafic lamprophyre, diabase/dolerite, gabbro, foid gabbro, basalt, tephrite, marlstone + sandstone, limestone + sandstone, amphibolite, limestone + marlstone, marble, limestone. |
| basic very heavy | - | - | +++ | very heavy | Marlstone + limestone, marlstone, dolomite rock. |
| ultramafic heavy | - - | - * | var. | heavy | Basanite (igneous), ultramafic lamprophyre, pyroxenite, serpentinite, peridotite, metaperidotite, dunite. |
- (i)
- boundaries between granitoid rocks and metasediments were sometimes recognizable in the orthoimages and particularly valuable in the centre and north of mainland Portugal.
- (ii)
- in the south, the transition between limestone and metasediments was often distinguishable in the orthoimages.
- (iii)
- ultramafic rocks, which are known to affect greatly the vegetation cover, allowed to add some correction points in the north-east of mainland Portugal, using the shift to other basic rocks.
- (iv)
- alluvial plains have usually dendritic shapes, matched to the easily recognizable river networks; additionally, their limits are also identifiable straightforwardly in orthoimages, as they often produce clear-cut geomorphologic transitions to the surrounding hills. Therefore, almost all the patches corresponding to alluvial sediments produced several georeferencing correction points.

References
- Food and Agriculture Organization of the United Nations. Global Forest Resources Assessment 2010: Main Report; FAO Forestry Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010; ISBN 978-92-5-106654-6. [Google Scholar]
- Bongiorno, S.F. Land use and summer bird populations in northwestern galicia, Spain. Ibis 1982, 124, 1–20. [Google Scholar] [CrossRef]
- Kwok, H.K.; Corlett, R.T. The bird communities of a natural secondary forest and a Lophostemon confertus plantation in Hong Kong, South China. For. Ecol. Manag. 2000, 130, 227–234. [Google Scholar] [CrossRef]
- Barlow, J.; Mestre, L.A.M.; Gardner, T.A.; Peres, C.A. The value of primary, secondary and plantation forests for Amazonian birds. Biol. Conserv. 2007, 136, 212–231. [Google Scholar] [CrossRef]
- Farwig, N.; Sajita, N.; Böhning-Gaese, K. Conservation value of forest plantations for bird communities in western Kenya. For. Ecol. Manag. 2008, 255, 3885–3892. [Google Scholar] [CrossRef]
- Calviño-Cancela, M.; Rubido-Bará, M.; van Etten, E.J.B. Do eucalypt plantations provide habitat for native forest biodiversity? For. Ecol. Manag. 2012, 270, 153–162. [Google Scholar] [CrossRef]
- Calviño-Cancela, M. Effectiveness of eucalypt plantations as a surrogate habitat for birds. For. Ecol. Manag. 2013, 310, 692–699. [Google Scholar] [CrossRef]
- Pina, J.P. Breeding bird assemblages in eucalyptus plantations in Portugal. Ann. Zool. Fenn. 1989, 26, 287–290. [Google Scholar]
- Proença, V.M.; Pereira, H.M.; Guilherme, J.; Vicente, L. Plant and bird diversity in natural forests and in native and exotic plantations in NW Portugal. Acta Oecol. Int. J. Ecol. 2010, 36, 219–226. [Google Scholar] [CrossRef]
- Williams, M. Deforesting the Earth: From Prehistory to Global Crisis: An Abridgment; University of Chicago Press: Chicago, IL, USA, 2006; ISBN 0-226-89947-0. [Google Scholar]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-scale patterns of Quercus ilex, Quercus suber and Quercus pyrenaica regeneration in central-western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Pons, X.; Ninyerola, M.; Vayreda, J.; Peñuelas, J. Large-scale recruitment limitation in Mediterranean pines: The role of Quercus ilex and forest successional advance as key regional drivers. Glob. Ecol. Biogeogr. 2014, 23, 371–384. [Google Scholar] [CrossRef]
- Benavides, R.; Escudero, A.; Coll, L.; Ferrandis, P.; Ogaya, R.; Gouriveau, F.; Penuelas, J.; Valladares, F. Recruitment patterns of four tree species along elevation gradients in Mediterranean mountains: Not only climate matters. For. Ecol. Manag. 2016, 360, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Benito-Garzón, M.; Ruiz-Benito, P.; Zavala, M.A. Interspecific differences in tree growth and mortality responses to environmental drivers determine potential species distributional limits in Iberian forests. Glob. Ecol. Biogeogr. 2013, 22, 1141–1151. [Google Scholar] [CrossRef]
- Stevens-Rumann, C.S.; Kemp, K.B.; Higuera, P.E.; Harvey, B.J.; Rother, M.T.; Donato, D.C.; Morgan, P.; Veblen, T.T. Evidence for declining forest resilience to wildfires under climate change. Ecol. Lett. 2017. [Google Scholar] [CrossRef] [PubMed]
