Ecological Stoichiometric Characteristics of Two Typical Plantations in the Karst Ecosystem of Southwestern China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Sample Collection and Preparation
2.3. Chemical Analyses
2.4. Statistical Analyses
3. Results
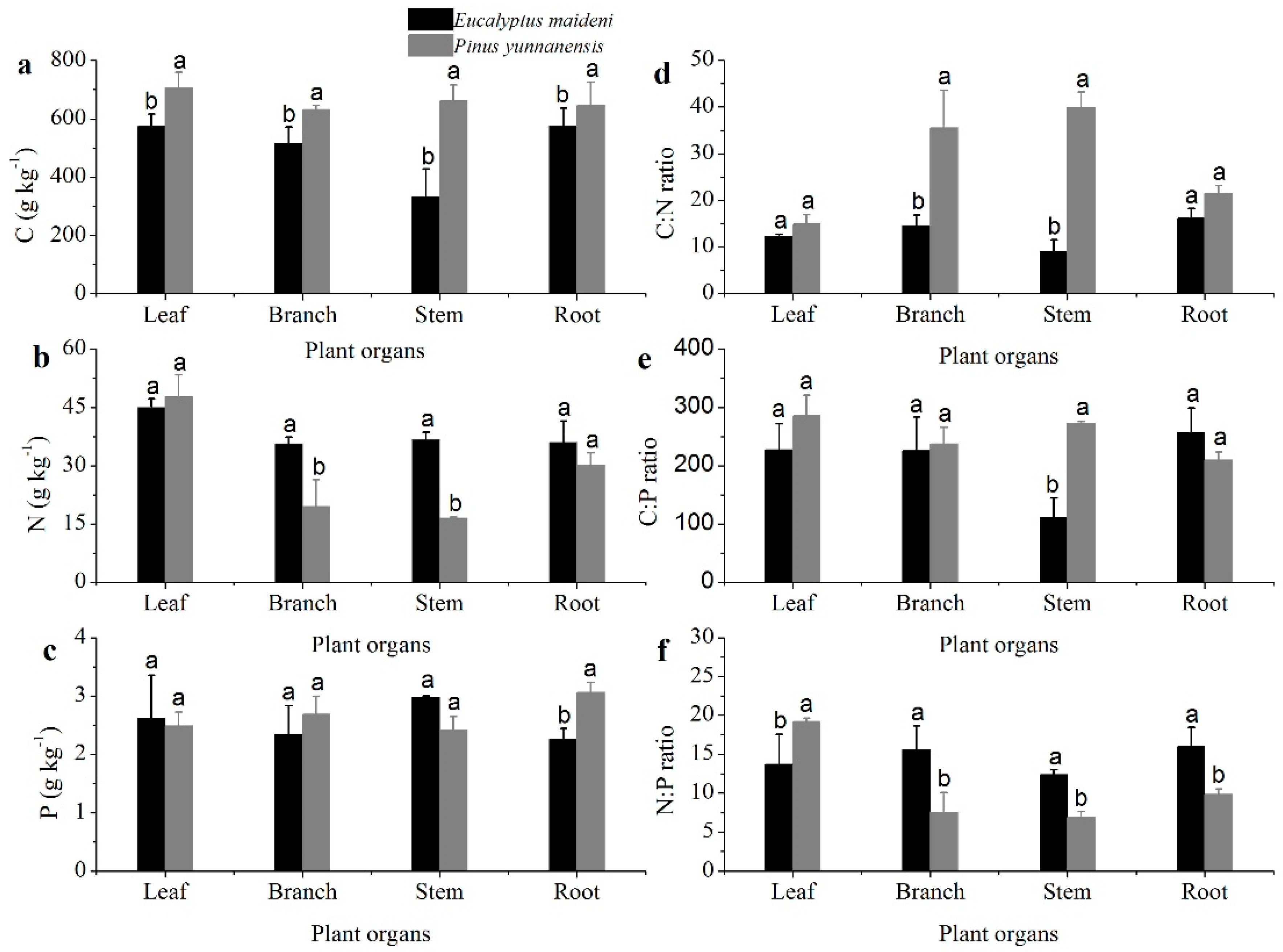
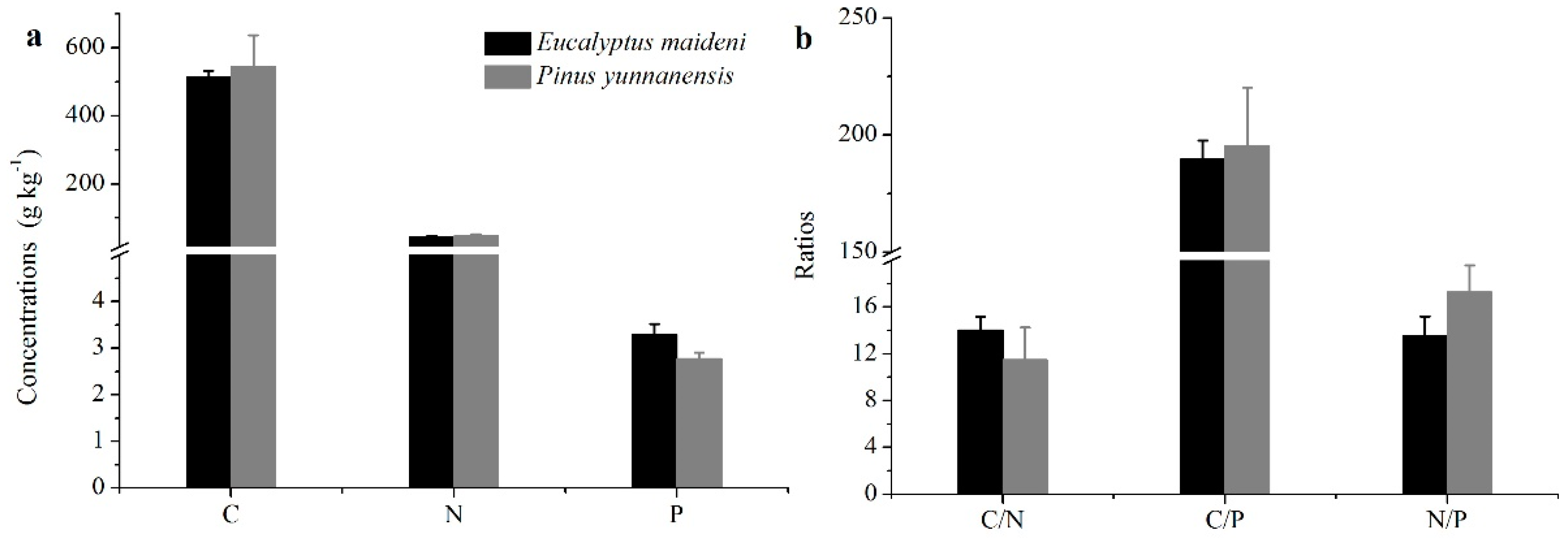
3.1. The Concentrations and Ratios of C, N, and P in Different Plant Organs
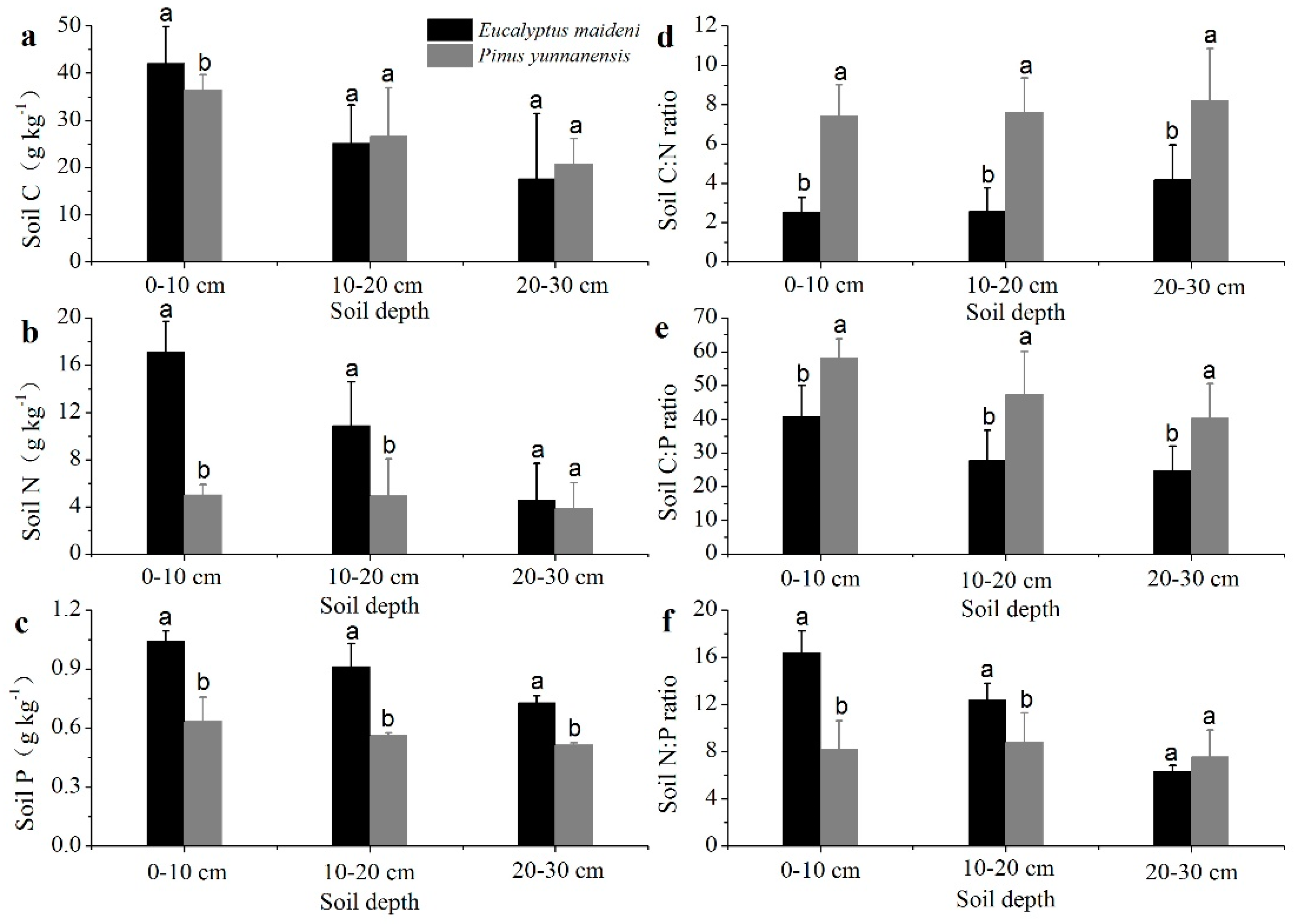
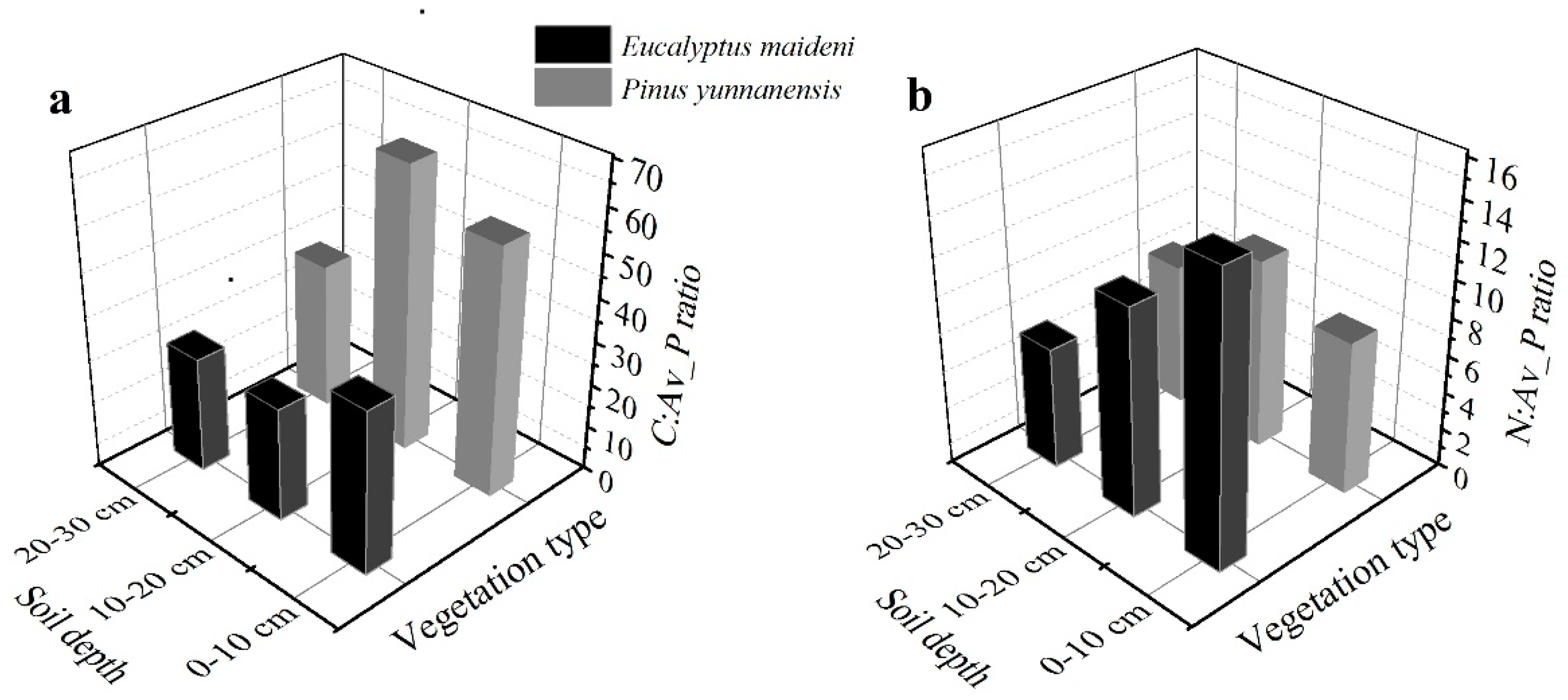
3.2. The Concentrations and Ratios of C, N, and P in Soils
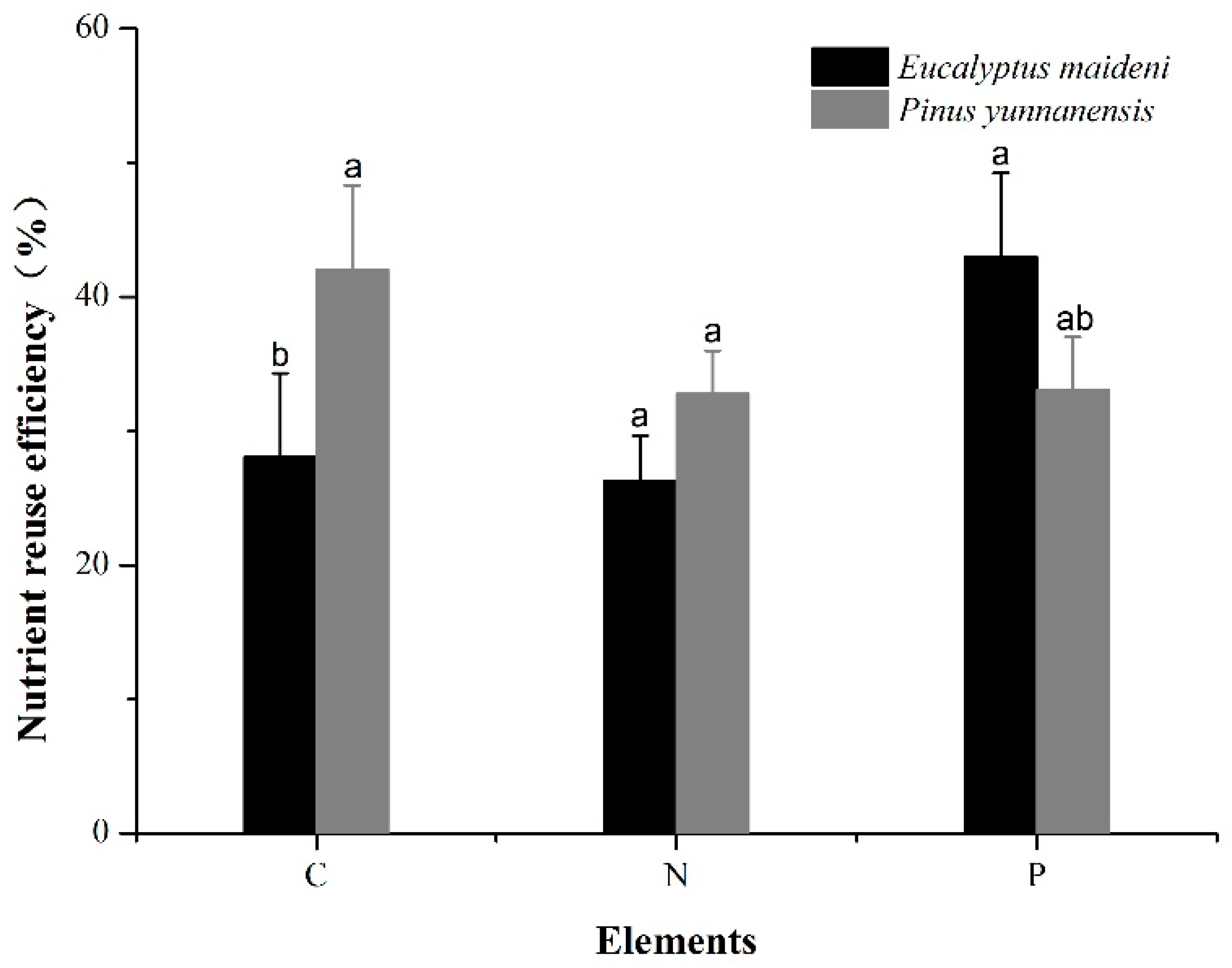
3.3. The Nutrient Resorption Efficiency (NRE) of the Two Different Vegetation Types
4. Discussion
4.1. General Patterns of C, N, and P Concentrations and C:N:P Stoichiometry in Different Plant Organs
4.2. Possible Factors Affecting the Nutrient Concentrations and Stoichiometry of C, N, and P in the Soils
4.3. Nutrient Limitation
4.4. Nutrient Resorption Efficiency (NRE)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, C.C.; Wei, Y.F.; Liu, Y.G.; Guo, K. Biomass of canopy and shrub layers of karst forests in Puding, Guizhou, China. J. Plant Ecol. 2009, 33, 698–705. [Google Scholar]
- Jiang, Z.C.; Lian, Y.; Qin, X. Rocky desertification in Southwest China Impacts, causes, and restoration. Earth-Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Du, Y.X.; Pan, G.X.; Li, L.Q.; Hu, Z.L.; Wang, X.Z. Leaf N/P ratio and nutrient reuse between dominant species and stands: Predicting phosphorus deficiencies in Karst ecosystems, southwestern China. Environ. Earth Sci. 2011, 64, 299–309. [Google Scholar] [CrossRef]
- Behera, N.; Sahani, U. Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. For. Ecol. Manag. 2003, 174, 1–11. [Google Scholar] [CrossRef]
- Zhu, H.H.; He, X.Y.; Wang, K.L.; Su, Y.R.; Wu, J.S. Interaction of vegetation succession, soil biochemical properties and microbial communities in Karst ecosystem. Eur. J. Soil Biol. 2012, 51, 1–7. [Google Scholar] [CrossRef]
