Permafrost Regime Affects the Nutritional Status and Productivity of Larches in Central Siberia



 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling and Analyses
2.2.1. Stand Inventory and Tree Biomass Measurements
2.2.2. Soils
2.2.3. Larch Needles
2.3. Statistical Analyses
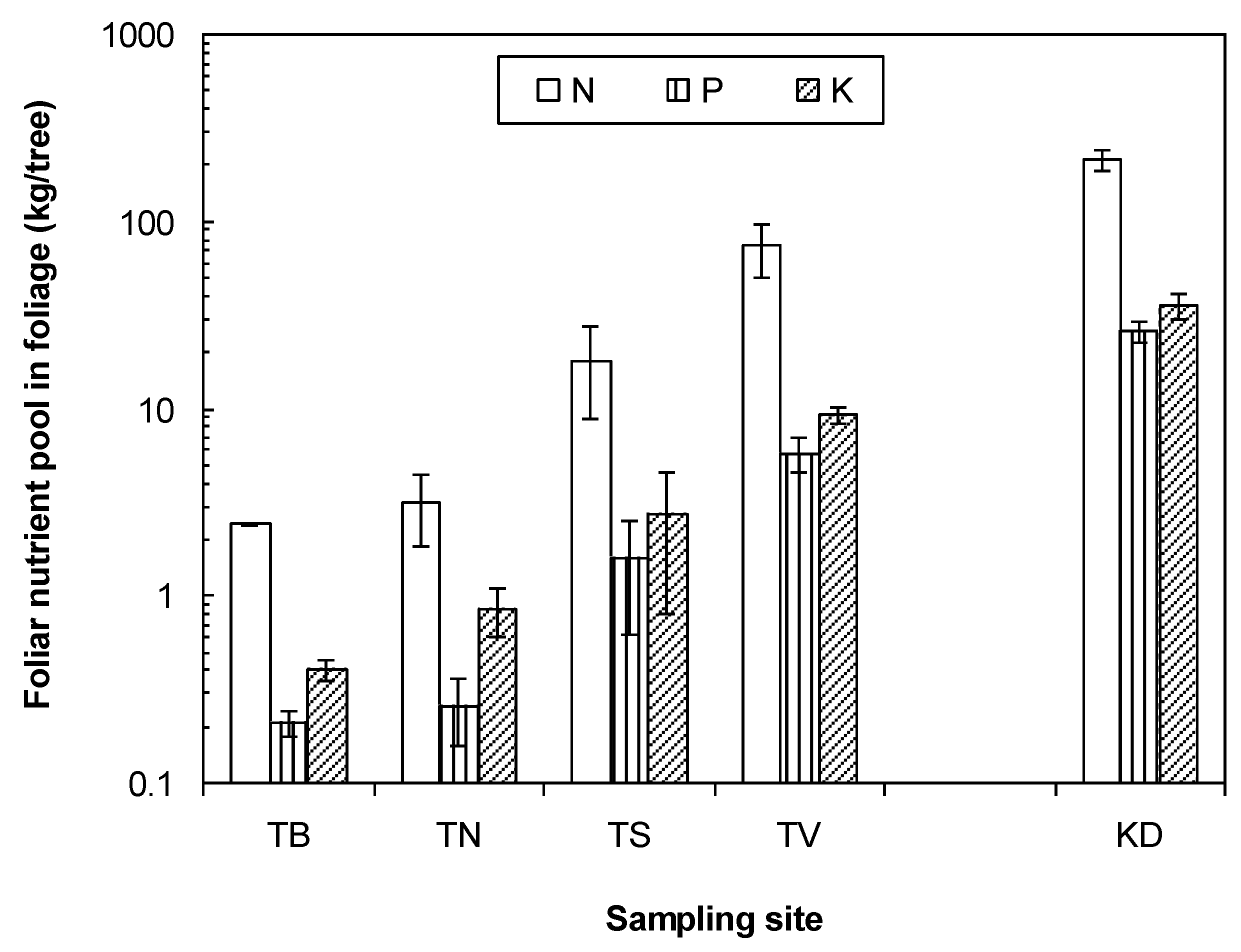
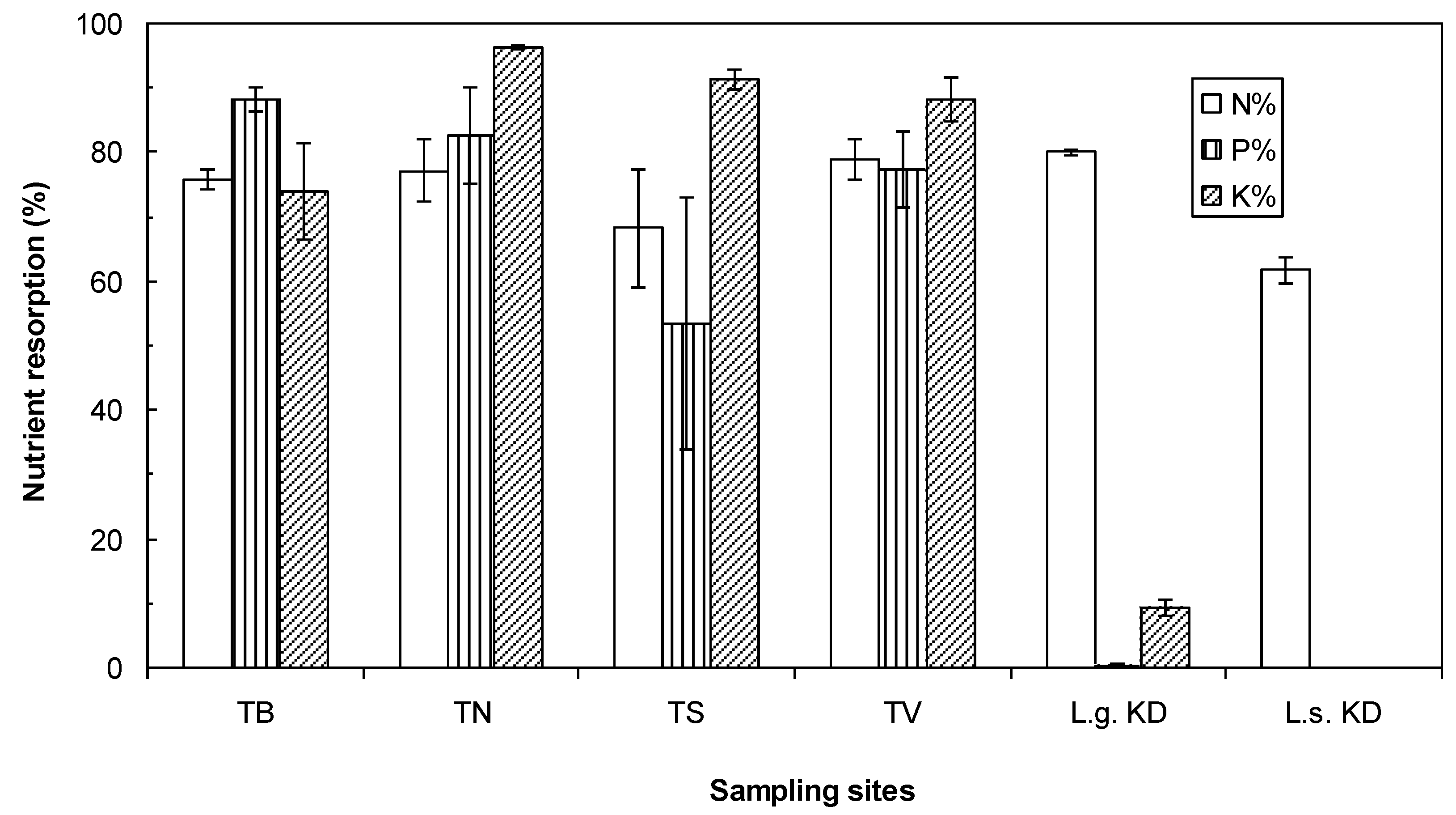
3. Results
3.1. Tree and Soil Parameters
3.2. Seasonal Dynamics of Foliar Element Concentrations and Stable Isotope Composition
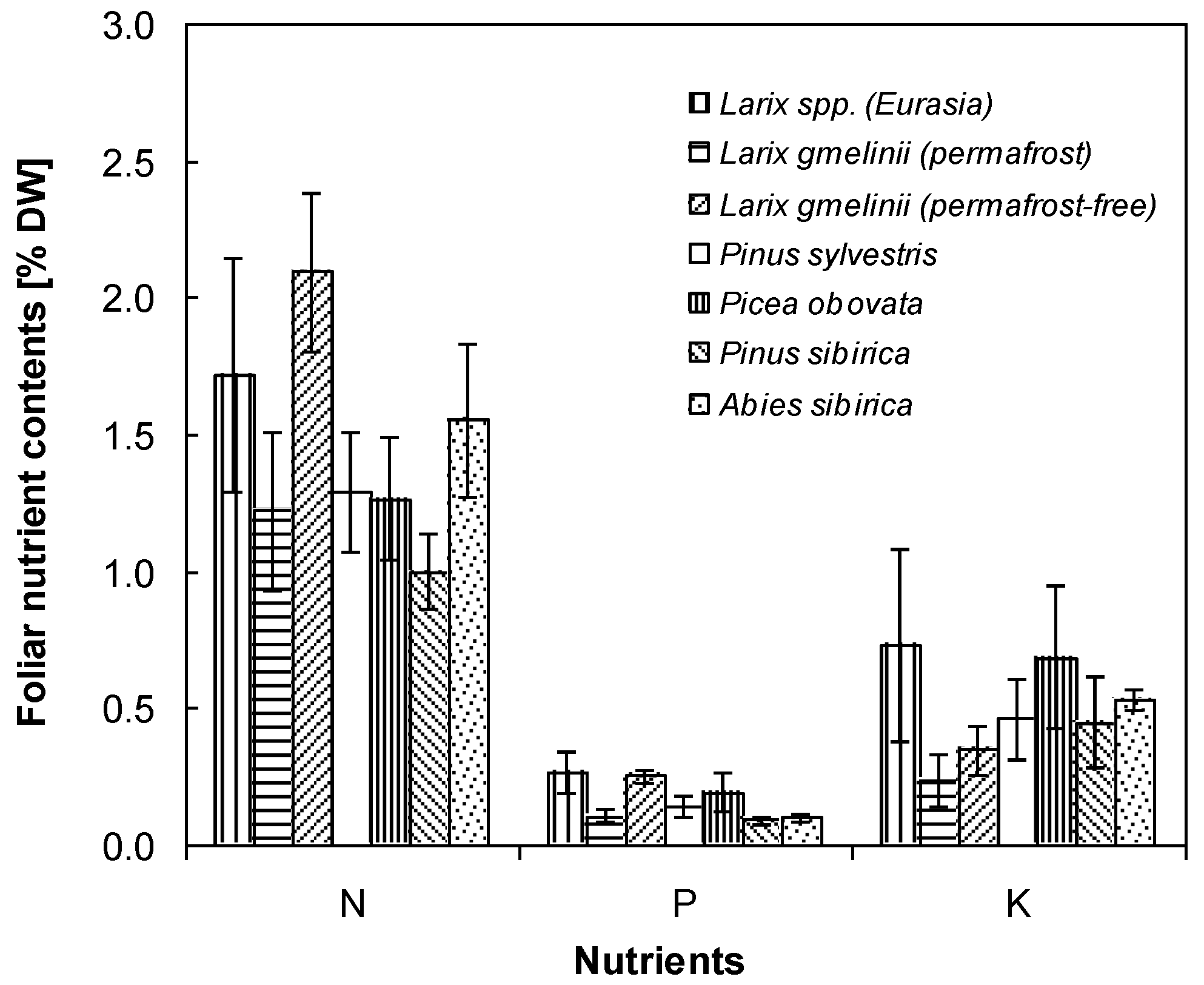
4. Discussion
4.1. Nutrient Status of Larch Improves with Active Layer Depth
4.2. 15N Enrichment of Needles on Warmer Soils
4.3. Strong Seasonal Dynamics in Nutrient Concentrations and 13C
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Goodale, C.L.; Apps, M.J.; Birdsey, R.A. Forest carbon sinks in the Northern Hemisphere. Ecol. Appl. 2002, 12, 891–899. [Google Scholar] [CrossRef]
- Gower, S.T.; Richards, J.H. Larches: Deciduous conifers in an evergreen world. BioScience 1990, 40, 818–826. [Google Scholar] [CrossRef]
- Vedrova, E.F.; Pleshikov, F.I.; Kaplunov, V.Y. Net ecosystem production of boreal Larch ecosystems on the Yenisei transect. Mitig. Adapt. Strat. Glob. Chang. 2006, 11, 173–190. [Google Scholar] [CrossRef]
- Abaimov, A.P. Geographical Distribution and Genetics of Siberian Larch Species. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Kajimoto, T., Zyryanova, O.A., Matsuura, Y., Wein, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 41–58. ISBN 978-1-4020-9692-1. [Google Scholar]
- Kloeppel, B.D.; Gower, S.T.; Triechel, I.W.; Kharuk, S. Foliar carbon isotope discrimination in Larix species and sympatric evergreen conifers: A global comparison. Oecologia 1998, 114, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Vygodskaya, N.N.; Milyukova, I.; Varlagin, A.; Tatarinov, F.; Sogachev, A.; Kobak, K.I.; Desyatkin, R.; Bauer, G.; Hollinger, D.Y.; Kelliher, F.M.; et al. Leaf conductance and CO2 assimilation of Larix gmelinii growing in an eastern Siberian boreal forest. Tree Physiol. 