Age-Effect on Intra-Annual δ13C-Variability within Scots Pine Tree-Rings from Central Siberia

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
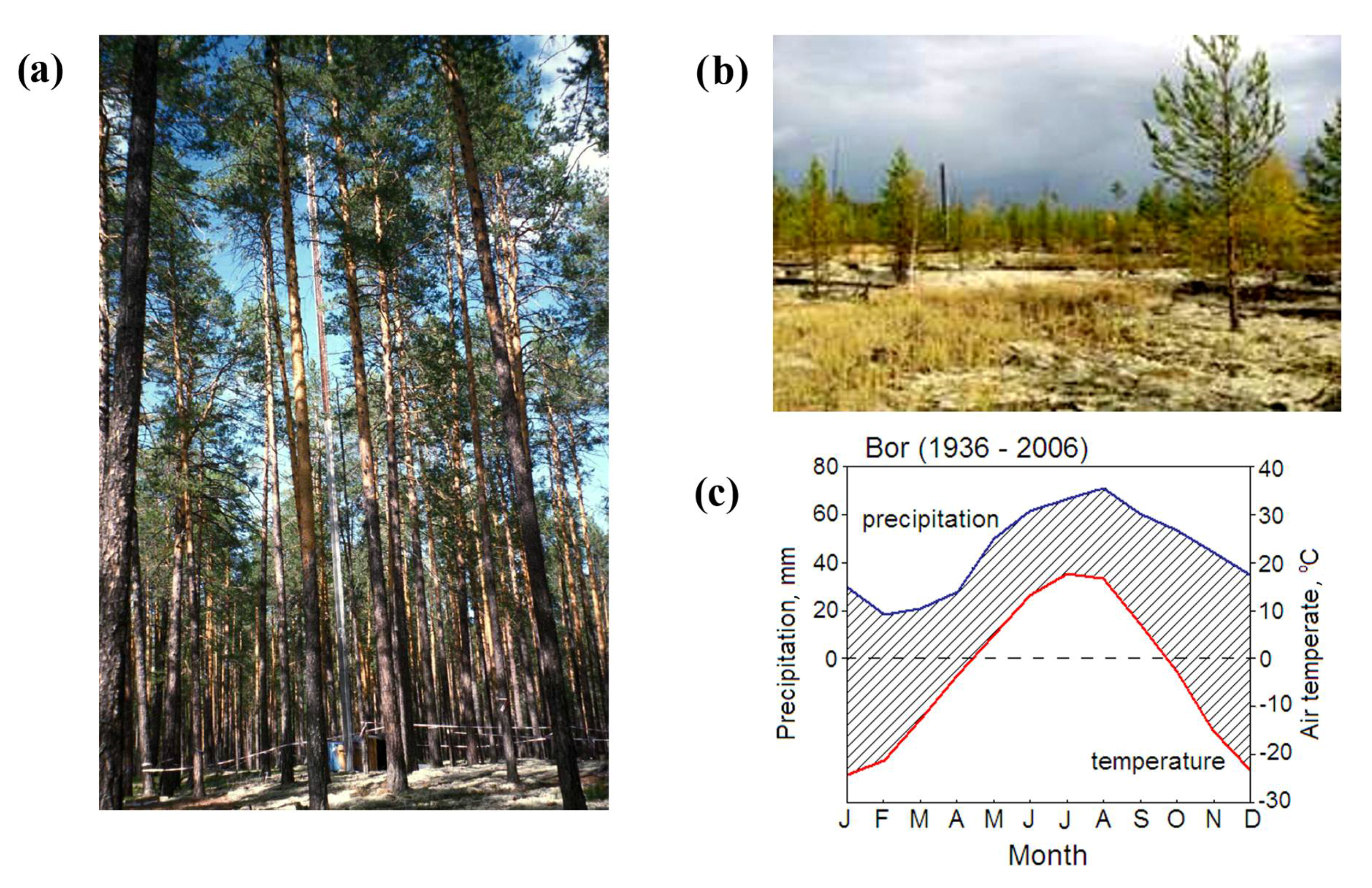
2.1. Study Sites and Climate