- Hofmeister, J.; Mihaljevic, M.; Hosek, J.; Sadlo, J. Eutrophication of deciduous forests in the Bohemian Karst (Czech Republic): The role of nitrogen and phosphorus. For. Ecol. Manag. 2002, 169, 213–230. [Google Scholar] [CrossRef]
- Hatano, R.; Shinano, T.; Zheng, T.G.; Okubo, M.; Li, Z.W. Nitrogen budgets and environmental capacity in farm systems in a largescale karst region, southern China. Nutr. Cycl. Agroecosyst. 2002, 63, 139–149. [Google Scholar] [CrossRef]
- Niinemets, U.; Kull, K. Co-limitation of plant primary productivity by nitrogen and phosphorus in a species-rich wooded meadow on calcareous soils. Acta Oecol. 2005, 28, 345–356. [Google Scholar] [CrossRef]
- Pan, F.J.; Zhang, W.; Liu, S.J.; Wang, K. Leaf N:P stoichiometry across plant functional groups in the karst region of southwestern China. Trees 2015, 29, 883–892. [Google Scholar] [CrossRef]
- Kirkby, C.A.; Kirkegaard, J.A.; Richardson, A.E.; Wade, L.J.; Blanchard, C.; Batten, G. Stable soil organic matter: A comparison of C:N:P:S ratios in Australian and other world soils. Geoderma 2011, 163, 197–208. [Google Scholar] [CrossRef]
- Ågren, G.I.; Wetterstedt, J.Å.; Billberger, M.F.K. Nutrient limitation on terrestrial plant growth—modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Ostrowska, A.; Porebska, G. Assessment of the C/N ratio as an indicator of the decomposability of organic matter in forest soils. Ecol. Indic. 2015, 49, 104–109. [Google Scholar] [CrossRef]
- Güsewell, S.; Koerselman, W.; Verhoeven, J.T.A. Biomass N: P ratios as indicators of nutrient limitation for plant populations in wetlands. Ecol. Appl. 2003, 13, 372–384. [Google Scholar] [CrossRef]
- Bing, H.J.; Wu, Y.H.; Zhou, J.; Sun, H.Y.; Luo, J.; Dong, Y. Stoichiometric variation of carbon, nitrogen,and phosphorus in soils and its implication for nutrient limitation in alpine ecosystem of Eastern Tibetan Plateau. J. Soils Sediment. 2016, 16, 405–416. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.I. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Li, Y.; Wu, J.S.; Liu, S.L.; Shen, J.L.; Huang, D.; Su, Y.; Wei, W.; Syers, J.K. Is the C:N:P stoichiometry in soil and soil microbial biomass related to the landscape and land use in southern subtropical China? Glob. Biogeochem. Cycles 2012, 26, GB4002. [Google Scholar] [CrossRef]