- Mather, A.S.; Pereira, J.M.C. Transição Florestal e Fogo em Portugal. In Incêndios Florestais em Portugal: Caracterização, Impactes e Prevenção; Pereira, J.S., Pereira, J.M.C., Rego, F.C., Silva, J.M.N., da Silva, T.P., Eds.; ISAPress: Lisboa, Portugal, 2006; pp. 257–282. [Google Scholar]
- Louro, G.; do Monteiro, M.L.; Constantino, L.; Tomé, M.; Rego, F. Evolução do material lenhoso de pinheiro-bravo e eucalipto. Silva Lusit. 2010, 18, 133–149. [Google Scholar]
- Moreira, F.; Rego, F.C.; Ferreira, P.G. Temporal (1958–1995) pattern of change in a cultural landscape of northwestern Portugal: Implications for fire occurrence. Landsc. Ecol. 2001, 16, 557–567. [Google Scholar] [CrossRef]
- Costa, A.; Madeira, M.; Santos Lima, J. Is cork oak (Quercus suber L.) woodland loss driven by eucalyptus plantation? A case-study in southwestern Portugal. IFor. Biogeosci. For. 2014, 7, 193–203. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Loureiro, C.; Guiomar, N.; Pezzatti, G.B.; Manso, F.T.; Lopes, L. The dynamics and drivers of fuel and fire in the Portuguese public forest. J. Environ. Manag. 2014, 146, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, M.C.S.; Vasconcelos, M.J.; Pereira, J.M.C.; Dasgupta, N.; Alldredge, R.J.; Rego, F.C. Land cover type and fire in Portugal: Do fires burn land cover selectively? Landsc. Ecol. 2005, 20, 661–673. [Google Scholar] [CrossRef]
- Watt, A.S. On the causes of failure of natural regeneration in British oakwoods. J. Ecol. 1919, 7, 173–203. [Google Scholar] [CrossRef]
- Buckley, D.S.; Sharik, T.L.; Isebrands, J.G. Regeneration of northern red oak: Positive and negative effects of competitor removal. Ecology 1998, 79, 65–78. [Google Scholar] [CrossRef]
- Carnicer, J.; Barbeta, A.; Sperlich, D.; Coll, M.; Peñuelas, J. Contrasting trait syndromes in angiosperms and conifers are associated with different responses of tree growth to temperature on a large scale. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čater, M.; Batič, F. Groundwater and light conditions as factors in the survival of pedunculate oak (Quercus Robur L.) Seedlings. Eur. J. For. Res. 2006, 125, 419–426. [Google Scholar] [CrossRef]
- Latham, R. Co-occurring tree species change rank in seedling performance with resources varied experimentally. Ecology 1992, 73, 2129–2144. [Google Scholar] [CrossRef]
- Bazzaz, F.; Miao, S. Successional status, seed size and responses of tree seedlings to CO2, light and nutrients. Ecology 1993, 74, 104–112. [Google Scholar] [CrossRef]
- Canham, C.; Finzi, A.; Pacala, S.; Burbank, D. Causes and consequences of resource heterogeneity in forests: Interspecific variation in light transmission by canopy trees. Can. J. For. Res. Rev. 1994, 24, 337–349. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Pardos, J.A.; Gil, L.; Reich, P.B.; Aranda, I. Light response in seedlings of a temperate (Quercus petraea) and a sub-Mediterranean species (Quercus pyrenaica): Contrasting ecological strategies as potential keys to regeneration performance in mixed marginal populations. Plant Ecol. 2008, 195, 273–285. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Gómez, J.M.; Hódar, J.A.; Castro, J.; Baraza, E. Applying plant facilitation to forest restoration: A meta-analysis of the use of shrubs as nurse plants. Ecol. Appl. 2004, 14, 1128–1138. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R. Effect of Shrubs on Recruitment of Quercus-Douglasii and Quercus-Lobata. Ecology 1992, 73, 2118–2128. [Google Scholar] [CrossRef]
- Smit, C.; den Ouden, J.; Diaz, M. Facilitation of Quercus ilex recruitment by shrubs in Mediterranean open woodlands. J. Veg. Sci. 2008, 19, 193–200. [Google Scholar] [CrossRef]
- Vander Wall, S.B. The evolutionary ecology of nut dispersal. Bot. Rev. 2001, 67, 74–117. [Google Scholar] [CrossRef]
- Gómez, J.M. Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography 2003, 26, 573–584. [Google Scholar] [CrossRef]
- Tilghman, N. Impacts of white-tailed deer on forest regeneration in Northwestern Pennsylvania. J. Wildland Manag. 1989, 53, 524–532. [Google Scholar] [CrossRef]
- Hewitt, N.; Kellman, M. Tree seed dispersal among forest fragments: II. Dispersal abilities and biogeographical controls. J. Biogeogr. 2002, 29, 351–363. [Google Scholar] [CrossRef]
- Boiffin, J.; Aubin, I.; Munson, A.D. Ecological controls on post-fire vegetation assembly at multiple spatial scales in eastern North American boreal forests. J. Veg. Sci. 2015, 26, 360–372. [Google Scholar] [CrossRef]
