Allometry, Growth and Survival of Three Eucalyptus Species (Eucalyptus benthamii Maiden and Cambage, E. dunnii Maiden and E. grandis Hill ex Maiden) in High-Density Plantations in Uruguay

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Sampling and Measurements
2.4. Volume Equation
2.5. Optimum Time of Harvest
2.6. Statistical Analysis
3. Results
3.1. Survival
3.2. Diameter at Breast Height, Total Height and Slenderness
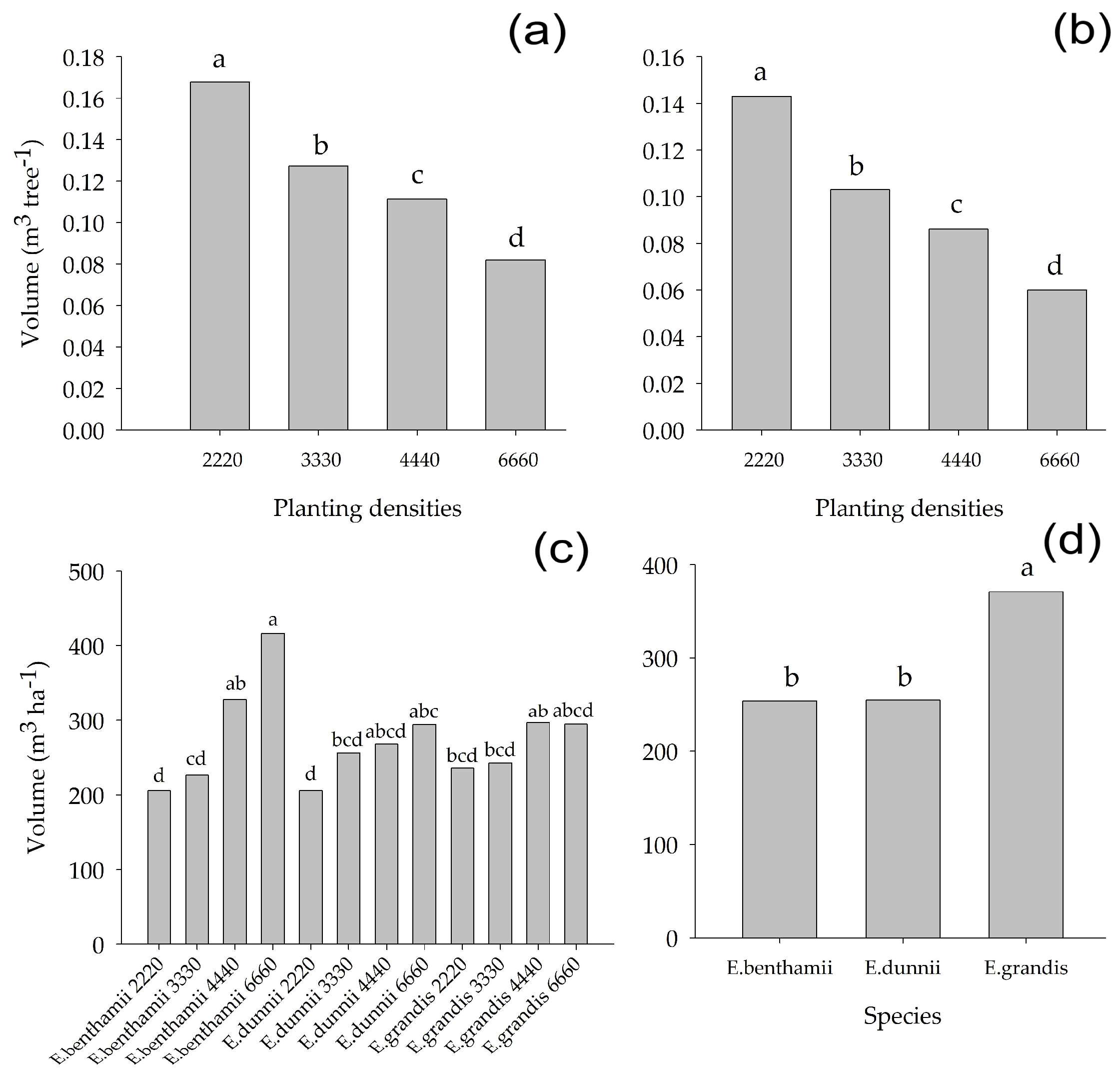
3.3. Alometric Equations and Volume
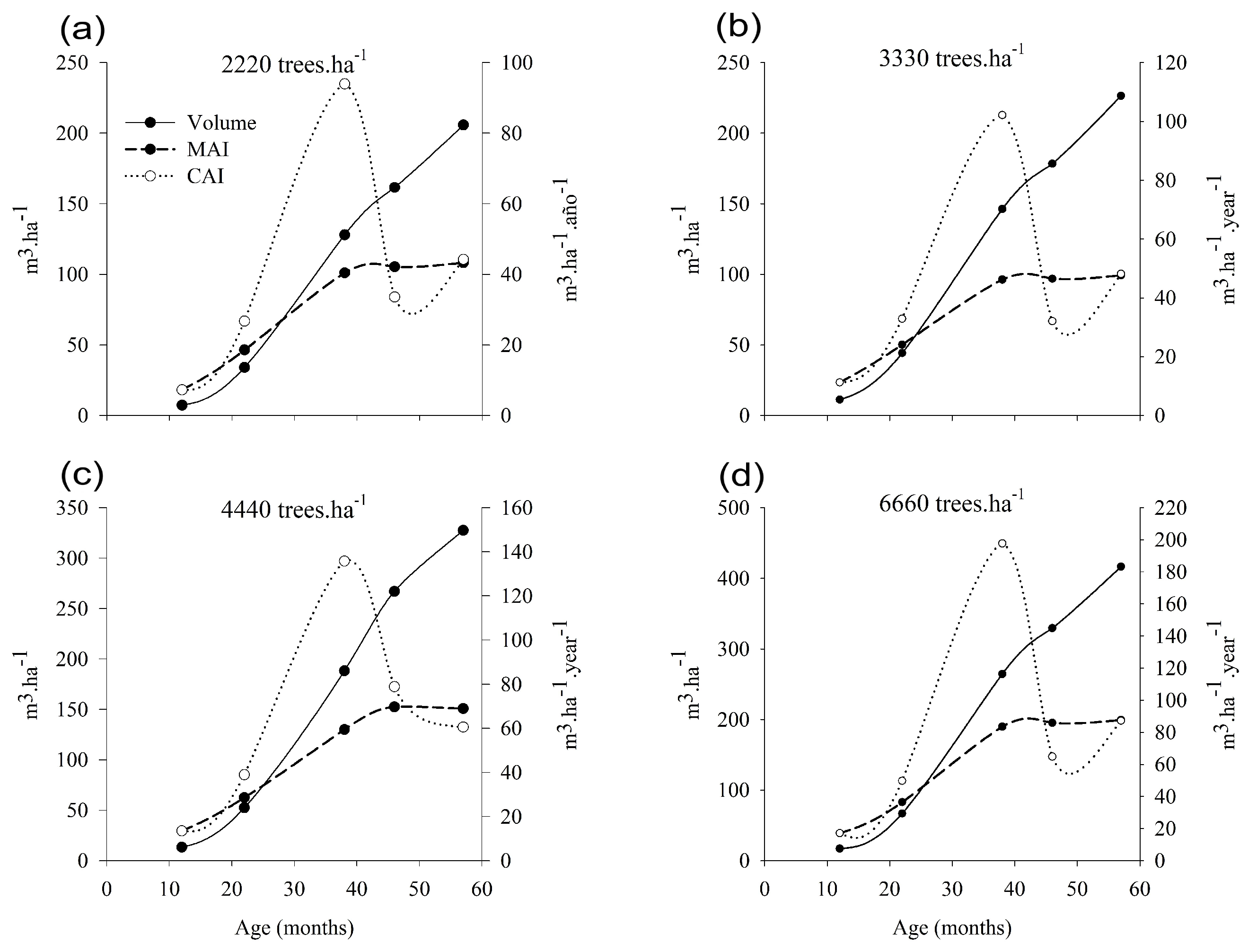
3.4. Optimum Time of Harvest
4. Discussion
4.1. Survival
4.2. Height, Diameter and Slenderness
4.3. Alometric Equations and Volume
4.4. Optimum Time of Harvest
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sixto, H.; Hernández, M.J.; Barrio, M.; Cañellas, J.C.I. Plantaciones del género Populus para la producción de biomasa con fines energéticos: Revisión. Investig. Agrar. Sist. Recur. For. 2007, 16, 277–294. [Google Scholar] [CrossRef]
- Estrada, C.A.; Meneses, A.Z. Gasificación de biomasa para producción de combustibles de bajo poder calorífico y su utilización en generación de potencia y calor. Scientia et Technica. 2004, 2. [Google Scholar] [CrossRef]
- Gelfand, I.; Sahajpal, R.; Zhang, X.; Izaurralde, R.C.; Gross, K.L.; Robertson, G.P. Sustainable bioenergy production from marginal lands in the US Midwest. Nature 2013, 493, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef]
- Bravo, F.; Bravo-Oviedo, A.; Diaz-Balteiro, L. Carbon sequestration in Spanish Mediterranean forests under two management alternatives: A modeling approach. Eur. J. For. Res. 2008, 127, 225–234. [Google Scholar] [CrossRef]
- González García, M.; Hevia Cabal, M.; Barrio Anta, M. Modelo dinámico de crecimiento y producción de biomasa para cultivos energéticos de Eucalyptus nitens (Maiden) en el noroeste de España. In 6 to Congreso Forestal Español; Sociedad Española de Ciencias Forestales: Vitoria Gateiz, Spain, 2013; pp. 2–8. Available online: http://secforestales.org/publicaciones/index.php/congresos_forestales/article/viewFile/14333/14176 (accessed on 15 June 2017).