- Batjes, N. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 2014, 65, 10–21. [Google Scholar] [CrossRef]
- Paul, E.A. Soil Microbiology, Ecology, and Biochemistry, 3rd ed.; Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2007. [Google Scholar]
- Penuelas, J.; Sardans, J.; Rivas-ubach, A.; Janssens, I.A. The human-induced imbalance between C, N and P in Earth’s life system. Glob. Chang. Biol. 2012, 18, 3–6. [Google Scholar] [CrossRef]
- Chen, F.; Wang, J.M.; Sun, B.G.; Chen, X.M.; Yang, Z.X.; Duan, Z.Y. Relationship between geographical distribution of Pinus yunnanensis and climate. J. For. Res. 2012, 25, 163–168. [Google Scholar]
- Yang, M.; Xie, Y.; Liu, J. Thirty Years of Eucalyptus Research in China (1981–2010); China Forestry Press: Beijing, China, 2011. [Google Scholar]
- Wu, J.; Liu, Z.; Huang, G.; Chen, D.; Zhang, W.; Shao, Y.; Wan, S.; Fu, S. Response of soil respiration and ecosystem carbon budget to vegetation removal in eucalyptus plantations with contrasting ages. Sci. Rep. 2014, 4, 6262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.T.; Dong, Y.R. Factors affecting species diversity of plant communities and the restoration process in the loess area of China. Ecol. Eng. 2010, 36, 345–350. [Google Scholar] [CrossRef]
- Luo, Y.Z.; Zeng, T.; Dong, W.; He, Y.L.; Xu, S.M. Research of ecological stoichiometry of plant community in alpine Kobresia meadow. Chin. J. Grassl. 2013, 35, 92–96. [Google Scholar]
- Killingbeck, K.T. Nutrients in senesced leaves: Keys to the search for potential reuse and reuse proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Van, H.L.M.; Toet, S.R.; Aerts, R. Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: Facts and solutions. Oikos 2003, 101, 664–669. [Google Scholar]
- Leonardus, V.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Fairfield, OH, USA, 2002. [Google Scholar]
