Compound-Specific Carbon Isotopes and Concentrations of Carbohydrates and Organic Acids as Indicators of Tree Decline in Mountain Pine

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Climate Data
2.3. Selection of Trees and Sampling of Needles
2.4. Extraction and Purification of Sugars and Organic Acids
2.5. Compound-Specific δ13C Isotope Analysis
2.6. Statistical Analysis
3. Results and Discussion
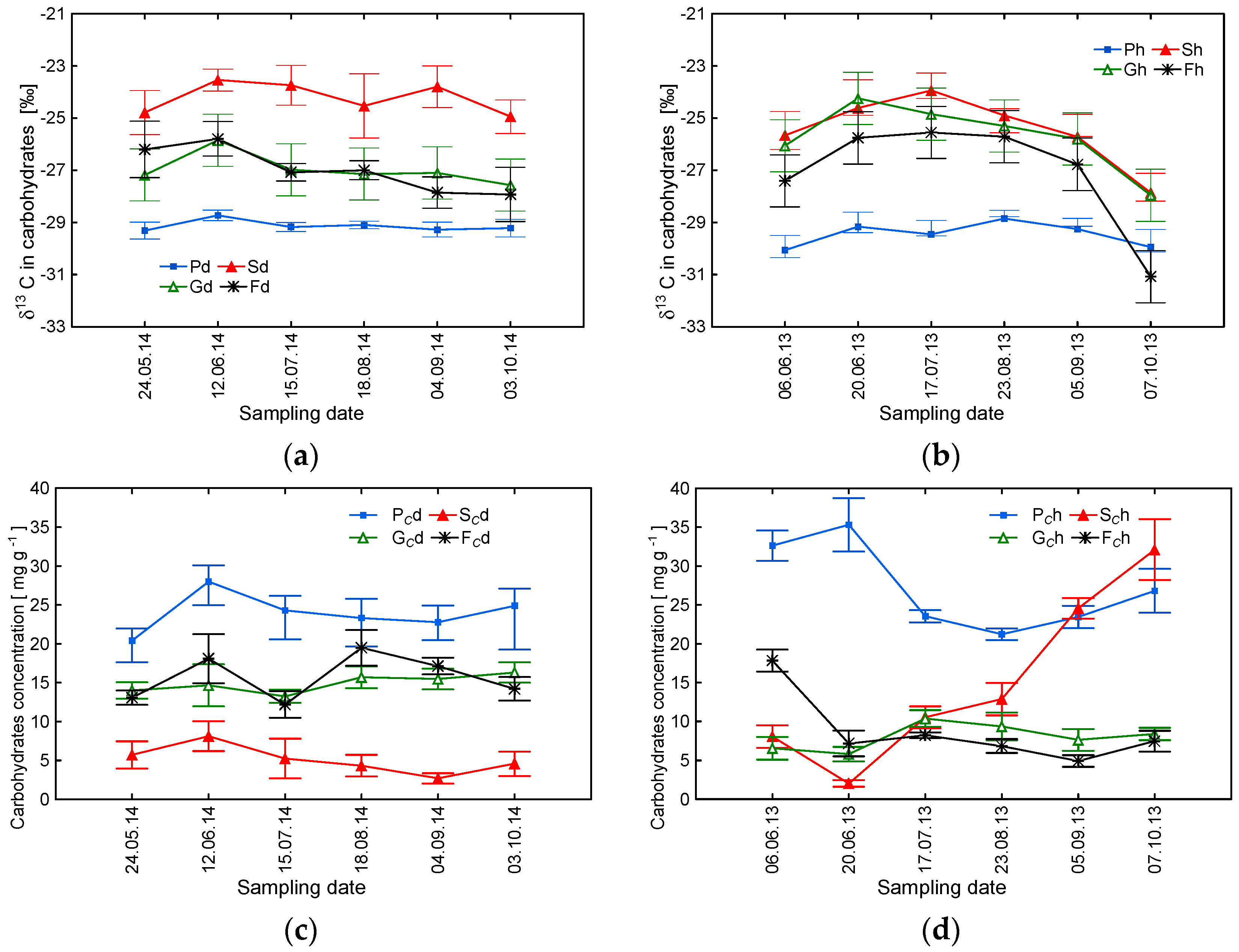
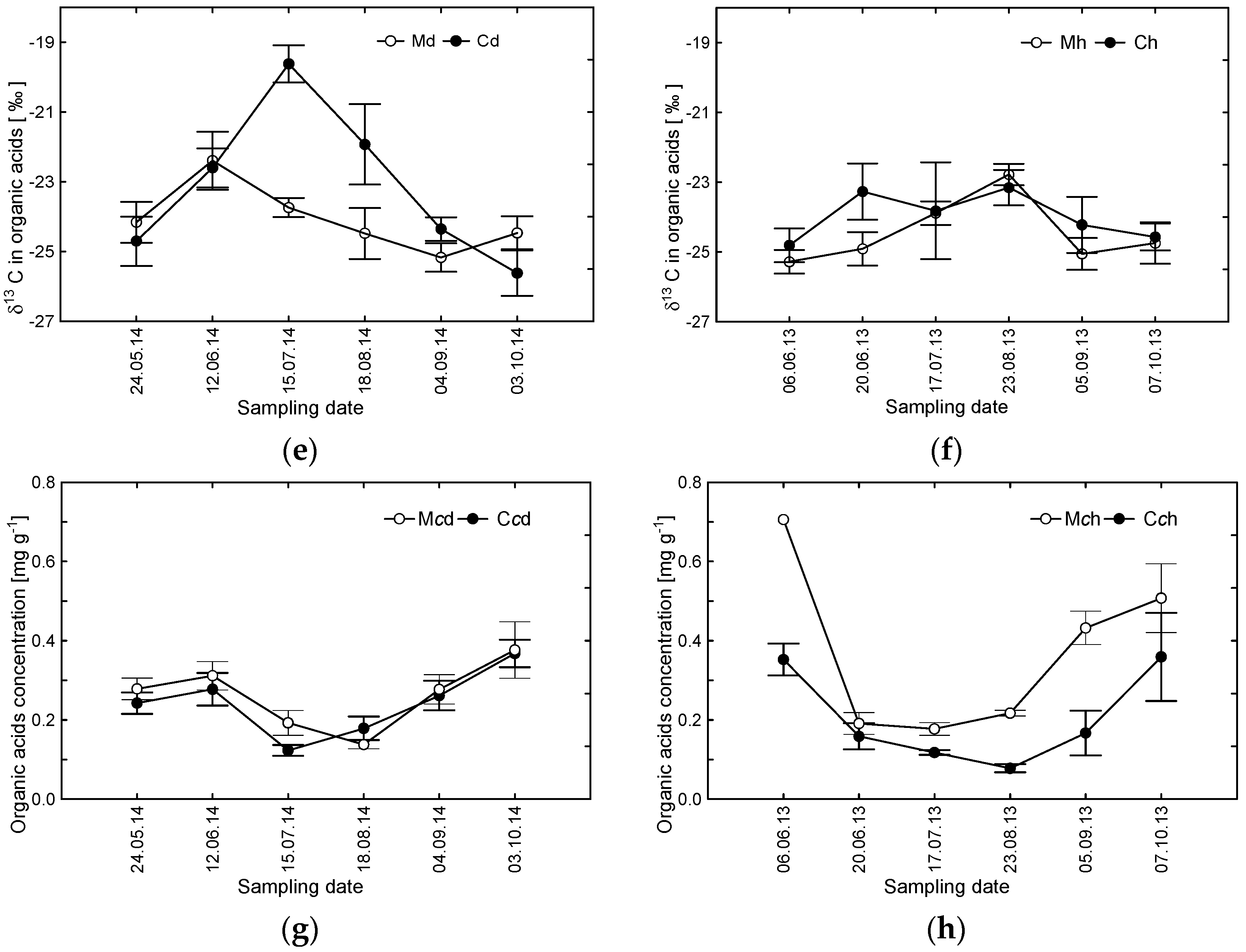
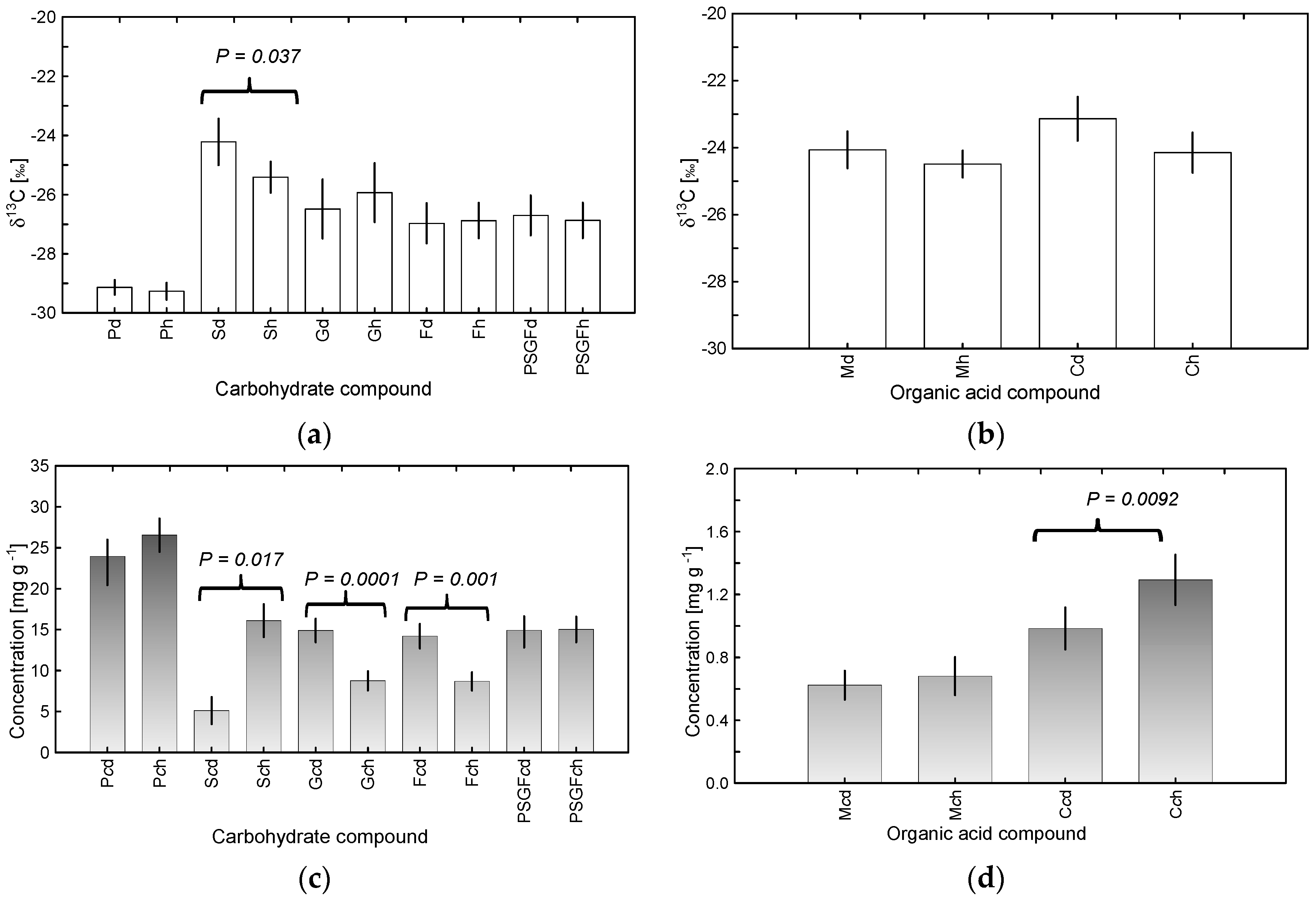
3.1. δ13C Values and Concentrations of Carbohydrates and Organic Acids as a Potential Indicator of Tree Decline
3.2. Averaged Seasonal Patterns
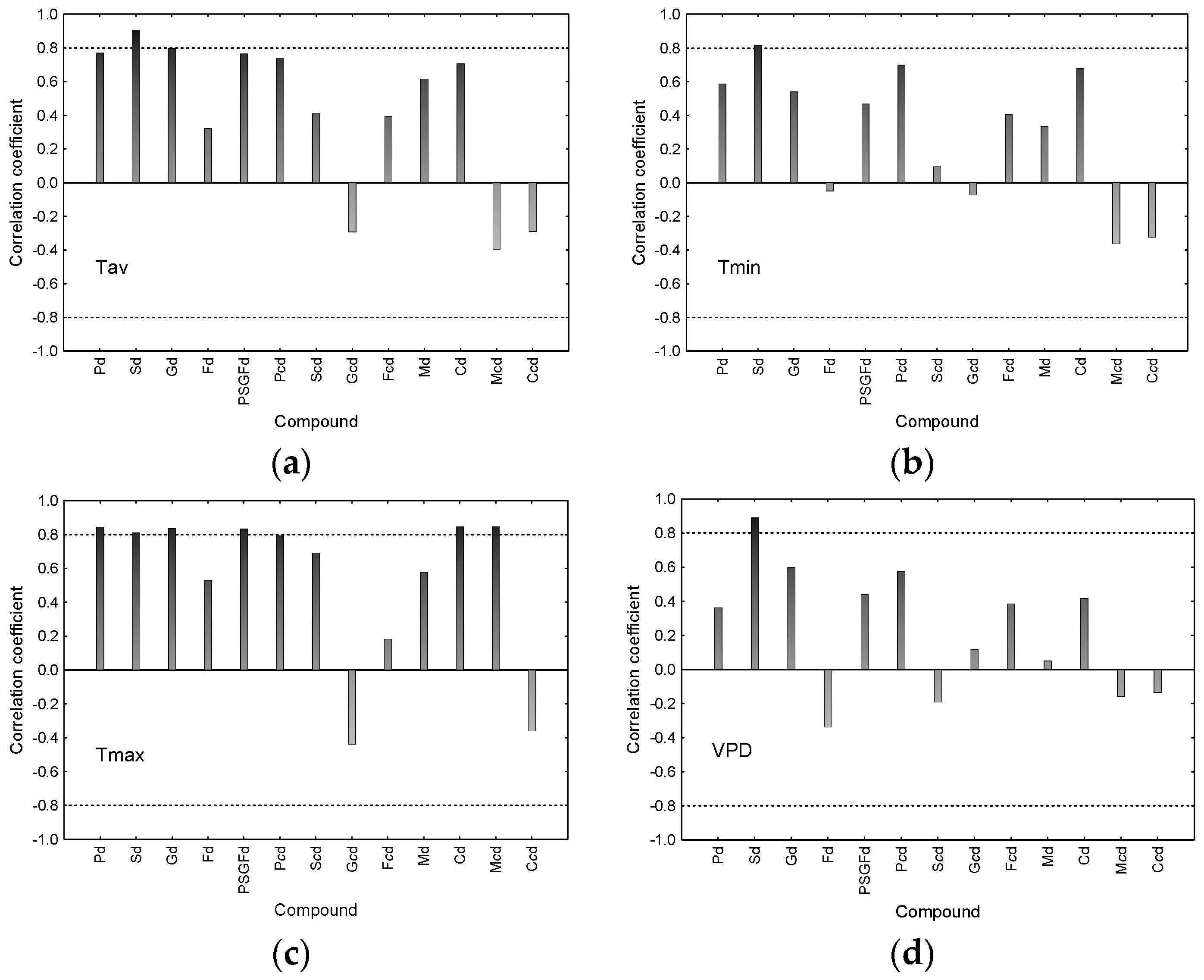
3.3. Correlations between δ13C and Concentrations of Individual Compounds in Declining Trees with Climate Data
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Fischer, E.M.; Knutti, R.; Lustenberger, A.; Schär, C. Expected changes in extremes. In Swiss Climate Change Scenarios CH 2011; C2SM, Meteo Swiss, ETH, NCCR Climate, and OcCC: Zürich, Switzerland, 2011; 88p. [Google Scholar]
- Anderegg, W.R.; Kane, J.M.; Anderegg, L.D. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- McDowell, N.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669–672. [Google Scholar] [CrossRef]
- IPCC; Pachauri, R.K.; Meyer, L.A. (Eds.) Climate Change 2014: Synthesis Report Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- Bigler, C.; Braker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an inciting mortality factor in Scots pine stands of the Valais. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef]