- Razakamanarivo, R.H.; Razakavololona, A.; Razafindrakoto, M.-A.; Vieilledent, G.; Albrecht, A. Below-ground biomass production and allometric relationships of eucalyptus coppice plantation in the central highlands of Madagascar. Biomass Bioenergy 2012, 45, 1–10. [Google Scholar] [CrossRef]
- Winck, R.A.; Fassola, H.E.; Barth, S.R.; Crechi, E.; Keller, A.E.; Videla, D.; Zaderenko, C. Modelos predictivos de biomasa aérea de Eucalyptus grandis para el noreste de Argentina. Ciência Florestal 2015, 25, 595–606. Available online: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1980-50982015000300595&nrm=iso (accessed on 1 August 2017). [CrossRef]
- Ounban, W.; Puangchit, L.; Diloksumpun, S. Development of general biomass allometric equations for Tectona grandis Linn. f. and Eucalyptus camaldulensis Dehnh. plantations in Thailand. Agric. Nat. Resour. 2016, 50, 48–53. [Google Scholar] [CrossRef]
- Methol, R. Comparisons of Approaches to Modelling Tree Taper, Stand Structure and Stand Dynamics in Forest Plantations. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2001. [Google Scholar]
- Hirigoyen, A.; Rachid, C.; Varela, F. Actualizaciones de herramientas informáticas para la toma de decisiones forestales. Revista INIA 2013, 35, 58–62. [Google Scholar]
- Rachid, C. SAG Eucalyptus: Un nuevo sistema de apoyo a la gestión de plantaciones de Eucalyptus orientadas a la producción de celulosa. Revista INIA 2008, 15, 35–37. [Google Scholar]
- Rachid-Casnati, C.; Mason, E.G.; Woollons, R.; Resquin, F. Volume and taper equations for P. taeda (L.) and E. grandis (Hill ex. Maiden). Agrociencia (Montevideo) 2014, 18, 47–60. [Google Scholar]
- Castaño, J.P.G.; Ceroni, A.; Furest, M.; Aunchayna, J.; Bidegain, R. Caracterización agroclimática del Uruguay 1980–2009. Ser. Técnica INIA 2011, 193, 33. [Google Scholar]
- Bentancor, L. Extracción de Nutrientes por Eucalyptus dunnii Maiden de 4 años con Destino a la Producción de Biomasa para Energía y Celulosa. Master’s Thesis, Universidad de la República, Montevideo, Uruguay, December 2017; p. 99. [Google Scholar]
- Rodríguez, A.; Cancino, J.; Acuña, E.; Rubilar, R.; Muñoz, E. Evaluación del crecimiento de plantaciones dendroenergéticas de Eucalyptus globulus, según densidad de plantación y turno de rotacion en suelos contrastantes de la región del Biobío, Chile. Cienc. Investig. For. INFOR Chile 2013, 19, 18. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2012. [Google Scholar]
- Harris, F. The Effect of Competition on Stand, Tree, and Wood Growth and Structure in Subtropical Eucalyptus Grandis Plantations. Ph.D. Thesis, Southern Cross University, Lismore, Australia, January 2007. [Google Scholar]
- Schneider, P.R.; Finger, C.A.G.; Schneider, P.S.P.; Fleig, F.D.; da Cunha, T.A. Influência do espaçamento no autodesbaste de povoamento monoclonal de Eucalyptus Saligna Smith. Ciênc. Florest. 2015, 25, 119–126. [Google Scholar] [CrossRef]
- Larson, A.J.; Lutz, J.A.; Donato, D.C.; Freund, J.A.; Swanson, M.E.; HilleRisLambers, J.; Sprugel, D.G.; Franklin, J.F. Spatial aspects of tree mortality strongly differ between young and old-growth forests. Ecology 2015, 96, 2855–2861. [Google Scholar] [CrossRef] [PubMed]
- Van Gunst, K.J.; Weisberg, P.J.; Yang, J.; Fan, Y. Do denser forests have greater risk of tree mortality: A remote sensing analysis of density-dependent forest mortality. For. Ecol. Manag. 2016, 359, 19–32. [Google Scholar] [CrossRef]
- Schönau, A.P.G.; Coetzee, J. Initial spacing, stand density and thinning in eucalypt plantations. For. Ecol. Manag. 1989, 29, 245–266. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Fensham, R.J.; Fairfax, R.J.; Buckley, Y.M. Neighbourhood effects influence drought-induced mortality of savanna trees in Australia. J. Veg. Sci. 2010, 21, 573–585. [Google Scholar] [CrossRef]
- Lonsdale, W.M. The Self-Thinning Rule: Dead or Alive? Ecology 1990, 71, 1373–1388. [Google Scholar] [CrossRef]