- Tong, Z.L.; Chen, Q.B.; Wang, Y.X.; Xiong, H.Q.; Wu, J.X. Accumulation and distribution characteristics of nutrients in Pinus yunnanensis forests with different ages. J. Northwest A&F Univ. 2014, 42, 100–114. (In Chinese) [Google Scholar]
- Yang, Y.; Ran, F.; Wang, G.X.; Zhu, W.Z.; Yang, Y.; Zhou, P. Biomass model and carbon storage of Pinus yunnanensis on Tibet Plateau of China. Chin. J. Ecol. 2013, 32, 1674–1682. (In Chinese) [Google Scholar]
- Xu, Y.X.; Wang, Z.C.; Zhu, W.K.; Du, A.P. Stochiometric characteristics of C,N and P in leaf-litter-soil of different Eucalyptus in varieties. J. Northwest A&F Univ. 2017, 46, 1–8. (In Chinese) [Google Scholar]
- Yuan, Z.Y.; Chen, H.Y.H. Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob. Ecol. Biogeogr. 2009, 18, 11–18. [Google Scholar] [CrossRef]
- Gorokhova, E.; Kyle, M. Analysis of nucleic acids in daphnia: Development of methods and ontogenetic variations in RNA–DNA content. J. Plankton Res. 2002, 24, 511–522. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Contner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 6, 540–550. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Wright, I.J.; Niklas, K.J.; Hedin, L.; Elser, J.J. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes. Proc. R. Soc. B Biol. Sci. 2010, 277, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Brundrett, M.C.; Raven, J.A.; Hopper, S.D. Plant mineral nutrition in ancient landscapes: High plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant Soil 2010, 334, 11–31. [Google Scholar] [CrossRef]
- Fan, H.B.; Wu, J.P.; Liu, W.F.; Yuan, Y.H.; Hu, L.; Cai, Q.K. Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 2015, 392, 127–138. [Google Scholar] [CrossRef]
- Cao, Y.B.; Wang, B.T.; Wei, T.T.; Ma, H. Ecological stoichiometric characteristics and element reserves of three stands in a closed forest on the Chinese loess plateau. Environ. Monit. Assess. 2016, 188, 80. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.T.; Gu, Z.J.; Shao, H.B.; Zhou, F.; Peng, S.Y. N–P stoichiometry in soil and leaves of Pinus massoniana forest at different stand ages in the subtropical soil erosion area of China. Environ. Earth Sci. 2016, 75, 1091. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.J.; Yu, G.R.; Tao, B.; Wang, S.Q. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC. Environ. Sci. 2007, 28, 2665–2673. [Google Scholar]
