High-Efficiency Somatic Embryogenesis from Seedlings of Koelreuteria paniculata Laxm.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Initiation
2.2. Embryogenic Callus Induction and Proliferation
2.3. Somatic Embryogenesis and Maturation
2.4. Histology
2.5. Clonal Culture
2.6. Statistical Analysis
3. Results
3.1. Embryogenic Callus Induction and Proliferation
3.2. Somatic Embryogenesis and Maturation
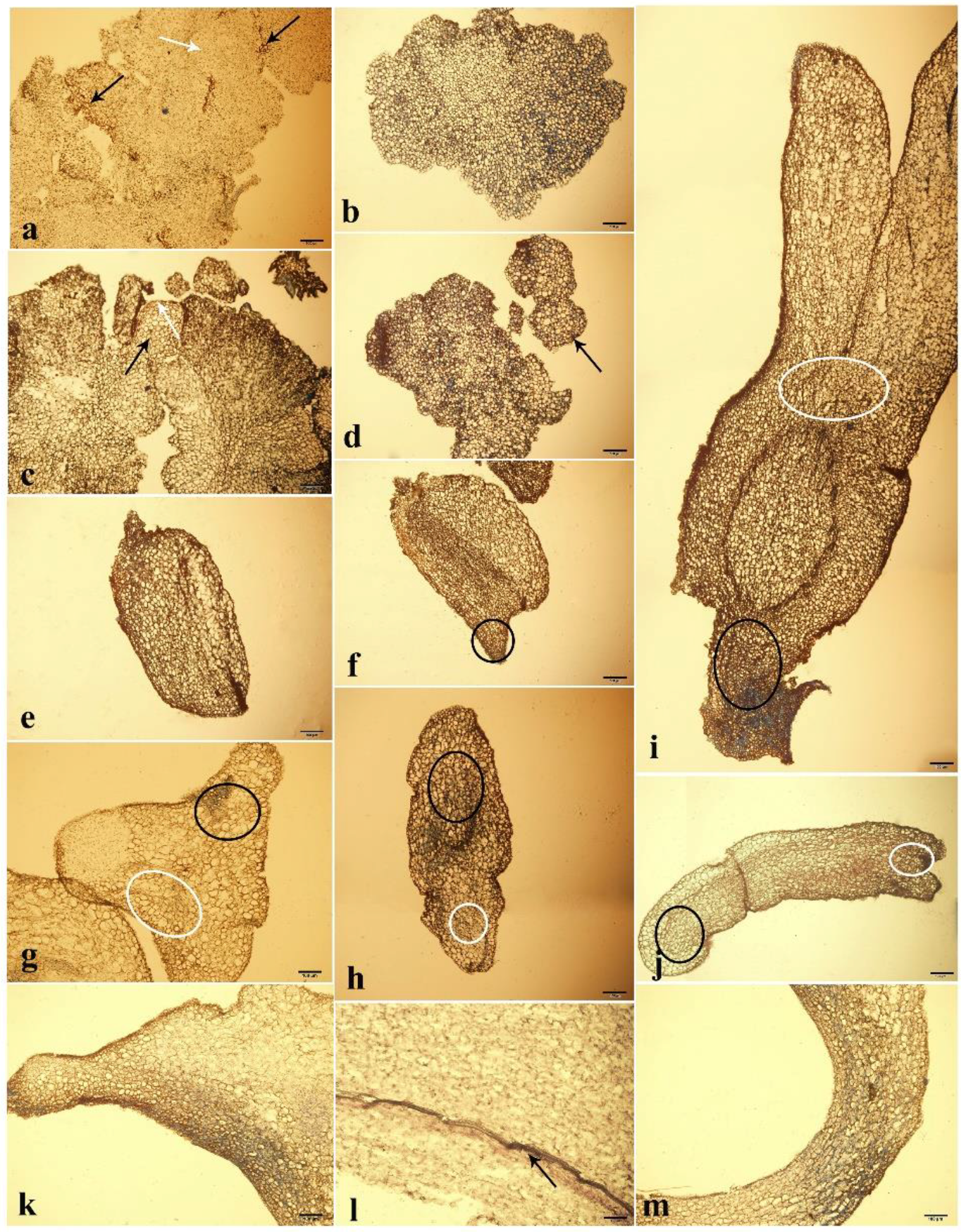
3.3. Morphology and Histology Analysis
3.4. Clonal Culture
4. Discussion
4.1. Effect of Plant Growth Regulators on Somatic Embryogenesis
4.2. Effect of Basic Media on Somatic Embryogenesis
4.3. Other Factors Influencing Somatic Embryogenesis
4.4. Morphology and Histology Analysis
4.5. Clonal Culture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kamala-Kannan, S.; Park, S.-M.; Oh, B.-T.; Kim, H.-M.; Lee, K.-J. First Report of Aster Yellows Phytoplasma in Goldenrain Tree (Koelreuteria paniculata) in Korea. J. Phytopathol. 2010, 158, 197–199. [Google Scholar] [CrossRef]
- Rehman, S.; Park, I.-H. Effect of scarification, GA and chilling on the germination of goldenrain-tree (Koelreuteria paniculata Laxm.) seeds. Sci. Hortic. 2000, 85, 319–324. [Google Scholar] [CrossRef]
- Mostafa, A.E.; El-Hela, A.A.; Mohammad, A.I.; Cutler, S.J.; Ross, S.A. New triterpenoidal saponins from Koelreuteria paniculata. Phytochem. Lett. 2016, 17, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Mikolajczak, K.L.; Smith, C.R.; Tjarks, L.W. Cyanolipids of Koelreuteria paniculata Laxm. seed oil. Lipids 1970, 5, 672–677. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Damico, D.C.S.; Freire, M.D.G.M.; Toyama, M.H.; Marangoni, S.; Novello, J.C. Purification and characterization of an N-acetylglucosamine-binding lectin from Koelreuteria paniculata seeds and its effect on the larval development of Callosobruchus maculatus (Coleoptera: Bruchidae) and Anagasta kuehniella (Lepidoptera: Pyralidae). J. Agric. Food Chem. 2003, 51, 2980–2986. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.H.; Deng, Z.W.; Lei, H.M.; Fu, H.Z.; Li, J. Polyphenolic compounds from the leaves of Koelreuteria paniculata Laxm. J. Asian Nat. Prod. Res. 2002, 4, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Sutiashvili, M.G.; Alaniya, M.D.; Mshvildadze, V.D.; Skhirtladze, A.V.; Pichette, A.; Lavoie, S. Flavonoid and cycloartane glycosides from seeds of Koelreuteria paniculata. Chem. Nat. Compd. 2013, 49, 395–397. [Google Scholar] [CrossRef]