1997, 17, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, A.; Yanagisawa, N.; Naito, D.; Fujita, N.; Maximov, T.C. Importance of permafrost as a source of water for plants in east Siberian taiga. Ecol. Res. 2002, 17, 493–503. [Google Scholar] [CrossRef]
- Alexeev, V.A.; Birdsey, R.A.; Stakanov, V.D.; Korotkov, I. Carbon in vegetation of Russian forests: Methods to estimate storage and geographical distribution. Water Air Soil Pollut. 1995, 82, 271–282. [Google Scholar] [CrossRef]
- Li, S.-G.; Tsujimura, M.; Sugimoto, A.; Davaa, G.; Oyunbaatar, D.; Sugita, M. Temporal variation of δ13C of larch leaves from a montane boreal forest in Mongolia. Trees 2007, 21, 479–490. [Google Scholar] [CrossRef]
- Dolman, A.J.; Maximov, T.C.; Moors, E.J.; Maximov, A.P.; Elbers, J.A.; Kononov, A.V.; Waterloo, M.J.; van der Molen, M.K. Net ecosystem exchange of carbon dioxide and water of far eastern Siberian Larch (Larix cajanderii) on permafrost. Biogeosciences 2004, 1, 133–146. [Google Scholar] [CrossRef]
- Nakai, Y.; Matsuura, Y.; Kajimoto, T.; Abaimov, A.P.; Yamamoto, S.; Zyryanova, O.A. Eddy covariance CO2 flux above a Gmelin larch forest on continuous permafrost in Central Siberia during a growing season. Theor. Appl. Climatol. 2008, 9, 133–147. [Google Scholar] [CrossRef]
- Sidorova, O.V.; Siegwolf, R.; Saurer, M.; Shashkin, A.V.; Knorre, A.A.; Prokushkin, A.S.; Vaganov, E.A.; Kirdyanov, A.V. Do centennial tree-ring and stable isotope trends of Larix gmelinii (Rupr.) indicate increasing water shortage in the Siberian north? Oecologia 2009, 161, 25–835. [Google Scholar] [CrossRef] [PubMed]
- Girs, G.I. Accumulation of Nitrogen, Phosphorus and Potassium by Soil-Forming Species of Russia; V.N. Sukachev Institute of Forest SB RAS: Krasnoyarsk, Russia, 1996. (In Russian) [Google Scholar]