- Lintz, H.E.; Gray, A.N.; Yost, A.; Sniezko, R.; Woodall, C.; Reilly, M.; Hutten, K.; Elliott, M. Quantifying density-independentmortality of temperate tree species. Ecol. Indic. 2016, 66, 1–9. [Google Scholar] [CrossRef]
- Puettman, K.; Hibbs, D.; Hann, D. The Dynamics of Mixed Stands of Alnus Rubra and Pseudotsuga Menziesii: Extension of Size—Density Analysis to Species Mixture. J. Ecol. 2016, 80, 449–458. [Google Scholar] [CrossRef]
- Gendreau-Berthiaume, B.; Macdonald, S.E.; Stadt, J.J. Extended density-dependent mortality in mature conifer forests: Causes and implications for ecosystem management. Ecol. Appl. 2016, 26, 1486–1502. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, J.P.R.; Ponce, E.V. Growth and economic analysis of a Eucalyptus globulus clonal spacing trial in Chile. In Proceedings of the Australasian Forest Genetics Conference, Breeding for Wood Quality, Hobart, Australia, 11–14 April 2007. [Google Scholar]
- Akhtar, J.; Saqib, Z.A.; Qureshi, R.H.; Haq, M.A.; Iqbal, M.S.; Marcar, N.E. The effect of spacing on the growth of Eucalyptus camaldulensis on salt-affected soils of the Punjab, Pakistan. Can. J. For. Res. 2008, 38, 2434–2444. [Google Scholar] [CrossRef]
- Sale, G. A Comparison of Between-Tree Competition Effects in Separate Stands of a Eucalyptus Grandis Seedling and a Single Eucalyptus Grandis x Urophylla Hybrid. Master’s Thesis, University of the Witwatersrand, Johannesburg, South Africa, November 2005. [Google Scholar]
- Jacobs, M.R. Growth Habits of the Eucalypts; Forestry and Timber Bureau: Canberra, Australia, 1955; Available online: http://trove.nla.gov.au/version/28637821 (accessed on 25 April 2017).
- INUMET. Available online: https://www.inumet.gub.uy/clima/estadisticas-climatologicas (accessed on 4 January 2017).
- Bouvet, J.M. Effect of spacing on juvenile growth and variability of eucalyptus clones. Can. J. For. Res. 1997, 27, 174–179. [Google Scholar] [CrossRef]
- Newton, P.F.; Jolliffe, P.A. Assessing processes of intraspecific competition within spatially heterogeneous black spruce stands. Can. J. For. Res. 1998, 28, 259–275. [Google Scholar] [CrossRef]
- Brand, D.G.; Magnussen, S. Asymmetric, two-sided competition in even-aged monocultures of red pine. Can. J. For. Res. 1988, 18, 901–910. [Google Scholar] [CrossRef]
- Von Euler, F.; Baradat, P.; Lemoine, B. Effects of plantation density and spacing on competitive interactions among half-sib families of maritime pine. Can. J. For. Res. 1992, 22, 482–489. [Google Scholar] [CrossRef]
- Tomé, M.; Tomé, J.A.; Araújo, M.C.; Pereira, J.S. Intraspecific competition in irrigated and fertilized eucalypt plantations. For. Ecol. Manag. 1994, 69, 211–218. [Google Scholar] [CrossRef]
- De Moraes Goncalves, J.L.; Stape, J.L.; Laclau, J.-P.; Smethurst, P.; Gava, J.L. Silvicultural effects on the productivity and wood quality of eucalypt plantations. For. Ecol. Manag. 2004, 193, 45–61. [Google Scholar] [CrossRef]
- De Córdova Machado, F.; Guerra, S.P.S.; Ceragioli, N.; Oguri, G.; Denadai, M.S. Influência do espaçamento na produtividade e alocação de biomassa em um plantio de eucalyptus grandis. In Proceedings of the Congresso Internacional de Bioenergia, São Paulo, Brazil, January 2012; pp. 1–6. [Google Scholar]
- De Jesus, H.; Junior, E.; Ballarin, A.W. Influência Do Espaçamento Na Densidade Básica Da Madeira Em Sistemas Florestais De Curta Rotação. In Proceedings of the 8th Congresso Internacional de Bioenergia, São Paulo, Brazil, 5–7 November 2013; pp. 5–9. [Google Scholar]
- Alcorn, P.J.; Pyttel, P.; Bauhus, J.; Smith, R.G.B.; Thomas, D.; James, R.; Nicotra, A. Effects of initial planting density on branch development in 4-year-old plantation grown Eucalyptus pilularis and Eucalyptus cloeziana trees. For. Ecol. Manag. 2007, 252, 41–51. [Google Scholar] [CrossRef]
- De Oliveira Neto, S.N.; dos Reis, G.G.; Reis, M.D.G.F.; Leite, H.G.; Neves, J.C.L. Crescimento e Distribuição Diamétrica de Eucalyptus camaldulensis em Diferentes Espaçamentos e Níveis de Adubação na Região de Cerrado de Minas Gerais. Floresta 2010, 40, 755–762. [Google Scholar] [CrossRef]