- Zweifel, R.; Rigling, A.; Dobbertin, M. Species-specific stomatal response of trees to drought—A link to vegetation dynamics? J. Veg. Sci. 2009, 20, 442–454. [Google Scholar] [CrossRef]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Graf Pannatier, E.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timofeeva, G.; Treydte, K.; Bugmann, H.; Rigling, A.; Schaub, M.; Siegwolf, R.; Saurer, M. Long-term effects of drought on tree-ring growth and carbon isotope variability in Scots pine in a dry environment. Tree Physiol. 2017, 37, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Solar, J. Effect of climate change on mountain pine distribution in western Tatra Mountains. In Climate Change—Realities, Impacts Over Ice Cap, Sea level and Risks; Singh, B.R., Ed.; Intech: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef]
- Brang, P. Decline of mountain pine (Pinus mugo ssp. uncinata) stands in the Swiss National Park: A dendrochronological approach. Dendrochronologia 1988, 6, 151–162. [Google Scholar]
- Dobbertin, M.; Baltensweiler, A.; Rigling, D. Tree mortality in an unmanaged mountain pine (Pinus mugo var. uncinata) stand in the Swiss National Park impacted by root fungi. For. Ecol. Manag. 2001, 145, 79–89. [Google Scholar] [CrossRef]
- Cherubini, P.; Fontana, G.; Rigling, D.; Dobbertin, M.; Brang, P.; Innes, J.L. Tree-life history prior to death: Two fungal root pathogens affect tree-ring growth differently. J. Ecol. 2002, 90, 839–850. [Google Scholar] [CrossRef]
- Bendel, M. The Spread of Root Rot Fungi in Mountain Pine Stands in the Swiss National Park: A Case Study of Its Influence on Forest Dynamic. Ph.D. Thesis, ETH, Zurich, Switzerland, 2006; 100p. [Google Scholar]