- Schulze, E.D.; Schulze, W.; Kelliher, F.M.; Vygodskaya, N.N.; Ziegler, W.; Kobak, K.I.; Koch, H.; Arneth, A.; Kusnetsova, W.A.; Sogachev, A.; et al. Aboveground biomass and nitrogen nutrition in a chronosequence of pristine Gmelin’s Larix stands in Eastern Siberia. Can. J. For. Res. 1995, 25, 943–960. [Google Scholar] [CrossRef]
- Kajimoto, T.; Matsuura, Y.; Osawa, A.; Abaimov, A.P. Size-mass allometry and biomass allocation of two larch species growing on the continuous permafrost region in Siberia. For. Ecol. Manag. 2006, 22, 314–325. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Aerts, R.; Chapin, F.S., III. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar] [CrossRef]
- Lambers, H.; Shaver, G.; Raven, J.A. N- and P-acquisition change as soils age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.P.; Law, R.M.; Pak, B. A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosciences 2009, 6, 9891–9944. [Google Scholar] [CrossRef]
- Martinelli, L.A.; Piccolo, M.C.; Townsend, A.R.; Vitousek, P.M.; Cuevas, E.; McDowell, W.; Robertson, G.P.; Santos, O.C.; Treseder, K. Nitrogen stable isotopic composition of leaves and soil: Tropical versus temperate forests. Biogeochemistry 1999, 46, 45–65. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.Y.; Makoto, K.; Choi, D.S.; Quoreshi, A.M.; Koike, T. The role of ectomycorrhiza in boreal forest ecosystem. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Kajimoto, T., Zyryanova, O.A., Matsuura, Y., Wein, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 413–426. ISBN 978-1-4020-9692-1. [Google Scholar]
- Michelsen, A.; Quarmby, C.; Sleep, D.; Jonasson, S. Vascular plant 15N natural abundance in heath and forest tundra ecosystems is closely correlated with presence and type of mycorrhizal fungi in roots. Oecologia 1998, 115, 406–418. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, E.A.; Ouimette, A.P. Controls of nitrogen isotope patterns in soil profiles. Biogeochemistry 2009, 95, 355–371. [Google Scholar] [CrossRef]
- Aerts, R. Nutrient resorption from senescing leaves of perennials: Are there general patterns? J. Ecol. 1996, 4, 597–608. [Google Scholar] [CrossRef]