- Patiño-valera, F.; Kageyama, P.Y. Interaction genotype X spacing in progenies of Eucalyptus saligna Smith. IPEF Int. 1990, 1, 12–22. [Google Scholar]
- Assis, R.; Ferreira, M.; de Morais, E.; Fernandes, L. Produção de biomassa de Eucalyptus urophylla s.t. blake sob diferentes espaçamentos na região de cerrado de Minas Gerais. Rev. Arvore 1999, 23, 151–156. [Google Scholar]
- Forrester, D.I.; Wiedemann, J.C.; Forrester, R.I.; Baker, T.G. Effects of planting density and site quality on mean tree size and total stand growth of E.ucalyptus globulus plantations. Can. J. For. Res. 2013, 43, 846–851. [Google Scholar] [CrossRef]
- Christina, M.; Laclau, J.-P.; Gonçalves, J.L.M.; Jourdan, C.; Nouvellon, Y.; Bouillet, J.-P. Almost symmetrical vertical growth rates above and below ground in one of the world’s most productive forests. Ecosphere 2011, 2, 1–10. [Google Scholar] [CrossRef]
- Trouvé, R.; Bontemps, J.D.; Collet, C.; Seynave, I.; Lebourgeois, F. Growth partitioning in forest stands is affected by stand density and summer drought in sessile oak and Douglas-fir. For. Ecol. Manag. 2014, 334, 358–368. [Google Scholar] [CrossRef]
- Campoe, O.C.; Stape, J.L.; Nouvellon, Y.; Laclau, J.P.; Bauerle, W.L.; Binkley, D.; Le Maire, G. Stem production, light absorption and light use efficiency between dominant and non-dominant trees of Eucalyptus grandis across a productivity gradient in Brazil. For. Ecol. Manag. 2013, 288, 14–20. [Google Scholar] [CrossRef]
- Binkley, D.; Campoe, O.C.; Gspaltl, M.; Forrester, D.I. Light absorption and use efficiency in forests: Why patterns differ for trees and stands. For. Ecol. Manag. 2013, 288, 5–13. [Google Scholar] [CrossRef]
- Xue, L.; Pan, L.; Zhang, R.; Xu, P.B. Density effects on the growth of self-thinning Eucalyptus urophylla stands. Trees 2011, 25, 1021–1031. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Bauerle, W.L.; Ryan, M.G. Explaining growth of individual trees: Light interception and efficiency of light use by Eucalyptus at four sites in Brazil. For. Ecol. Manag. 2010, 259, 1704–1713. [Google Scholar] [CrossRef]
- Rance, S.J.; Mendham, D.S.; Cameron, D.M.; Grove, T.S. An evaluation of the conical approximation as a generic model for estimating stem volume, biomass and nutrient content in young Eucalyptus plantations. New For. 2012, 43, 109–128. [Google Scholar] [CrossRef]
- Kuyah, S.; Dietz, J.; Muthuri, C.; van Noordwijk, M.; Neufeldt, H. Allometry and partitioning of above- and below-ground biomass in farmed eucalyptus species dominant in Western Kenyan agricultural landscapes. Biomass Bioenergy 2013, 55, 276–284. [Google Scholar] [CrossRef]
- Vega-nieva, D.J. Modeling the above and belowground biomass of planted and coppiced Eucalytpus globulus stands in NW Spain. Ann. For. Sci. 2015, 72, 967–980. [Google Scholar] [CrossRef]
- Segura Madrigal, M.A.; Andrade Castañeda, H.J. Cómo construir modelos alométricos de volume. biomasa o carbono de especies leñosas perennes. J. Agroforestería en las Américas 2008, 46, 89–96. [Google Scholar]
- Berger, R.; Schneider, P.R.; Finger, C.A.G.; Haselein, C.R. Efeito do espaçamento e da aduabação no crescimento de um clone de Eucalyptus saligna Smith. Ciênc. Florest. 2002, 12, 75–87. [Google Scholar] [CrossRef]
- Pinkard, E.A.; Neilsen, W.A. Crown and stand characteristics of Eucalyptus nitens in response to initial spacing: Implications for thinning. For. Ecol. Manag. 2003, 172, 215–227. [Google Scholar] [CrossRef]
- Morais, V.de.M. Eucalipto Clonal Sob Diferentes Espaçamentos, Na Região Noroeste Do Estado De Minas Gerais. Available online: http://repositorio.ufla.br/bitstream/1/3766/1/DISSERTA%c3%87%c3%83O_Din%c3%a2mica%20de%20crescimento%20de%20eucalipto%20clonal%20sob%20diferentes%20espa%c3%a7amentos%2c%20na%20regi%c3%a3o%20noroeste%20do%20estado%20de%20Minas%20Gerais.pdf (accessed on 13 November 2018).