- Magel, E.; Einig, W.; Hampp, R. Carbohydrates in trees. In Carbohydrate Reserves in Plant—Synthesis and Regulation; Guppa, A.K., Kaur, N., Eds.; Elsevier Science BV: Amsterdam, The Netherlands, 2000; pp. 317–336. [Google Scholar]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytologist. 2016, 211, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Sevanto, S.; Dickman, L.T. Where does the carbon go?—Plant carbon allocation under climate change. Tree Physiol. 2015, 35, 581–584. [Google Scholar] [CrossRef] [PubMed]
- Rinne, K.T.; Saurer, M.; Kirdyanov, A.V.; Bryukhanova, M.V.; Prokushkin, A.S.; Churakova (Sidorova), O.V.; Siegwolf, R.T.W. Examining the response of needles carbohydrates from Siberian larch trees to climate using compound-specific δ13C and concentration analyses. Plant Cell Environ. 2015, 38, 2340–2352. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.M.; Wegener, F.; Werner, R.A.; Werner, C. Diel variations in carbon isotopic composition and concentration of organic acids and their impact on plant dark respiration in different species. Plant Biol. 2016, 18, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.M.; Rinne, K.T.; Blessing, C.; Siegwolf, R.T.W.; Buchmann, N.; Werner, R.A. Malate as a key carbon source of leaf dark-respired CO2 across different environmental conditions in potato plants. Exp. Bot. 2015, 66, 5769–5781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simard, S.; Giovanelli, A.; Treydte, K.; Traversi, M.L.; King, G.M.; Frank, D.; Fonti, P. Intra-annual dynamics of non-structural carbohydrates in the cambium of mature conifer trees reflects radial growth demands. Tree Physiol. 