- Killingbeck, K.T. Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency. Ecology 1996, 77, 1716–1727. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, Y.; Hirobe, M. Soil carbon and nitrogen, and characteristics of soil active layer in Siberian permafrost region. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Kajimoto, T., Zyryanova, O.A., Matsuura, Y., Wein, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 149–164. ISBN 978-1-4020-9692-1. [Google Scholar]
- Eckstein, R.L.; Karlsson, P.S.; Weih, M. Leaf life span and nutrient resorption as determinants of plant nutrient conservation in temperate-arctic regions. New Phytol. 1999, 143, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Prokushkin, A.S.; Tokareva, I.V.; Prokushkin, S.G.; Abaimov, A.P.; Guggenberger, G. Fluxes of dissolved organic matter in larch forests in the Cryolithozone of Central Siberia. Rus. J. Ecol. 2008, 39, 151–159. [Google Scholar] [CrossRef]
- Wild, B.; Schnecker, J.; Knoltsch, A.; Takriti, M.; Mooshammer, M.; Gentsch, N.; Mikutta, R.; Eloy Alves, R.J.; Gittel, A.; Lashchinskiy, N.; et al. Microbial nitrogen dynamics in organic and mineral soil horizons along a latitudinal transect in western Siberia. Glob. Biogeochem. Cycles 2015, 29, 567–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viers, J.; Prokushkin, A.S.; Pokrovsky, O.S.; Auda, Y.; Kirdyanov, A.V.; Beaulieu, E.; Zouiten, C.; Oliva, P.; Dupre, B. Seasonal and spatial variability of elemental concentrations in boreal forest larch foliage of Central Siberia on continuous permafrost. Biogeochemistry 2013, 113, 435–449. [Google Scholar] [CrossRef]
- Larjavaara, M.; Berninger, F.; Palviainen, M.; Prokushkin, A.; Wallenius, T. Post-fire carbon and nitrogen accumulation and succession in Central Siberia. Sci. Rep. 2017, 12776. [Google Scholar] [CrossRef] [PubMed]