- Yuan, Z.Y.; Chen, H.Y.H. Global trends in senesced-leaf nitrogen and phosphorus. Glob. Ecol. Biogeogr. 2009, 18, 532–542. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). For. Ecol. Manag. 2011, 262, 2024–2034. [Google Scholar] [CrossRef]
- He, M.Z.; Dijkstra, F.A.; Zhang, K.; Li, X.; Tan, H.; Gao, Y.; Li, G. Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Sci. Rep. 2014, 4, 6932. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.G.; Wang, G.G.; Wu, Q.T.; Cheng, X.; Yu, M.; Wang, W.; Yu, X. Patterns of leaf nitrogen and phosphorus stoichiometry among Quercus acutissima provenances across China. Ecol. Complex. 2014, 17, 32–39. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Pendall, E.; Morgan, J.A.; Blumenthal, D.M.; Carrillo, Y.; LeCain, D.R.; Follett, R.F.; Williams, D.G. Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland. New Phytol. 2012, 196, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Houlton, B.Z.; Wang, Y.P.; Vitousek, P.M.; Field, C.B. A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 2008, 454, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, S.; Trofymow, J.A.; Jackson, R.B.; Porporato, A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecol. Monogr. 2010, 80, 89–106. [Google Scholar] [CrossRef]
- Chen, Y.H.; Han, W.X.; Tang, L.Y.; Tang, Z.Y.; Fan, J.Y. Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography 2011, 36, 178–184. [Google Scholar] [CrossRef]
- Aponte, C.; Marañón, T.; García, L. Microbial C, N, and P in soils of Mediterranean oak forests: Influence of season, canopy cover and soil depth. Biogeochemistry 2010, 101, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.K.; Wang, S.L.; Fan, B.; Yu, X.J. Litter production, leaf litter decomposition and nutrient return in Cunninghamia lanceolata plantations in south China: Effect of planting conifers with broadleaved species. Plant Soil 2007, 297, 201–211. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Carbon and nitrogen dynamics of soil organic fractions from cultivated grassland soils. Soil Sci. Soc. Am. J. 1992, 58, 123–130. [Google Scholar] [CrossRef]