2013, 33, 913–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streit, K.; Siegwolf, R.; Hagerdon, F.; Schaub, M.; Buchmann, N. Lack of photosynthetic or stomatal regulation after 9 years of elevated [CO2] and 4 years of soil warming in two conifer species at the alpine treeline. Plant Cell Environ. 2014, 37, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Wild, B.; Wanek, W.; Postl, W.; Richter, A. Contribution of carbon fixed by Rubisco and PEPC to phloem export in the Crassulacean acid metabolism plant Kalanchoe daigremontiana. J. Exp. Biol. 2010, 61, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Hettmann, E.; Gleixner, G.; Juchelka, D. irm-LC/MS. δ13C Analysis of Organic Acids in Plants; Application Note 30075; Thermo Fischer Scientific: Ashville, NC, USA, 2005. [Google Scholar]
- Balicka, A. Untersuchung von Bergföhren im Schweizerischen Nationalpark mit Analyse Stabiler Isotope. Bachelor Thesis, ETH, Zürich, Switzerland, 2014. 31s. [Google Scholar]
- Churakova (Sidorova), O.V.; Lehmann, M.M.; Siegwolf, R.T.W.; Saurer, M.; Fonti, M.V.; Schmid, L.; Timofeeva, G.; Rinne-Garmston (Rinne), K.T.; Bigler, C. Compound-specific carbon isotope patterns in needles of conifer tree species from the Swiss National Park under recent climate change. Clim. Chang. 2018. under review. [Google Scholar]
- Moing, A. Sugar alcohols as carbohydrate reserves in some higher plants. In Carbohydrate Reserves in Plant—Synthesis and Regulation; Guppa, A.K., Kaur, N., Eds.; Elsevier Science BV: Amsterdam, The Netherlands, 2000; pp. 337–358. [Google Scholar]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic acids: The pools of fixed carbon involved in redox regulation and energy balance in higher plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [PubMed]