- Wood. Method of Selection of Model Trees and Logs for Determination of Physical and Mechanical Properties of Wood Plantations. State Standard of the USSR (ГОСТ 16483.6-80). 1981. Available online: http://docs.cntd.ru/document/1200014947 (accessed on 4 April 2014).
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology. Application in Environmental Sciences; Springer: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Farquhar, G.D.; Ehleringer, R.; Hubic, K.T. Carbon isotope discrimination and photosynthesis. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Kajimoto, T.; Matsuura, Y.; Sofronov, M.A.; Volokitina, A.V.; Mori, S.; Osawa, A.; Abaimov, A.P. Above- and belowground biomass and net primary productivity of a Larix gmelinii stand near Tura, Central Siberia. Tree Physiol. 1999, 19, 815–822. [Google Scholar] [CrossRef] [PubMed]
- McGroddy, M.E.; Daufresne, T.; Hedin, L.O. Scaling of C:N:P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios. Ecology 2004, 85, 2390–2401. [Google Scholar] [CrossRef]
- Prokushkin, A.S.; Knorre, A.A.; Kirdyanov, A.V.; Schulze, E.D. Productivity of mosses and organic matter accumulation in the litter of sphagnum larch forest in the permafrost zone. Rus. J. Ecol. 2006, 37, 225–232. [Google Scholar] [CrossRef]
- Tokuchi, N.; Hirobe, M.; Kondo, K.; Arai, H.; Hobara, S.; Fukushima, K.; Matssura, Y. Soil nitrogen dynamics in larch ecosystems. In Permafrost Ecosystems: Siberian Larch Forests; Osawa, A., Kajimoto, T., Zyryanova, O.A., Matsuura, Y., Wein, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 229–244. ISBN 978-1-4020-9692-1. [Google Scholar]
- Shugaley, L.S.; Vedrova, E.F. Nitrogen pool in northern taiga larch forests of Central Siberia. Biol. Bull. 2004, 31, 200–208. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Högberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen dynamics. New Phytol. 2012, 196, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Mayor, J.R.; Schuur, E.A.G.; Mack, M.C.; Hollingsworth, T.N.; Baath, E. Nitrogen isotope patterns in Alaskan black spruce reflect organic nitrogen sources and the activity of ectomycorrhizal fungi. Ecosystems 2012, 15, 819–831. [Google Scholar] [CrossRef]