- Aerts, R.; Caluwe, H.D.; Beltman, B. Is the relation between nutrient supply and biodiversity co-limited by the type of nutrient limitation? Oikos 2003, 101, 489–498. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N: P ratio: A new tool to detect the Nature of Nutrient Limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, H.; Zhang, T.; Zhao, X.; Drake, S. Community succession along a chronosequence of vegetation restoration on sand dunes in Horqin Sandy Land. J. Arid Environ. 2005, 62, 555–566. [Google Scholar] [CrossRef]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data. Biogeochemistry 2010, 98, 139–151. [Google Scholar] [CrossRef]
- Izquierdo, J.; Houlton, B.; Huysen, T. Evidence for progressive phosphorus limitation over long-term ecosystem development: Examination of a biogeochemical paradigm. Plant Soil 2013, 367, 135–147. [Google Scholar] [CrossRef]
- Chiwa, M.; Ikezaki, S.; Katayama, A.; Enoki, T. Topographic Influence on Plant Nitrogen and Phosphorus Stoichiometry in a Temperate Forested Watershed. Water Air Soil Pollut. 2013, 227, 6. [Google Scholar] [CrossRef]
- Solomon, D.; Lehmann, J.; Mano, T.; Fritzsche, F.; Zech, W. Phosphorus forms and dynamics as influenced by land use changes in the sub-humid Ethiopian highland. Geoderma 2002, 105, 21–48. [Google Scholar] [CrossRef]
- Yang, J.C.; Huang, J.H.; Pan, Q.M.; Tang, J.W.; Han, X.G. Soil phosphorus dynamics as influenced by land use changes in humid tropical, Southwest China. Pedosphere 2005, 15, 24–32. [Google Scholar]
- Zhao, X.L.; He, X.D.; Xue, P.P.; Zhang, N.; We, W.; Li, R.; Ci, H.C.; Xu, J.J.; Gao, Y.B.; Zhao, H.L. Effects of soil stoichiometry of the CaCO3/available phosphorus ratio on plant density in Artemisia ordosica communities. Chin. Sci. Bull. 2012, 57, 492–499. [Google Scholar] [CrossRef]