- Tian, D.; Zhu, F.; Yan, W.; Fang, X.; Xiang, W.; Deng, X.; Wang, G.; Peng, C. Heavy metal accumulation by panicled goldenrain tree (Koelreuteria paniculata) and common elaeocarpus (Elaeocarpus decipens) in abandoned mine soils in southern China. J. Environ. Sci. 2009, 21, 340–345. [Google Scholar] [CrossRef]
- Ocokoljić, M.; Medarević, M.; Nikić, Z.; Galečić, N.; Stojičić, Đ. Variability of features in half-sib posterity as a basis in plant breeding of the species Koelreuteria paniculata Laxm. Arch. Biol. Sci. 2010, 62, 693–703. [Google Scholar] [CrossRef]
- Ertekın, M. Effects of microorganisms, hormone treatment and stratification on seed germination of goldenrain tree (Koelreuteria paniculata). Int. J. Agric. Biol. 2011, 13, 38–42. [Google Scholar]
- Sevik, H.; Çetin, M. Effects of Water Stress on Seed Germination for Select Landscape Plants. Pol. J. Environ. Stud. 2015, 24, 689–693. [Google Scholar] [CrossRef]
- Rosa, Y.B.C.J.; Bello, C.C.M.; Dornelas, M.C. Species-dependent divergent responses to in vitro somatic embryo induction in Passiflora spp. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 120, 69–77. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Bandupriya, H.D.D.; López-Villalobos, A.; Sisunandar, S.; Foale, M.; Adkins, S.W. Tissue culture and associated biotechnological interventions for the improvement of coconut (Cocos nucifera L.): A review. Planta 2015, 242, 1059–1076. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Shirakawa, M.; Koumoto, Y.; Honda, M.; Asami, Y.; Kondo, Y.; Hara-Nishimura, I. A simple and reliable multi-gene transformation method for switchgrass. Plant Cell Rep. 2014, 33, 1161–1172. [Google Scholar] [CrossRef] [PubMed]
- Teixeira da Silva, J.A.; Zeng, S.; Galdiano, R.F.; Dobránszki, J.; Cardoso, J.C.; Vendrame, W.A. In vitro conservation of Dendrobium germplasm. Plant Cell Rep. 2014, 33, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Quiroz-Figueroa, F.R.; Rojas-Herrera, R.; Galaz-Avalos, R.M.; Loyola-Vargas, V.M. Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell Tissue Organ Cult. (PCTOC) 2006, 86, 285. [Google Scholar] [CrossRef]
- Corredoira, E.; Ballester, A.; Ibarra, M.; Vieitez, A.M. Induction of somatic embryogenesis in explants of shoot cultures established from adult Eucalyptus globulus and E. saligna × E. maidenii trees. Tree Physiol. 2015, 35, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.L. Somatic Embryogenesis: A Model for Early Development in Higher Plants. Plant Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef] [PubMed]