- Högberg, M.N.; Briones, M.J.I.; Keel, S.G.; Metcalfe, D.B.; Campbell, C.; Midwood, A.J.; Thornton, B.; Hurry, V.; Linder, S.; Nasholm, T.; et al. Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest. New Phytol. 2010, 187, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Dawes, M.A.; Schleppi, P.; Hagedorn, F. The fate of nitrogen inputs in a warmer alpine treeline ecosystem: A 15N labelling study. J. Ecol. 2017, 105, 1723–1737. [Google Scholar] [CrossRef]
- Wurzburger, N.; Hartshorn, A.S.; Hendrick, R.L. Ectomycorrhizal fungal community structure across a bog-forest ecotone in southeastern Alaska. Mycorrhiza 2004, 14, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Oleksyn, J.; Wright, I.J. Leaf phosphorus influences the photosynthesis–nitrogen relation. A cross-biome analysis of 314 species. Oecologia 2009, 160, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Woods, H.A.; Makino, W.; Cotner, J.B.; Hobbie, S.; Harrison, J.F.; Acharya, K.; Elser, J.J. Temperature and the chemical composition of poikilothermic organisms. Funct. Ecol. 2003, 17, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Prokushkin, S.G. Mineral Nutrition of Pine; Nauka: Novosibirsk, Russia, 1982. (In Russian) [Google Scholar]
- Lukina, N.V. Seasonal variation in chemical composition of Pinus sylvestris L. needles in the Kola Peninsula. Lesovedenie 1996, 1, 41–53. (In Russian) [Google Scholar]
- Glynn, C.; Herms, D.A.; Orians, C.M.; Hansen, R.C.; Larsson, S. Testing the growth–differentiation balance hypothesis: Dynamic responses of willows to nutrient availability. New Phytol. 2007, 176, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Bazilevich, N.I.; Titlyanova, A.A. Biotic Turnover on Five Continents. Element Exchange Processes in Terrestrial Natural Ecosystems; Publishing House SB RAS: Novosibirsk, Russia, 2008; ISBN 978-5-7692-0941-3. (In Russian) [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Dominant | Study Site | Latitude/Longitude | Stand Density, Tree/ha | Age, Years | Mean Tree Ring Width, mm | DBH, cm | Tree Height, m | Aboveground Biomass, Mg/ha | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Trunk | Needle | Total | ||||||||