2.2. Tree Sampling and Dendrochronological Analysis
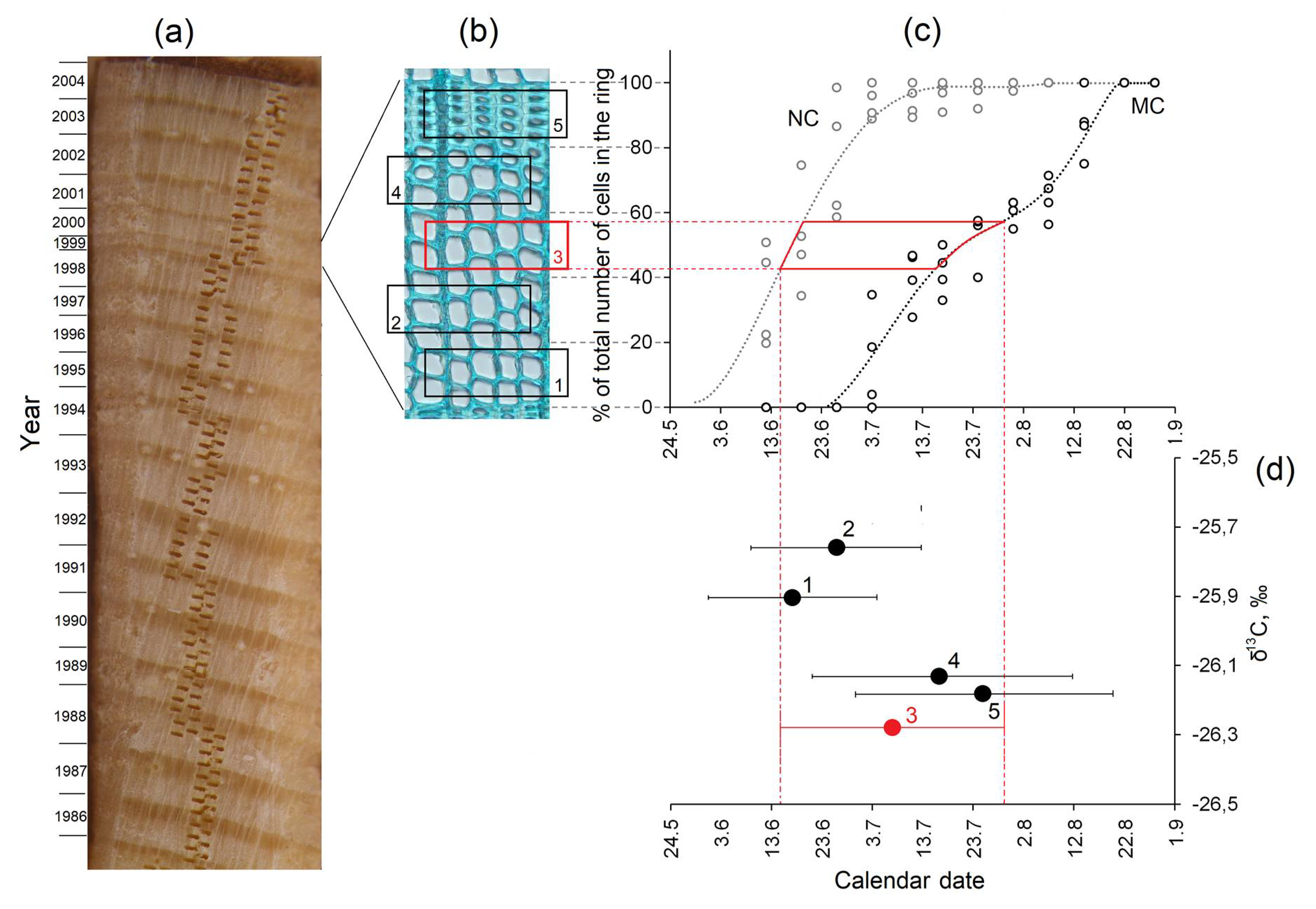
2.3. Image Analysis of Tracheid Dimensions
2.4. Observation of Wood Formation
2.5. Carbon Isotope Analysis
2.6. Climate-Growth Relationship and Statistical Analysis