- Coll, M.; Peñuelas, J.; Ninyerola, M.; Pons, X.; Carnicer, J. Multivariate effect gradients driving forest demographic responses in the Iberian Peninsula. For. Ecol. Manag. 2013, 303, 195–209. [Google Scholar] [CrossRef]
- Caldeira, M.C.; Ibáñez, I.; Nogueira, C.; Bugalho, M.N.; Lecomte, X.; Moreira, A.; Pereira, J.S. Direct and indirect effects of tree canopy facilitation in the recruitment of Mediterranean oaks. J. Appl. Ecol. 2014, 51, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Busing, R.T. Canopy cover and tree regeneration in old-growth cove forests of the Appalachian Mountains. Vegetatio 1994, 115, 19–27. [Google Scholar]
- Camisón, Á.; Miguel, R.; Marcos, J.L.; Revilla, J.; Tardáguila, M.Á.; Hernández, D.; Lakicevic, M.; Jovellar, L.C.; Silla, F. Regeneration dynamics of Quercus pyrenaica Willd. in the Central System (Spain). For. Ecol. Manag. 2015, 343, 42–52. [Google Scholar] [CrossRef]
- Lorimer, C.; Chapman, J.; Lambert, W. Tall understorey vegetation as a factor in the poor development of oak seedlings beneath mature stands. J. Ecol. 1994, 82, 227–237. [Google Scholar] [CrossRef]
- Hannon, G.E.; Bradshaw, R.; Emborg, J. 6000 years of forest dynamics in Suserup Skov, a seminatural Danish woodland. Glob. Ecol. Biogeogr. 2000, 9, 101–114. [Google Scholar] [CrossRef]
- Pausas, J.C.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin?—A review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Carrión, Y.; Kaal, J.; Antonio Lopez-Saez, J.; Lopez-Merino, L.; Martinez Cortizas, A. Holocene vegetation changes in NW Iberia revealed by anthracological and palynological records from a colluvial soil. Holocene 2010, 20, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Capelo, J.; Mesquita, S.; Costa, J.C.; Ribeiro, S.; Arsénio, P.; Neto, C.; Monteiro-Henriques, T.; Aguiar, C.; Honrado, J.; Espírito-Santo, D.; et al. A methodological approach to potential vegetation modeling using GIS techniques and phytosociological expert-knowledge: Application to mainland Portugal. Phytocoenologia 2007, 37, 399–415. [Google Scholar] [CrossRef]
- Costa, J.C.; Neto, C.; Aguiar, C.; Capelo, J.; Espírito Santo, M.D.; Honrado, J.; Pinto-Gomes, C.; Monteiro-Henriques, T.; Sequeira, M.; Lousã, M.; et al. Vascular plant communities in Portugal (Continental, the Azores and Madeira). Glob. Geobot. 2012, 2, 1–180. [Google Scholar] [CrossRef]
- Costa, J.C.; Monteiro-Henriques, T.; Neto, C.; Arsénio, P.; Aguiar, C. The application of the habitats directive in Portugal. Fitosociologia 2007, 44, 23–28. [Google Scholar]
- Eira, J.; Pinho, J.; Santos, C.; Buxo, J.; Godinho Ferreira, P.; Uva, J.; Moreira, J. Global Forest Resources Assessment 2010-Country Report-Portugal; Food and Agriculture Organization: Rome, Italy, 2010. [Google Scholar]
- Svenning, J.-C. A review of natural vegetation openness in north-western Europe. Biol. Conserv. 2002, 104, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.H.W.; Hannon, G.E.; Lister, A.M. A long-term perspective on ungulate-vegetation interactions. For. Ecol. Manag. 2003, 181, 267–280. [Google Scholar] [CrossRef]
- Mitchell, F.J.G. How open were European primeval forests? Hypothesis testing using palaeoecological data. J. Ecol. 2005, 93, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S.; Fernández, S.; González-Sampériz, P.; Gil-Romera, G.; Badal, E.; Carrión-Marco, Y.; López-Merino, L.; López-Sáez, J.A.; Fierro, E.; Burjachs, F. Expected trends and surprises in the Lateglacial and Holocene vegetation history of the Iberian Peninsula and Balearic Islands. Rev. Palaeobot. Palynol. 2010, 162, 458–475. [Google Scholar] [CrossRef] [Green Version]
- Direcção-Geral dos Recursos Florestais. Resultados do Inventário Florestal Nacional 2005/2006, 5a Revisão; Direcção-Geral dos Recursos Florestais: Lisboa, Portugal, 2007; p. 70. [Google Scholar]
- Aguiar, C. Flora e Vegetação da Serra de Nogueira e do Parque Natural de Montesinho. Ph.D. Thesis, Universidade Técnica de Lisboa-Instituto Superior de Agronomia, Universidade Técnica de Lisboa (TULisbon), Lisboa, Portugal, 2001. [Google Scholar]
- Monteiro-Henriques, T. Landscape and Phytosociology of the Paiva River’s Hydrographical Basin. Ph.D. Thesis, Instituto Superior de Agronomia, Universidade Técnica de Lisboa (TULisbon), Lisboa, Portugal, 2010. [Google Scholar]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling (Version 2.3-40). Available online: http://CRAN.R-project.org/package=raster (accessed on 28 December 2015).