- Leite, F.P.; de Barros, N.F.; de Novais, R.F.; Sans, L.M.A.; Fabres, A.S. Crescimento de Eucalyptus grandis em diferentes densidades populacionais. Rev. Árvore 1997, 21, 313–321. [Google Scholar]
- Balloni, E.A.; Simoes, J.W. O espaçamento de plantio e suas implicações silviculturais. Piracicaba IPEF 1980, 1, 1–16. [Google Scholar]
- Goulart, M.; Haselein, C.R.; Hoppe, J.M.; Farias, J.A.; Pauleski, D.T. Massa específica básica e massa seca de madeira de Eucalyptus grandis sob o efeito do espaçamento de plantio e da posição axial no tronco. Ciênc. Florest. 2003, 13, 167–175. [Google Scholar] [CrossRef]
- Sochacki, S.J.; Harper, R.J.; Smettem, K.R.J. Estimation of woody biomass production from a short-rotation bio-energy system in semi-arid Australia. Biomass Bioenergy 2007, 31, 608–616. [Google Scholar] [CrossRef]
- Chen, S.; Arnold, R.; Li, Z.; Li, T.; Zhou, G.; Wu, Z.; Zhou, Q. Tree and stand growth for clonal E. urophylla × grandis across a range of initial stockings in southern China. New For. 2011, 41, 95–112. [Google Scholar] [CrossRef]
- Sartório, I.P. Avaliação e Modelagem do Crescimento de Florestas Energéticas de Eucalipto Plantadas em Diferentes Densidades. Master’s Thesis, Universidade Federal do Paraná, Paraná, Brasil, February 2014. [Google Scholar]
- Cunningham, S.C.; Thomson, J.R.; Read, J.; Baker, P.J.; Mac Nally, R. Does stand structure influence susceptibility of eucalypt floodplain forests to dieback? Austral Ecol. 2010, 35, 348–356. [Google Scholar] [CrossRef]
- Binkley, D.; Stape, J.L.; Ryan, M.G.; Barnard, H.R.; Fownes, J. Age-related decline in forest ecosystem growth: An individual-tree, stand-structure hypothesis. Ecosystems 2002, 5, 58–67. [Google Scholar] [CrossRef]
- Campoe, O.C.; Stape, J.L.; Albaugh, T.J.; Lee Allen, H.; Fox, T.R.; Rubilar, R.; Binkley, D. Fertilization and irrigation effects on tree level aboveground net primary production, light interception and light use efficiency in a loblolly pine plantation. For. Ecol. Manag. 2013, 288, 43–48. [Google Scholar] [CrossRef]
- Berger, U.; Hildenbrandt, H.; Grimm, V. Age-related decline in forest productivity: Modelling the role of neighbourhood competition. J. Ecol. 2004, 92, 846–853. [Google Scholar] [CrossRef]
- Altamirano, A.; da Silva, H.; Durán, A.; Echeverría, A.; Panario, D.; Puentes, R. Dirección de Suelos y Fertilizantes, Carta de Reconocimiento de Suelos del Uruguay; Tomo III; Descripción de las Unidades de Suelos—Charter Soil Survey of Uruguay. Volume III. Description of Units Soil; Dirección de Suelos y Fertilizantes, Ministerio de Agricultura y Pesca: Montevideo, Uruguay, 1979. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Degrees of Freedom |
|---|---|
| Blocks | B − 1 |
| Species | S − 1 |
| Error (Blocks × Species) | (B − 1) × (S − 1) |
| Planting density | Pd − 1 |
| Species × Planting density | (S − 1) × (Pd − 1) |
| Error (Blocks × Species × Planting density) | (B − 1) × (S − 1) × (Pd − 1) |
| Total | (B × S × Pd) − 1 |
| Effects | Variables | Sites | |
|---|---|---|---|
| Tacuarembó | Paysandú | ||
| p | p | ||
| Species | dbh | 0.642 | 0.055 |
| Ht | 0.401 | 0.082 | |
| slenderness | 0.363 | 0.002 | |
| Vi | 0.913 | 0.030 | |
| Vt | 0.353 | 0.031 | |
| Planting density | dbh | <0.001 | <0.001 |
| Ht | <0.001 | <0.001 | |
| slenderness | <0.001 | 0.001 | |