- Jariteh, M.; Ebrahimzadeh, H.; Niknam, V.; Mirmasoumi, M.; Vahdati, K. Developmental changes of protein, proline and some antioxidant enzymes activities in somatic and zygotic embryos of Persian walnut (Juglans regia L.). Plant Cell Tissue Organ Cult. (PCTOC) 2015, 122, 101–115. [Google Scholar] [CrossRef]
- Driver, J.A.; Kuniyuki, A.H. In Vitro Propagation of Paradox Walnut Rootstock [Juglans hindsii × Juglans regia, Tissue Culture]; HortScience: Pleasanton, CA, USA, 1984. [Google Scholar]
- Sabooni, N.; Shekafandeh, A. Somatic embryogenesis and plant regeneration of blackberry using the thin cell layer technique. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 130, 313–321. [Google Scholar] [CrossRef]
- Ahn, C.-H.; Tull, A.R.; Montello, P.M.; Merkle, S.A. A clonal propagation system for Atlantic white cedar (Chamaecyparis thyoides) via somatic embryogenesis without the use of plant growth regulators. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 130, 91–101. [Google Scholar] [CrossRef]
- Mukhopadhyay, M.; Mondal, T.K.; Chand, P.K. Biotechnological advances in tea (Camellia sinensis [L.] O. Kuntze): A review. Plant Cell Rep. 2016, 35, 255–287. [Google Scholar] [CrossRef] [PubMed]
- Isah, T. Induction of somatic embryogenesis in woody plants. Acta Physiol. Plant. 2016, 38, 118. [Google Scholar] [CrossRef]
- Leljak-Levanić, D.; Mihaljević, S.; Bauer, N. Somatic and zygotic embryos share common developmental features at the onset of plant embryogenesis. Acta Physiol. Plant. 2015, 37, 127. [Google Scholar] [CrossRef]
- Żur, I.; Dubas, E.; Krzewska, M.; Waligórski, P.; Dziurka, M.; Janowiak, F. Hormonal requirements for effective induction of microspore embryogenesis in triticale (× Triticosecale Wittm.) anther cultures. Plant Cell Rep. 2015, 34, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, M.; Susanthi, B.; Mohan, N.M.; Mandal, P.K. In vitro somatic embryogenesis and plantlet regeneration from immature male inflorescence of adult dura and tenera palms of Elaeis guineensis (Jacq.). SpringerPlus 2015, 4, 256. [Google Scholar] [CrossRef]
- Prakash, M.G.; Gurumurthi, K. Effects of type of explant and age, plant growth regulators and medium strength on somatic embryogenesis and plant regeneration in Eucalyptus camaldulensis. Plant Cell Tissue Organ Cult. (PCTOC) 2009, 100, 13. [Google Scholar] [CrossRef]