- Monteiro-Henriques, T.; Martins, M.J.; Cerdeira, J.O.; Silva, P.; Arsénio, P.; Silva, Á.; Bellu, A.; Costa, J.C. Bioclimatological mapping tackling uncertainty propagation: Application to mainland Portugal. Int. J. Climatol. 2016, 36, 400–411. [Google Scholar] [CrossRef]
- Gavilán, R. The use of climatic parameters and indices in vegetation distribution. A case study in the Spanish Sistema Central. Int. J. Biometeorol. 2005, 50, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, A.; Reuter, H.I.; Nelson, A.; Guevara, E. Hole-Filled SRTM for the Globe Version 4. Available online: http://www.cgiar-csi.org/data/srtm-90m-digital-elevation-database-v4-1 (accessed on 9 April 2014).
- Evans, J. Topographic Radiation Index-ArcScripts. Available online: http://arcscripts.esri.com/details.asp?dbid=12643 (accessed on 12 April 2016).
- Tarboton, D.G. A New method for the determination of flow directions and contributing areas in grid digital elevation models. Water Resour. Res. 1997, 33, 309–319. [Google Scholar] [CrossRef]
- Tarboton, D.G. TAUDEM: Terrain Analysis Using Digital Elevation Model (Version 5.0.6). Available online: http://hydrology.usu.edu/taudem/taudem5/downloads5.0.html (accessed on 1 December 2013).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria (Version 3.0.1). Available online: http://www.R-project.org/ (accessed on 1 December 2013).
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Use R! Springer: New York, NY, USA, 2008; ISBN 978-0-387-75968-5. [Google Scholar]
- Simpson, G.L. Permute: Functions for Generating Restricted Permutations of Data (Version 0.7-0). Available online: http://CRAN.R-project.org/package=permute (accessed on 1 December 2013).
- R Core Team. Foreign: Read Data Stored by Minitab, S, SAS, SPSS, Stata, Systat, dBase, … (Version 0.8-55). Available online: http://CRAN.R-project.org/package=foreign (accessed on 1 December 2013).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling (Version 2.1-49). Available online: http://CRAN.R-project.org/package=raster (accessed on 1 December 2013).
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the Geospatial Data Abstraction Library (Version 0.8-10). Available online: http://CRAN.R-project.org/package=rgdal (accessed on 1 December 2013).
- Pebesma, E.J.; Bivand, R.S. Classes and methods for spatial data in R. R News 2005, 5, 9–13. [Google Scholar]
- Bivand, R.S.; Pebesma, E.J.; Gomez-Rubio, V. Applied Spatial Data Analysis with R; Springer: New York, NY, USA, 2008. [Google Scholar]
- Nicolau, R. (Maria Rita Rodrigues Carmona Nicolau) Modelação e Mapeamento da Distribuição Espacial de Precipitação-Uma Aplicação a Portugal Continental. Ph.D. Thesis, Universidade Nova de Lisboa-Faculdade de Ciências e Tecnologia, Lisboa, Portugal, 2002. [Google Scholar]
- Gonzalo Jiménez, J. Diagnosis Fitoclimática de La España Peninsular: Hacia Un Modelo de Clasificación Funcional de La Vegetación Y de Los Ecosistemas Peninsulares Españoles, 1st ed.; Serie Técnica Naturaleza y Parques Nacionales; Organismo Autónomo de Parques nacionales, Ministerio de Medio Amiente y Medio Rural y Marino de España: Madrid, Spain, 2010; ISBN 978-84-8014-787-3. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Juan, L.P.; Jones, P.G.; Jarvis, A.; Richardson, K. Worldclim. Available online: http://www.worldclim.org/ (accessed on 31 May 2009).