| Tura: continuous permafrost terrain | ||||||||||
| L. gmelinii | Sphagnum bog (TB) | 64°19′30″ N 100°15′53″ E | 2480 | 86 | 0.11 ± 0.05 | 4.0 | 4.1 | 5.6 | 0.3 | 7.0 |
| L. gmelinii | North-facing slope (TN) | 64°19′21″ N 100°14′53″ E | 4400 | 99 | 0.11 ± 0.03 | 5.8 | 7.7 | 26.9 | 0.5 | 28.2 |
| L. gmelinii | South-facing slope (TS) | 64°19′32″ N 100°15′32″ E | 2700 | 91 | 0.17 ± 0.05 | 9.3 | 11.3 | 52.3 | 1.6 | 56.6 |
| L. gmelinii | Riparian zone (TV) | 64°19′29″ N 100°15′07″ E | 1100 | 156 | 0.51 ± 0.20 | 15.0 | 12.9 | 55.9 | 7.0 | 79.0 |
| Krasnoyarsk: permafrost-free terrain | ||||||||||
| L. gmelinii | Common garden (L.g. KD) | 55°59′ N 92°45′ E | 760 | 32 | 3.55 ± 2.56 | 17.5 | 14.8 | 38.6 | 7.7 | 70.5 |
| L. sibirica | Common garden (L.s. KD) | 55°59′ N 92°45′ E | 800 | 32 | 3.58 ± 2.50 | 24.0 | 16.3 | 42.0 | 6.2 | 76.3 |
| Study Site ID | Soil Type * | Organic Layer | Mineral Soil | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OC | δ13C | N | δ15N | OC | δ13C @ | N | δ15N @ | Active Layer Depth, cm ^ | T5 cm & | Rooting Depth ** | |||||
| kg/m2 | ‰ | g/m2 | ‰ | kg/m2 | ‰ | g/m2 | ‰ | June | July | August | Sept | °C | cm | ||
| Tura: continuous permafrost terrain | |||||||||||||||
| TN | Typic Aquiturbel | 2.1 | −28.35 | 54 | −0.20 | 6.6 | −27.12 | 307 | 3.24 | 1 ± 1 | 25 ± 12 | 36 ± 11 | 42 | 2.8 | 12 |
| TS | Typic Haplocryept | 0.8 | −29.04 | 22 | −1.14 | 5.4 | −27.47 | 309 | 3.43 | 18 ± 8 | 86 ± 15 | 112 ± 26 | 123 | 6.6 | 64 |
| TB | Sphagnic Fibristel | 6.4 | −28.64 | 96 | 0.19 | 5.5 | −28.02 | 425 | 1.58 | 0 | 1 ± 2 | 15 ± 8 | 20 | 0.1 | 0 |
| TV | Lithic Aquorthel | 1.8 | −29.67 | 60 | −0.06 | 7.3 | −28.21 | 365 | 2.31 | 5 ± 2 | 52 ± 16 | 63 ± 19 | 88 | 4.1 | 45 |