| Vi | <0.001 | <0.001 | |
| Vt | <0.001 | 0.071 | |
| Species × Planting density | dbh | 0.081 | 0.916 |
| Ht | 0.636 | <0.001 | |
| slenderness | 0.093 | 0.006 | |
| Vi | 0.099 | 0.170 | |
| Vt | 0.016 | 0.102 | |
| Sites | Species | Density (trees ha−1) | Ht (m) | dbh (cm) | Slenderness Ht/dbh | Volume (m3 tree−1) | Volume (m3 ha−1) | MAI (m3 ha−1 year−1) | CAI (m3 ha−1 year−1) |
|---|---|---|---|---|---|---|---|---|---|
| Tacuarembó | E. benthamii | 2220 | 17.5(0.2) | 14.9(0.9) | 1.18(0.030) | 0.17(0.016) | 0.17(0.016) | 43.3(1.3) | 44.2(6.2) |
| 3330 | 15.6(0.6) | 11.9(0.1) | 1.31(0.043) | 0.11(0.02) | 0.11(0.02) | 47.7(5.3) | 48.1(4.1) | ||
| 4440 | 15.5 (0.8) | 11.8(1.0) | 1.31(0.019) | 0.11(0.02) | 0.11(0.02) | 68.9(5.6) | 60.5(12.4) | ||
| 6660 | 14.1 (0.4) | 10.3(0.9) | 1.37(0.053) | 0.09(0.017) | 0.09(0.017) | 87.7(9.5) | 87.2(22.4) | ||
| E. dunnii | 2220 | 16.2(0.1) | 15.1(0.3) | 1.07(0.016) | 0.17(0.008) | 0.17(0.008) | 43.3(0.9) | 43.7(6.0) | |
| 3330 | 16.5(0.4) | 14.2(0.6) | 1.17(0.151) | 0.14(0.009) | 0.14(0.009) | 53.9(1.3) | 43.0(3.3) | ||
| 4440 | 15.1(0.5) | 11.9(0.4) | 1.26(0.124) | 0.10(0.009) | 0.10(0.009) | 56.4(3.1) | 40.9(0.9) | ||
| 6660 | 13.7(0.7) | 9.4(0.3) | 1.45(0.077) | 0.07(0.006) | 0.07(0.006) | 61.9(3.2) | 51.9(12.7) | ||
| E. grandis | 2220 | 16.8(0.5) | 15.6(0.2) | 1.08(0.061) | 0.17(0.009) | 0.17(0.009) | 49.6(3.6) | 54.8(10.2) | |
| 3330 | 15.3(0.6) | 13.5(0.7) | 1.14(0.032) | 0.13(0.007) | 0.13(0.007) | 51.2(3.4) | 59.0(3.3) | ||
| 4440 | 15.8(1.2) | 12.7(0.3) | 1.24(0.018) | 0.12(0.007) | 0.12(0.007) | 62.5(2.4) | 64.9(9.0) | ||
| 6660 | 13.6(0.6) | 11.2(0.2) | 1.21(0.013) | 0.09(0.004) | 0.09(0.004) | 62.0(3.0) | 47.5(10.3) | ||
| Paysandú | E. benthamii | 2220 | 17.6(0.3) | 12.6(0.3) | 1.40(0.030) | 0.13(0.005) | 0.13(0.005) | 49.4(1.8) | 8.9(16.8) |
| 3330 | 14.4(0.4) | 10.8(0.1) | 1.34(0.043) | 0.08(0.002) | 0.08(0.002) | 50.5(3.8) | 54.2(29.7) | ||
| 4440 | 14.3(0.4) | 9.9(0.2) | 1.44(0.019) | 0.08(0.003) | 0.08(0.003) | 56.7(2.7) | 20.0(18.6) | ||
| 6660 | 11.9(0.1) | 8.3(0.4) | 1.44((0.053) | 0.05(0.005) | 0.05(0.005) | 57.4(4.0) | 49.7(17.0) | ||
| E. dunnii | 2220 | 17.4(0.8) | 12.7(0.3) | 1.37(0.016) | 0.14(0.008) | 0.14(0.008) | 53.9(3.3) | 15.7(8.4) | |
| 3330 | 17.2(0.7) | 10.4(0.4) | 1.65(0.151) | 0.09(0.009) | 0.09(0.009) | 56.8(5.1) | 16.5(7.4) | ||
| 4440 | 15.2(0.5) | 9.2(0.4) | 1.65(0.124) | 0.07(0.007) | 0.07(0.007) | 51.7(5.4) | 14.7(11.9) | ||
| 6660 | 14.7(0.3) | 7.8(0.2) | 1.88(0.077) | 0.05(0.004) | 0.05(0.004) | 52.4(2.8) | −9.2(8.3.8) | ||
| E. grandis | 2220 | 16.4(0.5) | 13.8(0.9) | 1.19(0.061) | 0.16(0.016) | 0.16(0.016) | 60.1(5.7) | 34.4(8.4) | |
| 3330 | 16.2(0.4) | 12.1(0.1) | 1.34(0.032) | 0.14(0.006) | 0.14(0.006) | 77.1(4.7) | 36.0(18.7) | ||
| 4440 | 15.0(0.6) | 10.6(0.6) | 1.41(0.018) | 0.11(0.014) | 0.11(0.014) | 85.8(6.4) | 71.2(6.0) | ||
| 6660 | 13.2(0.5) | 9.3(0.1) | 1.42(0.013) | 0.08(0.002) | 0.08(0.002) | 90.1(3.0) | 35.2(20.3) |
| Site | Species | Models | R2 | RMSE | Bias | AIC | F | p-Value |
|---|---|---|---|---|---|---|---|---|
| Tacuarembó | E. benthamii | Vi = exp((−9.86876 − 0.29901 × Ln(dbh) + 1.05411 × Ln(dbh2 × Ht)) | 0.99 | 0.009 | 0.0001 | −362 | 15150 | <0.001 |
| Vi = exp((−9.35966 + 2.75971 × Ln(dbh)) | 0.94 | 0.020 | −0.0010 | 121 | 5111 | <0.001 | ||