- Nunes, S.; Marum, L.; Farinha, N.; Pereira, V.T.; Almeida, T.; Sousa, D.; Mano, N.; Figueiredo, J.; Dias, M.C.; Santos, C. Somatic embryogenesis of hybrid Pinus elliottii var. elliottii × P. caribaea var. hondurensis and ploidy assessment of somatic plants. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 71–84. [Google Scholar] [CrossRef]
- Pullman, G.S.; Bucalo, K. Pine somatic embryogenesis: Analyses of seed tissue and medium to improve protocol development. New For. 2014, 45, 353–377. [Google Scholar] [CrossRef]
- Santos, R.S.; Labory, C.R.G.; Stein, V.C.; Mendonça, E.G.; Alves, E.; Paiva, L.V. Histodifferentiation of oil palm somatic embryo development at low auxin concentration. Protoplasma 2018, 255, 285–295. [Google Scholar]
- Ali, M.; Mujib, A.; Tonk, D.; Zafar, N. Plant regeneration through somatic embryogenesis and genome size analysis of Coriandrum sativum L. Protoplasma 2017, 254, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Viñas, S.A.; Lecona-Guzmán, C.A.; Canto-Flick, A.; López-Erosa, S.; Santana-Buzzy, N. Morpho-histological and ultrastructural study on direct somatic embryogenesis of Capsicum chinense Jacq. in liquid medium. Plant Biotechnol. Rep. 2013, 7, 277–286. [Google Scholar] [CrossRef]
- Kim, Y.-W. Initiation of embryogenic callus from mature zygotic embryos in Japanese larch (Larix kaempferi). J. Plant Biotechnol. 2015, 42, 223–227. [Google Scholar] [CrossRef]
- Martínez, M.T.; San José, M.C.; Vieitez, A.M.; Cernadas, M.J.; Ballester, A.; Corredoira, E. Propagation of mature Quercus ilex L. (holm oak) trees by somatic embryogenesis. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 131, 321–333. [Google Scholar] [CrossRef]
- Wu, Y.; Dor, E.; Hershenhorn, J. Strigolactones affect tomato hormone profile and somatic embryogenesis. Planta 2017, 245, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Remakanthan, A.; Menon, T.G.; Soniya, E.V. Somatic embryogenesis in banana (Musa acuminata AAA cv. Grand Naine): Effect of explant and culture conditions. Vitr. Cell. Dev. Biol.–Plant 2014, 50, 127–136. [Google Scholar] [CrossRef]
- Kumar, V.; Moyo, M.; Van Staden, J. Somatic embryogenesis of Pelargonium sidoides DC. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 121, 571–577. [Google Scholar] [CrossRef]