- Escudero, A.; Arco, J.M.; Sanz, I.C.; Ayala, J. Effects of leaf longevity and retranslocation efficiency on the retention time of nutrients in the leaf biomass of different wood species. Oecologia 1992, 90, 80–87. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Tree Species | Height (m) | DBH (cm) | Crown Diameter (m) | Dominant Shrub Species | Dominant Herb Species |
|---|---|---|---|---|---|
| Eucalyptus maideni | 7.06 | 6.42 | 2.25 × 2.33 | Campylotropis polyantha, Dodonaea viscosa (L.) Jacq., Osteomeles anthyllidifolia (Sm.) Lindl. | Arthraxon hispidus (Thunb.) Makino, Ainsliaea henryi Diels, Rubia cordifolia L., Juncus effusus, Arundinella anomala Steud. |
| Pinus yunnanensis | 3.38 | 3.66 | 3.32 × 3.26 | Carissa spinarum, Osteomeles anthyllidifolia (Sm.) Lindl, Myrsine africana L., Dodonaea viscosa (L.) Jacq. | Arundinella anomala, Gramineae, Barleria cristata L., Elsholtzia ciliata (Thumb.) Hyland. |
| Soil Layer | 0–10 cm | 10–20 cm | 20–30 cm | |||
|---|---|---|---|---|---|---|
| Vegetation Type | EM | PY | EM | PY | EM | PY |
| pH | 5.467 ± 0.196a | 5.990 ± 0.115a | 5.827 ± 0.096a | 5.997 ± 0.084a | 5.637 ± 0.318a | 5.990 ± 0.066a |
| SWC (%) | 33.976 ± 1.993a | 22.361 ± 1.983b | 30.494 ± 4.419a | 22.805 ± 0.955b | 30.816 ± 4.710a | 23.719 ± 0.940b |
| BD (g cm−3) | 1.026 ± 0.033a | 1.077 ± 0.048a | 1.120 ± 0.059a | 1.255 ± 0.025a | 1.172 ± 0.059a | 1.284 ± 0.078a |
| CP (%) | 50.274 ± 0.013a | 48.774 ± 0.035a | 44.402 ± 0.010a | 47.757 ± 0.003a | 44.688 ± 0.023a | 46.848 ± 0.021a |
| AP (mg kg−1) | 1.133 ± 0.061a | 1.013 ± 0.821a | 0.973 ± 0.122a | 1.000 ± 0.734a | 1.627 ± 1.103a | 0.667 ± 0.300b |
| AK (mg kg−1) | 73.333 ± 17.638b | 126.667 ± 32.145a | 70.000 ± 0.000b | 90.000 ± 17.321a | 60.000 ± 20.000b | 76.667 ± 11.548a |
| NO3−-N (mg kg−1) | 11.127 ± 0.562a | 12.907 ± 0.406a | 10.533 ± 0.284a | 12.480 ± 0.555a | 10.053 ± 0.511a | 12.147 ± 0.081a |
| NH4+-N (mg kg−1) | 4.437 ± 1.594a | 5.735 ± 4.048a | 3.567 ± 0.424a | 3.325 ± 0.058a | 2.625 ± 0.322a | 2.663 ± 0.411a |
| Variable | The Characteristics of C:N:P Stoichiometry | ||
|---|---|---|---|
| EM | PY | ||
| Plant organs | Leaves | 227:14:1 | 281:19:1 |