- Serviços Geológicos de Portugal. Carta Geológica de Portugal na Escala 1:500 000; SGP: Lisboa, Portugal, 1992. [Google Scholar]
- Naimi, B. Usdm: Uncertainty Analysis for Species Distribution Models (Version 1.1-12). Available online: https://rdrr.io/cran/usdm/ (accessed on 25 February 2016).
- Chessel, D.; Dufour, A.B.; Thioulouse, J. The ade4 package—I: One-table methods. R News 2004, 4, 5–10. [Google Scholar]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression models for count data in R. J. Stat. Softw. 2008, 27. [Google Scholar] [CrossRef]
- Mullahy, J. Specification and testing of some modified count data models. J. Econ. 1986, 33, 341–365. [Google Scholar] [CrossRef]
- Lambert, D. Zero-Inflated Poisson Regression, with an Application to Defects in Manufacturing. Technometrics 1992, 34, 1–14. [Google Scholar] [CrossRef]
- R Core Team. R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria (Version 3.2.0). Available online: http://www.R-project.org/ (accessed on 25 December 2015).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package (Version 2.2-1). Available online: http://CRAN.R-project.org/package=vegan (accessed on 25 February 2016).
- Jackman, S. Pscl: Classes and Methods for R Developed in the Political Science Computational Laboratory (Version 1.4.9); Department of Political Science, Stanford University: Stanford, CA, USA, 2015. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Burnham, K.P. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95364-9. [Google Scholar]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations and comparisons. Behav. Ecol. Sociobiol. 2010, 65, 23–35. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas Sáenz, S.; Penas, Á. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar] [CrossRef]
- Woodward, F.I.; Williams, B.G. Climate and plant distribution at global and local scales. Vegetatio 1987, 69, 189–197. [Google Scholar] [CrossRef]
- Franco, J. do Quercus L. In Flora iberica: Plantas Vasculares de la Península Ibérica e Islas Baleares; Castroviejo, S., Laínz, M., López González, G., Montserrat, P., Muñoz Garmendia, F., Paiva, J., Villar, L., Eds.; Real Jardín Botánico-Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1990; Volume II, pp. 15–36. ISBN 84-00-07034-8. [Google Scholar]
- Gómez-Aparicio, L.; Zavala, M.A.; Bonet, F.J.; Zamora, R. Are pine plantations valid tools for restoring mediterranean forests? An assessment along abiotic and biotic gradients. Ecol. Appl. 2009, 19, 2124–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hees, A.F.M. Growth and morphology of pedunculate oak (Quercus robur L) and beech (Fagus sylvatica L) seedlings in relation to shading and drought. Ann. Sci. For. 1997, 54, 9–18. [Google Scholar] [CrossRef]
- Welander, N.T.; Ottosson, B. The influence of shading on growth and morphology in seedlings of Quercus robur L. and Fagus sylvatica L. For. Ecol. Manag. 1998, 107, 117–126. [Google Scholar] [CrossRef]
- Ziegenhagen, B.; Kausch, W. Productivity of young shaded oaks (Quercus robur L.) as corresponding to shoot morphology and leaf anatomy. For. Ecol. Manag. 1995, 72, 97–108. [Google Scholar] [CrossRef]
- Valladares, F.; Chico, J.M.; Aranda, I.; Balaguer, L.; Dizengremel, P.; Manrique, E.; Dreyer, E. The greater seedling high-light tolerance of Quercus robur over Fagus sylvatica is linked to a greater physiological plasticity. Trees Struct. Funct. 2002, 16, 395–403. [Google Scholar] [CrossRef]
- Pausas, J.G.; Blade, C.; Valdecantos, A.; Seva, J.P.; Fuentes, D.; Alloza, J.A.; Vilagrosa, A.; Bautista, S.; Cortina, J.; Vallejo, R. Pines and oaks in the restoration of Mediterranean landscapes of Spain: New perspectives for an old practice—A review. Plant Ecol. 2004, 171, 209–220. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Loureiro, C.; Magalhães, M.; Ferreira, P.; Fernandes, M. Fuel age, weather and burn probability in Portugal. Int. J. Wildland Fire 2012, 21, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, M.; Gomes, L.; Tarelho, L.; Pio, C. Formaldehyde and acetaldehyde emissions from residential wood combustion in Portugal. Atmos. Environ. 2013, 72, 171–176. [Google Scholar] [CrossRef]