| Krasnoyarsk: permafrost-free terrain | |||||||||||||||
| L.g. KD | Typic Haplocryalf | 0.4 | - | 16 | - | 2.9 | - | 251 | - | - | - | - | - | 9.2 | 70 |
| L.s. KD | Typic Haplocryalf | 0.3 | - | 17 | - | 3.2 | - | 342 | - | - | - | - | - | 8.9 | 62 |
| Permafrost-Affected Plots | Permafrost-Free Plot | |||||
|---|---|---|---|---|---|---|
| Date of Sampling | TB | TN | TS | TV | Date of Sampling | L.g. KD |
| 07.06.2006 | 144:6.8:1 | 77:4.7:1 | 78:5.4:1 | 71:4.8:1 | 31.05.2006 | 209:11.1:1 |
| 18.07.2006 | 440:11.7:1 | 421:10.5:1 | 327:10.3:1 | 293:13.4:1 | 20.07.2006 | 190:8.8:1 |
| 23.08.2006 | 557:11.9:1 | 493:12.1:1 | 451:12.0:1 | 335:11.3:1 | 25.08.2006 | 190:8.3:1 |
| 12.09.2006 | 4566:24.8:1 | 3227:18.0:1 | 1020:8.4:1 | 1236:8.9:1 | 20.10.2006 | 194:1.7:1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prokushkin, A.S.; Hagedorn, F.; Pokrovsky, O.S.; Viers, J.; Kirdyanov, A.V.; Masyagina, O.V.; Prokushkina, M.P.; McDowell, W.H. Permafrost Regime Affects the Nutritional Status and Productivity of Larches in Central Siberia. Forests 2018, 9, 314. https://doi.org/10.3390/f9060314
Prokushkin AS, Hagedorn F, Pokrovsky OS, Viers J, Kirdyanov AV, Masyagina OV, Prokushkina MP, McDowell WH. Permafrost Regime Affects the Nutritional Status and Productivity of Larches in Central Siberia. Forests. 2018; 9(6):314. https://doi.org/10.3390/f9060314
Chicago/Turabian StyleProkushkin, Anatoly S., Frank Hagedorn, Oleg S. Pokrovsky, Jerome Viers, Alexander V. Kirdyanov, Oxana V. Masyagina, Marina P. Prokushkina, and William H. McDowell. 2018. "Permafrost Regime Affects the Nutritional Status and Productivity of Larches in Central Siberia" Forests 9, no. 6: 314. https://doi.org/10.3390/f9060314
APA StyleProkushkin, A. S., Hagedorn, F., Pokrovsky, O. S., Viers, J., Kirdyanov, A. V., Masyagina, O. V., Prokushkina, M. P., & McDowell, W. H. (2018). Permafrost Regime Affects the Nutritional Status and Productivity of Larches in Central Siberia. Forests, 9(6), 314. https://doi.org/10.3390/f9060314