| Branches | 226:16:1 | 273:7:1 | |
| Stems | 112:12:1 | 235:7:1 | |
| Roots | 257:16:1 | 211:10:1 | |
| Average | 206:14:1 | 250:11:1 | |
| Soil depth (cm) | 0–10 | 41:16:1 | 57:8:1 |
| 10–20 | 28:12:1 | 47:9:1 | |
| 20–30 | 25:6:1 | 40:8:1 | |
| Average | 31:11:1 | 48:8:1 | |
| Vegetation Type | Equations | y | F | R2 | P |
|---|---|---|---|---|---|
| EM | y = 0.015x1 − 0.389x2 − 2.587x3 + 0.874x4 − 0.017x5 + 2.806x6 + 0.734x7 − 23.015 | RCN | 35.06 | 0.996 | 0.1293 |
| PY | y = −0.740x1 − 2.340x2 + 43.140x3 + 9.193x4 + 0.334x5 − 11.207x6 − 1.046x7 + 117.997 | RCN | 8.61 | 0.984 | 0.2567 |
| EM | y = 1.140x1 − 0.375x2 − 41.786x3 − 3.365x4 + 0.001x5 + 1.869x6 + 1.277x7 + 20.026 | RCP | 798.87 | 0.999 | 0.0272 |
| PY | y = 1.704x1 + 0.759x2 − 75.863x3 + 3.296x4 + 0.141x5 − 8.381x6 − 0.796x7 + 131.985 | RCP | 3848.39 | 0.999 | 0.0124 |
| EM | y = −0.109x1 + 1.121x2 − 9.592x3 − 1.810x4 − 0.002x5 + 1.342x6 − 0.343x7 + 0.593 | RNP | 520.05 | 0.999 | 0.0338 |
| PY | y = 0.059x1 + 1.665x2 − 16.395x3 − 1.037x4 − 0.046x5 + 2.316x6 + 0.206x7 − 16.212 | RNP | 583.43 | 0.999 | 0.0319 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, D.; Wang, G.; Li, G.; Sun, Y.; Liu, Y.; Zhou, J. Ecological Stoichiometric Characteristics of Two Typical Plantations in the Karst Ecosystem of Southwestern China. Forests 2018, 9, 56. https://doi.org/10.3390/f9020056
Pang D, Wang G, Li G, Sun Y, Liu Y, Zhou J. Ecological Stoichiometric Characteristics of Two Typical Plantations in the Karst Ecosystem of Southwestern China. Forests. 2018; 9(2):56. https://doi.org/10.3390/f9020056
Chicago/Turabian StylePang, Danbo, Genzhu Wang, Guijing Li, Yonglei Sun, Yuguo Liu, and Jinxing Zhou. 2018. "Ecological Stoichiometric Characteristics of Two Typical Plantations in the Karst Ecosystem of Southwestern China" Forests 9, no. 2: 56. https://doi.org/10.3390/f9020056
APA StylePang, D., Wang, G., Li, G., Sun, Y., Liu, Y., & Zhou, J. (2018). Ecological Stoichiometric Characteristics of Two Typical Plantations in the Karst Ecosystem of Southwestern China. Forests, 9(2), 56. https://doi.org/10.3390/f9020056