- Gonçalves, C.; Alves, C.; Pio, C. Inventory of fine particulate organic compound emissions from residential wood combustion in Portugal. Atmos. Environ. 2012, 50, 297–306. [Google Scholar] [CrossRef]
- Tiscar-Oliver, P. Patterns of shrub diversity and tree regeneration across topographic and stand-structural gradients in a Mediterranean forest. For. Syst. 2015, 24, 11. [Google Scholar] [CrossRef] [Green Version]
- Bugalho, M.N.; Caldeira, M.C.; Pereira, J.S.; Aronson, J.; Pausas, J.G. Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front. Ecol. Environ. 2011, 9, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Simões, M.P.; Belo, A.F.; Fernandes, M.; Madeira, M. Regeneration patterns of Quercus suber according to montado management systems. Agrofor. Syst. 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Capelo, J. Nemorum Transtaganae Descriptio: Sintaxonomia Numérica das Comunidades Florestais e Pré-florestais do Baixo Alentejo. Ph.D. Thesis, Universidade Técnica de Lisboa-Instituto Superior de Agronomia, Lisboa, Portugal, 2007. [Google Scholar]
- Plieninger, T. Compatibility of livestock grazing with stand regeneration in Mediterranean holm oak parklands. J. Nat. Conserv. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Pulido, F.; Garcia, E.; Obrador, J.J.; Moreno, G. Multiple pathways for tree regeneration in anthropogenic savannas: Incorporating biotic and abiotic drivers into management schemes. J. Appl. Ecol. 2010, 47, 1272–1281. [Google Scholar] [CrossRef]
- Pulido, F.J.; Diaz, M.; de Trucios, S.J.H. Size structure and regeneration of Spanish holm oak Quercus ilex forests and dehesas: Effects of agroforestry use on their long-term sustainability. For. Ecol. Manag. 2001, 146, 1–13. [Google Scholar] [CrossRef]
- Maia, P.; Keizer, J.; Vasques, A.; Abrantes, N.; Roxo, L.; Fernandes, P.; Ferreira, A.; Moreira, F. Post-fire plant diversity and abundance in pine and eucalypt stands in Portugal: Effects of biogeography, topography, forest type and post-fire management. For. Ecol. Manag. 2014, 334, 154–162. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Sáenz Laín, C. Enumeración de los quercus de la península ibérica. Rivasgodaya 1991, 6, 101–110. [Google Scholar]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees Struct. Funct. 2004, 18, 83–92. [Google Scholar] [CrossRef]
- Corcuera, L.; Notivol, E. Differences in photosynthetic activity might explain the large-scale shifts in pine recruitment in favour of oaks in continental Mediterranean climates. Forestry 2015, 88, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Bugalho, M.N.; Milne, J.A.; Racey, P.A. The foraging ecology of red deer (Cervus elaphus) in a Mediterranean environment: Is a larger body size advantageous? J. Zool. 2001, 255, 285–289. [Google Scholar] [CrossRef]
- Bugalho, M.N.; Milne, J.A. The composition of the diet of red deer (Cervus elaphus) in a Mediterranean environment: A case of summer nutritional constraint? For. Ecol. Manag. 2003, 181, 23–29. [Google Scholar] [CrossRef]
- Pasquini, S.; Mizzau, M.; Petrussa, E.; Braidot, E.; Patui, S.; Gorian, F.; Lambardi, M.; Vianello, A. Seed storage in polyethylene bags of a recalcitrant species (Quercus ilex): Analysis of some bio-energetic and oxidative parameters. Acta Physiol. Plant. 2012, 34, 1963–1974. [Google Scholar] [CrossRef]
- Catry, F.X.; Pausas, J.G.; Moreira, F.; Fernandes, P.M.; Rego, F. Post-fire response variability in Mediterranean Basin tree species in Portugal. Int. J. Wildland Fire 2013, 22, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Enright, N.J.; Knox, K.J.E. Resprouting as a key functional trait: How buds, protection and resources drive persistence after fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Vayreda, J.; Gracia, M.; Martinez-Vilalta, J.; Retana, J. Patterns and drivers of regeneration of tree species in forests of peninsular Spain. J. Biogeogr. 2013, 40, 1252–1265. [Google Scholar] [CrossRef]
- Murdoch, D.; Chow, E.D. Ellipse: Functions for Drawing Ellipses and Ellipse-Like Confidence Regions (Version 0.3-8). Available online: http://CRAN.R-project.org/package=ellipse (accessed on 31 May 2013).