- Churakova (Sidorova), O.V.; Saurer, M.; Bryukhanova, M.; Siegwolf, R.; Bigler, C. Site-specific water-use strategies of mountain pine and larch to cope with recent climate change. Tree Physiol. 2016, 36, 942–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanek, W.; Heintel, S.; Richter, A. Preparation of starch and other carbon fractions from higher plan leaves for stable carbon isotope analysis. Rapid Commun. Mass Spectrom. 2001, 15, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Rinne, K.T.; Saurer, M.; Streit, K.; Siegwolf, R.T.W. Evaluation of a liquid chromatography method for compound-specific δ13C analysis of plant carbohydrates in alkane media. Rapid Commun. Mass Spectrom. 2012, 26, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon Isotope Discrimination and Photosynthesis. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Sudachkova, N.E.; Mulyutina, I.L.; Romanova, L.I.; Semenova, G.P. The annual dynamics of reserve compounds and hydrolitic enzymes activity in the tissues of Pinus sylvestris L. and Larix sibirica Ledeb. The metabolism of reserve compounds in the tissues of Siberian conifers. Eur. J. For. Res. 2004, 7, 1–10. [Google Scholar]
- Ainsworth, E.A.; Bush, D.R. Carbohydrate export from the leaf: A highly regulated process and target to enhance photosynthesis and productivity. Plant Physiol. 2011, 155, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Atkin, O.K.; Macherel, D. The crucial role of plant mitochondria in orchestrating drought tolerance. Ann. Bot. 2009, 103, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Granot, D.; David-Schwartz, R.; Kelly, G. Hexose kinases and their role in sugar-sensing and plant development. Front. Plant Sci. 2013, 4, 44. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.R.; Delhaize, E.; Jones, D.L. Function and mechanism of organic anion exudation from plant roots. Plant Mol. Biol. 2001, 52, 527–560. [Google Scholar]
- Hu, J.; Moore, D.J.P.; Monson, R.K. Weather and climate controls over the seasonal carbon isotope dynamics of sugars from subalpine forest trees. Plant Cell Environ. 2010, 33, 35–47. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Churakova, O.V.; Lehmann, M.M.; Saurer, M.; Fonti, M.V.; Siegwolf, R.T.W.; Bigler, C. Compound-Specific Carbon Isotopes and Concentrations of Carbohydrates and Organic Acids as Indicators of Tree Decline in Mountain Pine. Forests 2018, 9, 363. https://doi.org/10.3390/f9060363
Churakova OV, Lehmann MM, Saurer M, Fonti MV, Siegwolf RTW, Bigler C. Compound-Specific Carbon Isotopes and Concentrations of Carbohydrates and Organic Acids as Indicators of Tree Decline in Mountain Pine. Forests. 2018; 9(6):363. https://doi.org/10.3390/f9060363
Chicago/Turabian StyleChurakova (Sidorova), Olga V., Marco M. Lehmann, Matthias Saurer, Marina V. Fonti, Rolf T. W. Siegwolf, and Christof Bigler. 2018. "Compound-Specific Carbon Isotopes and Concentrations of Carbohydrates and Organic Acids as Indicators of Tree Decline in Mountain Pine" Forests 9, no. 6: 363. https://doi.org/10.3390/f9060363
APA StyleChurakova, O. V., Lehmann, M. M., Saurer, M., Fonti, M. V., Siegwolf, R. T. W., & Bigler, C. (2018). Compound-Specific Carbon Isotopes and Concentrations of Carbohydrates and Organic Acids as Indicators of Tree Decline in Mountain Pine. Forests, 9(6), 363. https://doi.org/10.3390/f9060363