- Mikuła, A.; Pożoga, M.; Tomiczak, K.; Rybczyński, J.J. Somatic embryogenesis in ferns: A new experimental system. Plant Cell Rep. 2015, 34, 783–794. [Google Scholar] [CrossRef]
- SanÉ, D.; Aberlenc-Bertossi, F.; Gassama-Dia, Y.K.; Sagna, M.; Trouslot, M.F.; Duval, Y.; Borgel, A. Histocytological analysis of callogenesis and somatic embryogenesis from cell suspensions of date palm (Phoenix dactylifera). Ann. Bot. 2006, 98, 301–308. [Google Scholar] [CrossRef]
- Pullman, G.S.; Zeng, X.; Copeland-Kamp, B.; Crockett, J.; Lucrezi, J.; May, S.W.; Bucalo, K. Conifer somatic embryogenesis: Improvements by supplementation of medium with oxidation–reduction agents. Tree Physiol. 2015, 35, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Ikeda-Iwai, M.; Umehara, M.; Satoh, S.; Kamada, H. Stress-induced somatic embryogenesis in vegetative tissues of Arabidopsis thaliana. Plant J. 2003, 34, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Soundar Raju, C.; Kathiravan, K.; Aslam, A.; Shajahan, A. An efficient regeneration system via somatic embryogenesis in mango ginger (Curcuma amada Roxb.). Plant Cell. Tissue Organ Cult. (PCTOC) 2013, 112, 387–393. [Google Scholar] [CrossRef]
- Cheruvathur, M.K.; Kumar, G.K.; Thomas, T.D. Somatic embryogenesis and synthetic seed production in Rhinacanthus nasutus (L.) Kurz. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 113, 63–71. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Lelu-Walter, M.-A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Niskanen, A.M.; Lu, J.; Seitz, S.; Keinonen, K.; von Weissenberg, K.; Pappinen, A. Effect of parent genotype on somatic embryogenesis in Scots pine (Pinus sylvestris). Tree Physiol. 2004, 24, 1259–1265. [Google Scholar] [CrossRef]
- Testillano, P.S.; Gómez-Garay, A.; Pintos, B.; Risueño, M.C. Somatic Embryogenesis of Quercus suber L. from Immature Zygotic Embryos. In Plant Cell Culture Protocols; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Springer New York: New York, NY, USA, 2018; pp. 247–256. [Google Scholar]
- San Pedro, T.; Gammoudi, N.; Peiró, R.; Olmos, A.; Gisbert, C. Somatic embryogenesis from seeds in a broad range of Vitis vinifera L. varieties: Rescue of true-to-type virus-free plants. BMC Plant Biol. 2017, 17, 226. [Google Scholar] [CrossRef]
- Corredoira, E.; Valladares, S.; Martínez, M.T.; Vieitez, A.M.; San José, M.C. Somatic embryogenesis in Alnus glutinosa (L.) Gaertn. Trees 2013, 27, 1597–1608. [Google Scholar] [CrossRef]