- Le Bas, M.; Streckeisen, L. The IUGS systematics of igneous rocks. J. Geol. Soc. Lond. 1991, 148, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Le Maitre, R.W.; Streckeisen, A.; Zanettin, B.; Le Bas, M.J.; Bonin, B.; Bateman, P.; Bellieni, G.; Dudeck, A.; Efremova, S.; Keller, J.; et al. Igneous Rocks: A Classification and Glossary of Terms: Recommendations of the International Union of Geological Sciences, Subcommission on the Systematics of Igneous Rocks, 2nd ed.; Cambridge University Press: Cambridge, NY, USA, 2004; ISBN 978-0-521-61948-6. [Google Scholar]
- Direção-Geral do Território Carta Administrativa Oficial de Portugal-Versão 2011. Available online: http://www.dgterritorio.pt/cartografia_e_geodesia/cartografia/carta_administrativa_oficial_de_portugal__caop_/caop__download_/carta_administrativa_oficial_de_portugal___versao_2011/ (accessed on 26 September 2011).
| Abbreviation | Variable Name (Units or Levels) | Variable Type | Related Physical Phenomenon |
|---|---|---|---|
| G | Basal area (m2 ha−1). | Forest structure | Light/space availability (proxy). |
| N | Tree density, independently of the species (no. of trees per m2). | Forest structure | Light/space availability (proxy). |
| GRAZI | Evidence of grazing by domestic animals, that is, cattle, equine, sheep, goat or swine (“yes”, “no”). | Anthropic (animal husbandry) | Vegetation disturbance. |
| UnderClear | Evidence of bushes removal, that is, mechanical removal, prescribed fire, manual removal, etc. (“yes”, “no”). | Anthropic (management) | Vegetation disturbance/space availability. |
| SeedAvail | Density of plots with conspecific adult trees (no. of plots in a predefined neighbourhood). | Seed source | Local to regional seed availability/conspecific adult presence. |
| DensPp | Density of plots with adult P. pinaster trees (no. of plots in a predefined neighbourhood). | Anthropic (land-use) | Local to regional forestry intensity (P. pinaster). |
| DensEg | Density of plots with adult E. globulus trees (no. of plots in a predefined neighbourhood). | Anthropic (land-use) | Local to regional forestry intensity (E. globulus). |
| FireFreq | Fire frequency (no. of fires between 1975–2006). | Fire | Recurrent vegetation disturbance. |
| TSLFire | Time since last fire (“≤5 years”, “>5 years”). | Fire | Vegetation disturbance/space availability. |
| OMBRO | Summer quarter ombrothermic index (mm °C−1). | Climatic | Summer climatic moisture (proxy). |
| THERM | Compensated thermicity index (°C*10). | Climatic | Air temperature. |
| TRASP | Transformed aspect (0 to 1). | Physiographic | Relative solar radiation; also related to moisture, temperature and wind shelter (proxy). |
| SLOPE | Slope (drop/distance). | Physiographic | Soil deepness (proxy); accessibility (proxy). |
| SoilMoist | Logarithm of the contributing area weighted by OMBRO or AOI (mm °C−1). | Hydrologic/edaphic | Summer soil moisture/annual soil moisture (proxy). |
| SPMType | Soil parent material types (“alluvia”, “siliceous light”, “siliceous loamy”, “siliceous heavy”, “intermediate loamy”, “basic heavy”, “ultramafic heavy”). | Edaphic | Parent material base richness and potential soil texture. |
| Taxa | No. of Removed Extreme Observations | SPMType Tested Classes 1 | Total No. of Plots Used in Modelling |
|---|---|---|---|
| Q. robur | 0 | 1, 2 | 4167 |
| Q. pyrenaica | 2 | 1, 2, 4 | 4461 |
| Q. broteroi | 1 | 1, 2, 3, 4 | 4631 |
| Q. suber | 3 | 1, 2, 4 | 4485 |
| Q. rotundifolia | 1 | 1, 2, 3, 4 | 4642 |