| Vi = −0.02471 + 0.006340 × (dbh) + 0.00003951 × (dbh2 × Ht) × −0.0003778 × (dbh2) | 0.98 | 0.009 | 0.0048 | −1111 | 4197 | <0.001 | ||
| E. dunnii | Vi = −0.005625 + 0.001523 × (dbh) + 0.00003184 × (dbh2Ht) | 0.98 | 0.002 | 0.0050 | −1019 | 6033 | <0.001 | |
| Vi = exp((−8.38487 + 2.38262 × Ln(dbh)) | 0.95 | 0.015 | 0.0004 | 104 | 5095 | <0.001 | ||
| Vi = exp((−9.86876 − 0.29901 × Ln(dbh) + 1.05411 × Ln(dbh2 × Ht)) | 0.96 | 0.013 | 0.0079 | 208 | 7893 | <0.001 | ||
| E. grandis | Vi = −0.01781 + 0.004755 × (dbh) − 0.0003141 × (dbh2) + 0.00004038 × (dbh2 × Ht) | 0.99 | 0.007 | 0.0042 | −1159 | 8847 | <0.001 | |
| Vi = exp((−9.48276 + 1.38990 × Ln(dbh2)) | 0.96 | 0.017 | 0.0001 | 118 | 8696 | <0.001 | ||
| Vi = exp((−9.79678 + 0.12407 × Ln(dbh) + 0.91076 × Ln(dbh2 × Ht)) | 0.99 | 0.008 | 0.0005 | 236 | 31360 | <0.001 | ||
| Paysandú | E. benthamii | Vi = exp((−9.711769 + 0.936637 × Ln(dbh2 × Ht)) | 0.99 | 0.005 | 0.0002 | −441 | 44070 | <0.001 |
| Vi = exp((−8.99697 + 02.65528 × Ln(dbh)) | 0.96 | 0.011 | 0.0040 | −1510 | 5885 | <0.001 | ||
| Vi = exp((−9.70756 + 0.01996 × Ln(dbh) + 0.92976 × Ln(dbh2 × Ht)) | 0.99 | 0.005 | 0.0002 | −441 | 21910 | <0.001 | ||
| E. dunnii | Vi = 0.02608 − 0.009986 × (dbh) + 0.001309 × (dbh2) | 0.98 | 0.009 | 0.0068 | −1093 | 3177 | <0.001 | |
| Vi = exp((−9.01150 + 2.65731 × Ln(dbh)) | 0.95 | 0.010 | 0.0020 | 116 | 4011 | <0.001 | ||
| Vi = exp((−9.01550 + 1.32866 × Ln(dbh2)) | 0.96 | 0.009 | 0.0016 | 116 | 4011 | <0.001 | ||
| E. grandis | Vi= −0.006259 + 0.002463 × (dbh) + 0.00004120 × (dbh2 × Ht) − 0.000226 × (dbh2) | 0.99 | 0.004 | 0.0026 | −1371 | 22980 | <0.001 | |
| Vi = exp((−9.25879 + 2.75792 × Ln(dbh)) | 0.96 | 0.015 | 0.0001 | 116 | 6477 | <0.001 | ||
| Vi = exp((−9.79678 + 0.12407 × Ln(dbh) + 0.91076 × Ln(dbh2 × Ht)) | 0.99 | 0.005 | 0.0005 | 233 | 28120 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Resquin, F.; Navarro-Cerrillo, R.M.; Rachid-Casnati, C.; Hirigoyen, A.; Carrasco-Letelier, L.; Duque-Lazo, J. Allometry, Growth and Survival of Three Eucalyptus Species (Eucalyptus benthamii Maiden and Cambage, E. dunnii Maiden and E. grandis Hill ex Maiden) in High-Density Plantations in Uruguay. Forests 2018, 9, 745. https://doi.org/10.3390/f9120745
Resquin F, Navarro-Cerrillo RM, Rachid-Casnati C, Hirigoyen A, Carrasco-Letelier L, Duque-Lazo J. Allometry, Growth and Survival of Three Eucalyptus Species (Eucalyptus benthamii Maiden and Cambage, E. dunnii Maiden and E. grandis Hill ex Maiden) in High-Density Plantations in Uruguay. Forests. 2018; 9(12):745. https://doi.org/10.3390/f9120745
Chicago/Turabian StyleResquin, Fernando, Rafael M. Navarro-Cerrillo, Cecilia Rachid-Casnati, Andrés Hirigoyen, Leonidas Carrasco-Letelier, and Joaquín Duque-Lazo. 2018. "Allometry, Growth and Survival of Three Eucalyptus Species (Eucalyptus benthamii Maiden and Cambage, E. dunnii Maiden and E. grandis Hill ex Maiden) in High-Density Plantations in Uruguay" Forests 9, no. 12: 745. https://doi.org/10.3390/f9120745
APA StyleResquin, F., Navarro-Cerrillo, R. M., Rachid-Casnati, C., Hirigoyen, A., Carrasco-Letelier, L., & Duque-Lazo, J. (2018). Allometry, Growth and Survival of Three Eucalyptus Species (Eucalyptus benthamii Maiden and Cambage, E. dunnii Maiden and E. grandis Hill ex Maiden) in High-Density Plantations in Uruguay. Forests, 9(12), 745. https://doi.org/10.3390/f9120745