- Sunandar, A.; Supena, E.D.J. Induction of somatic embryogenesis in sengon (Falcataria moluccana) with thidiazuron and light treatments. HAYATI J. Biosci. 2017, 24, 105–108. [Google Scholar] [CrossRef]
- El-Mahrouk, M.; Dewir, Y.H.; Omar, A.M.K. In vitro propagation of adult strawberry tree (Arbutus unedo L.) through adventitious shoots and somatic embryogenesis. Propag. Ornam. Plants 2010, 10, 93–98. [Google Scholar]
- Stasolla, C.; Yeung, E.C. Recent advances in conifer somatic embryogenesis: Improving somatic embryo quality. Plant Cell Tissue Organ Cult. 2003, 74, 15–35. [Google Scholar] [CrossRef]
- Gomes, H.T.; Bartos, P.M.C.; Scherwinski-Pereira, J.E. Dynamics of morphological and anatomical changes in leaf tissues of an interspecific hybrid of oil palm during acquisition and development of somatic embryogenesis. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 131, 269–282. [Google Scholar] [CrossRef]
- Soundar Raju, C.; Aslam, A.; Shajahan, A. High-efficiency direct somatic embryogenesis and plant regeneration from leaf base explants of turmeric (Curcuma longa L.). Plant Cell Tissue Organ Cult. (PCTOC) 2015, 122, 79–87. [Google Scholar] [CrossRef]
- Cantelmo, L.; Soares, B.O.; Rocha, L.P.; Pettinelli, J.A.; Callado, C.H.; Mansur, E.; Castellar, A.; Gagliardi, R.F. Repetitive somatic embryogenesis from leaves of the medicinal plant Petiveria alliacea L. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 115, 385–393. [Google Scholar] [CrossRef]
- Stevens, M.E.; Pijut, P.M. Rapid in vitro shoot multiplication of the recalcitrant species Juglans nigra L. Vitr. Cell. Dev. Biol.–Plant 2018, 54, 309–317. [Google Scholar] [CrossRef]
- García-Mendiguren, O.; Montalbán, I.A.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Environmental conditions at the initial stages of Pinus radiata somatic embryogenesis affect the production of somatic embryos. Trees 2016, 30, 949–958. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Basic Medium | PGR (mg L−1) | Calluses Induction Frequency + SE (%) | Cultured Conditions | ||
|---|---|---|---|---|---|---|
| BA | NAA | 2,4-D | ||||
| I1 | MS | 0.25 | 0.25 | 0.50 | 40.47 + 6.47 bc | rhizogenesis |
| I2 | MS | 0.50 | 0.50 | 1.00 | 51.87 + 6.47 b | non-embryogenic callus |
| I3 | MS | 0.75 | 0.75 | 1.50 | 0.00 + 00.00 d | browning |
| I4 | B5 | 0.25 | 0.50 | 1.50 | 25.90 + 6.41 c | rhizogenesis |
| I5 | B5 | 0.50 | 0.75 | 0.50 | 40.70 + 6.41 bc | rhizogenesis |
| I6 | B5 | 0.75 | 0.25 | 1.00 | 29.60 + 6.41 c | non-embryogenic callus |