| Taxa | % Plots without Saplings | Excluding Plots without Saplings | Including Plots without Saplings | ||
|---|---|---|---|---|---|
| Saplings Mean | Saplings Variance | Saplings Mean | Saplings Variance | ||
| Q. robur | 96.0% | 4.7 | 35.5 | 0.2 | 2.3 |
| Q. pyrenaica | 96.8% | 7.8 | 113.3 | 0.3 | 5.5 |
| Q. broteroi | 99.3% | 2.0 | 2.0 | 0.01 | 0.04 |
| Q. suber | 94.6% | 3.1 | 15.5 | 0.2 | 1.3 |
| Q. rotundifolia | 95.2% | 8.2 | 128.8 | 0.4 | 9.3 |
| Taxa | SeedAvail (count) | DensPp (count) | DensEg (count) | SoilMoist (count) | SeedAvail (zero) | SoilMoist (zero) |
|---|---|---|---|---|---|---|
| Q. robur | 2 km | 10 km | - | - | 10 km | summer |
| Q. pyrenaica | 2 km | 2 km | - | - | 5 km | - |
| Q. broteroi | 5 km | - | 10 km | annual | 10 km | annual |
| Q. suber | - | - | 10 km | - | 2 km | - |
| Q. rotundifolia | 10 km | - | - | - | 2 km | - |
| Taxa | No. of Models |
|---|---|
| Q. robur | 5289 |
| Q. pyrenaica | 1275 |
| Q. broteroi | 16,249 |
| Q. suber | 4272 |
| Q. rotundifolia | 18,551 |
| Variable | Variable Importance Values (Based on Model Akaike Weights) | |||||
|---|---|---|---|---|---|---|
| Q. robur | Q. pyrenaica | Q. broteroi | Q. suber | Q. rotundifolia | ||
| Count component | G | 95.8% (↑) | 72.1% (↓) | 26.7% (↑↓) | 99.9% (↑) | 28.3% (↑↓°) |
| N | 86.0% (↓) | 63.1% (↓) | 66.6% (↑↓°) | 45.8% (↓) | 33.2% (↑↓°) | |
| GRAZI | 45.2% (↓) | 61.6% (↓) | 40.2% (↓) | 98.9% (↓) | 30.1% (↑°↓) | |
| UnderClear | 56.6% (↓) | 99.4% (↓) | 35.1% (↓) | 29.8% (↓) | 31.5% (↑) | |
| SeedAvail | 100.0% (↑) | 99.9% (↑) | 99.9% (↑) | 34.9% (↑) | 98.8% (↑) | |
| DensPp | 76.4% (↑) | 78.0% (↓) | 31.2% (↑↓) | 30.7% (↑) | 32.8% (↑↓°) | |
| DensEg | 29.3% (↑↓) | 41.5% (↓) | 81.8% (↓) | 92.5% (↑) | 28.1% (↑↓) | |
| FireFreq | 100.0% (↓) | 93.6% (↓) | 29.2% (↑↓) | 31.9% (↑) | 60.8% (↑°↓) | |
| TSLFire | 34.0% (↑) | 27.7% (↑↓) | 47.4% (↑°↓) | 27.9% (↑↓°) | 73.0% (↓) | |
| OMBRO | 93.8% (↓) | 99.5% (↓) | 75.8% (↑°↓) | 99.0% (↓) | 54.5% (↑↓°) | |
| THERM | 36.5% (↑°↓) | 29.9% (↑↓) | 73.9% (↑) | 29.0% (↑↓) | 62.0% (↑°↓) | |
| TRASP | 83.7% (↓) | 94.2% (↓) | 68.5% (↑↓°) | 40.9% (↓) | 63.6% (↓) | |
| SLOPE | 27.5% (↑°↓) | 33.1% (↓) | 35.6% (↑) | 100.0% (↑) | 100.0% (↑) | |
| SoilMoist | 48.6% (↑) | 28.6% (↑↓) | 69.4% (↑°↓) | 30.6% (↓) | 41.5% (↑) | |
| SPMType | 28.8% | 99.8% | 29.3% | 15.2% | 71.9% | |
| Zero inflation component | SeedAvail | 100.0% (↓) | 100.0% (↓) | 100.0% (↓) | 100.0% (↓) | 100.0% (↓) |
| OMBRO | 100.0% (↓) | 100.0% (↓) | 100.0% (↑°↓) | 99.8% (↓) | 96.7% (↑) | |
| THERM | 92.1% (↓) | 100.0% (↑) | 76.5% (↑°↓) | 39.1% (↓) | 67.2% (↑↓°) | |
| SoilMoist | 75.5% (↓) | 48.7% (↑) | 69.1% (↑°↓) | 30.4% (↓) | 57.4% (↓) | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro-Henriques, T.; Fernandes, P.M. Regeneration of Native Forest Species in Mainland Portugal: Identifying Main Drivers. Forests 2018, 9, 694. https://doi.org/10.3390/f9110694
Monteiro-Henriques T, Fernandes PM. Regeneration of Native Forest Species in Mainland Portugal: Identifying Main Drivers. Forests. 2018; 9(11):694. https://doi.org/10.3390/f9110694
Chicago/Turabian StyleMonteiro-Henriques, Tiago, and Paulo M. Fernandes. 2018. "Regeneration of Native Forest Species in Mainland Portugal: Identifying Main Drivers" Forests 9, no. 11: 694. https://doi.org/10.3390/f9110694
APA StyleMonteiro-Henriques, T., & Fernandes, P. M. (2018). Regeneration of Native Forest Species in Mainland Portugal: Identifying Main Drivers. Forests, 9(11), 694. https://doi.org/10.3390/f9110694