| I7 | DKW | 0.25 | 0.75 | 1.00 | 85.20 + 6.41 a | rhizogenesis |
| I8 | DKW | 0.50 | 0.25 | 1.50 | 81.50 + 6.41 a | embryogenic callus |
| I9 | DKW | 0.75 | 0.50 | 0.50 | 77.80 + 11.10 a | embryogenic callus |
| Number | PGR (mg L−1) | Calluses Induction Frequency ± SE (%) | ||
|---|---|---|---|---|
| BA | NAA | 2,4-D | ||
| IEM1 | 0.50 | 0.25 | 1.50 | 80.25 ± 0.08 |
| IEM2 | 0.75 | 0.25 | 1.50 | 70.25 ± 0.09 |
| IEM3 | 0.50 | 0.50 | 0.50 | 69.50 ± 0.04 |
| IEM4 | 0.75 | 0.50 | 0.50 | 74.25 ± 0.05 |
| Plant Growth Regulators (mg L−1) | Frequency of Somatic Embryogenesis ± SE (%) | Number of Somatic Embryos |
|---|---|---|
| 2,4-D | ||
| 0 | 25.50 ± 0.04 bc | * |
| 0.50 | 23.25 ± 0.08 c | ** |
| 1.00 | 46.75 ± 0.05 ab | *** |
| 1.50 | 32.50 ± 0.12 abc | ** |
| NAA | ||
| 0.10 | 54.75 ± 0.16 a | *** |
| 0.20 | 37.75 + 0.09 abc | *** |
| NAA (mg L−1) | Condition of Culture | Length of Hypocotyl ± SE (cm) | Length of Cotyledon ± SE (cm) |
|---|---|---|---|
| 0.1 | Light | 1.19 ± 0.12 b | 0.53 ± 0.10 |
| 0.2 | Light | 1.50 ± 0.13 a | 0.58 ± 0.08 |
| 0.1 | Dark | 0.44 ± 0.10 d | 0.49 ± 0.08 |
| 0.2 | Dark | 0.64 ± 0.09 c | 0.51 ± 0.09 |
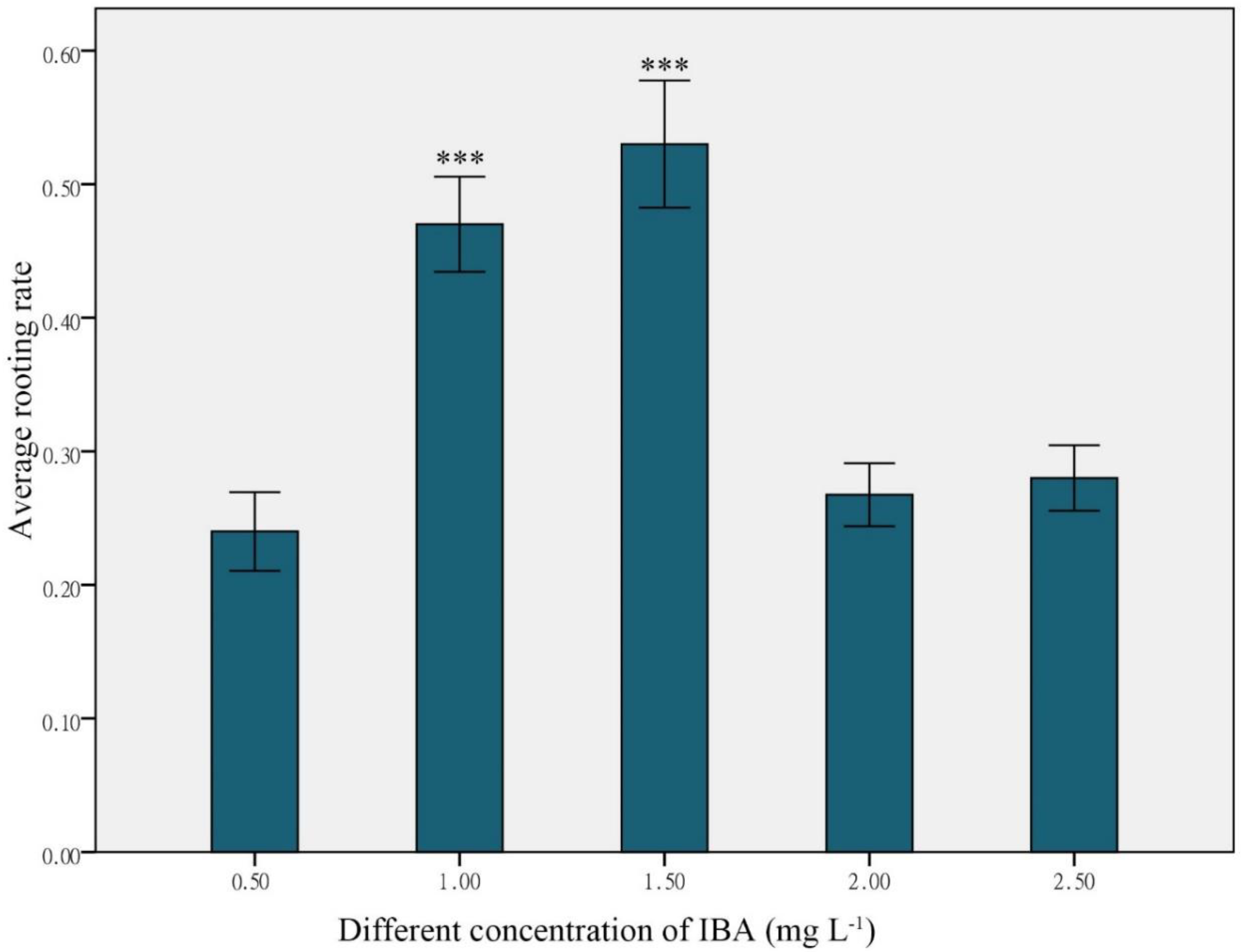
| IBA (mg L−1) | NAA (mg L−1) | Rooting Rate ± SE (%) |
|---|---|---|
| 1.0 | 0 | 47.67 ± 0.10 |
| 1.5 | 0 | 52.00 ± 0.13 |
| 1.0 | 0.1 | 42.67 ± 0.09 |
| 1.0 | 0.2 | 37.33 ± 0.03 |
| 1.0 | 0.3 | 33.17 ± 0.04 |
| 1.5 | 0.1 | 49.67 ± 0.05 |
| 1.5 | 0.2 | 37.67 ± 0.04 |
| 1.5 | 0.3 | 39.67 ± 0.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Yang, X.; Guo, T.; Gao, K.; Zhao, T.; Chen, Z.; An, X. High-Efficiency Somatic Embryogenesis from Seedlings of Koelreuteria paniculata Laxm. Forests 2018, 9, 769. https://doi.org/10.3390/f9120769
Yang X, Yang X, Guo T, Gao K, Zhao T, Chen Z, An X. High-Efficiency Somatic Embryogenesis from Seedlings of Koelreuteria paniculata Laxm. Forests. 2018; 9(12):769. https://doi.org/10.3390/f9120769
Chicago/Turabian StyleYang, Xiong, Xiaoyu Yang, Ting Guo, Kai Gao, Tianyun Zhao, Zhong Chen, and Xinmin An. 2018. "High-Efficiency Somatic Embryogenesis from Seedlings of Koelreuteria paniculata Laxm." Forests 9, no. 12: 769. https://doi.org/10.3390/f9120769
APA StyleYang, X., Yang, X., Guo, T., Gao, K., Zhao, T., Chen, Z., & An, X. (2018). High-Efficiency Somatic Embryogenesis from Seedlings of Koelreuteria paniculata Laxm. Forests, 9(12), 769. https://doi.org/10.3390/f9120769