3. Results
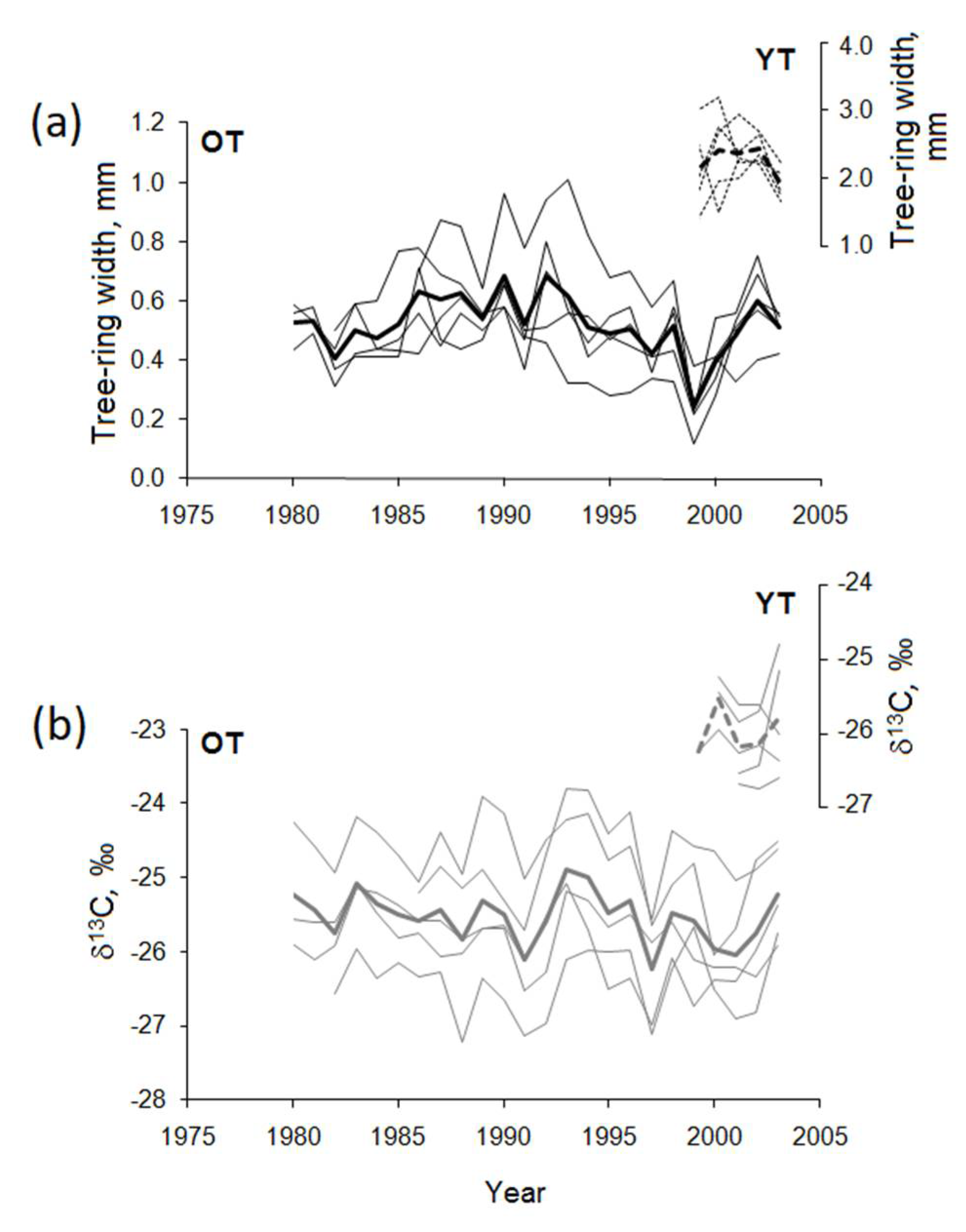
3.1. Time-Series and Correlations Among Parameters
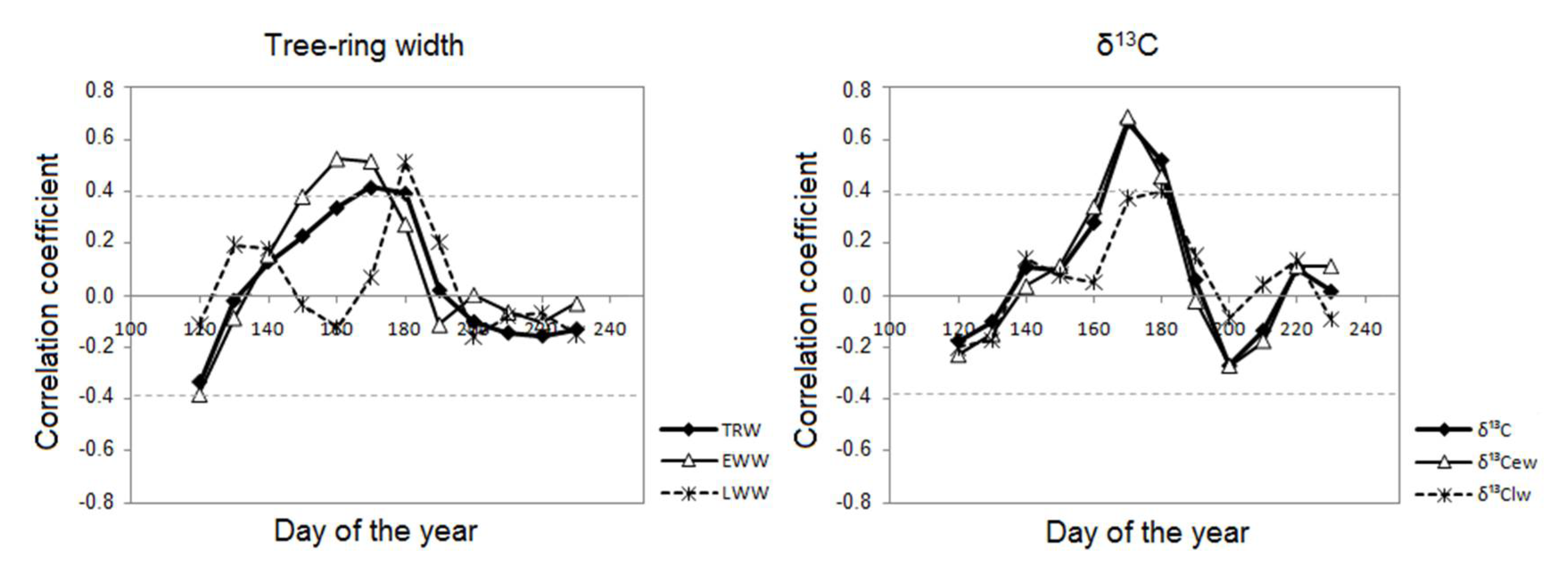
3.2. Tree-Ring Width and Average-Annual δ13С Responses to Climate
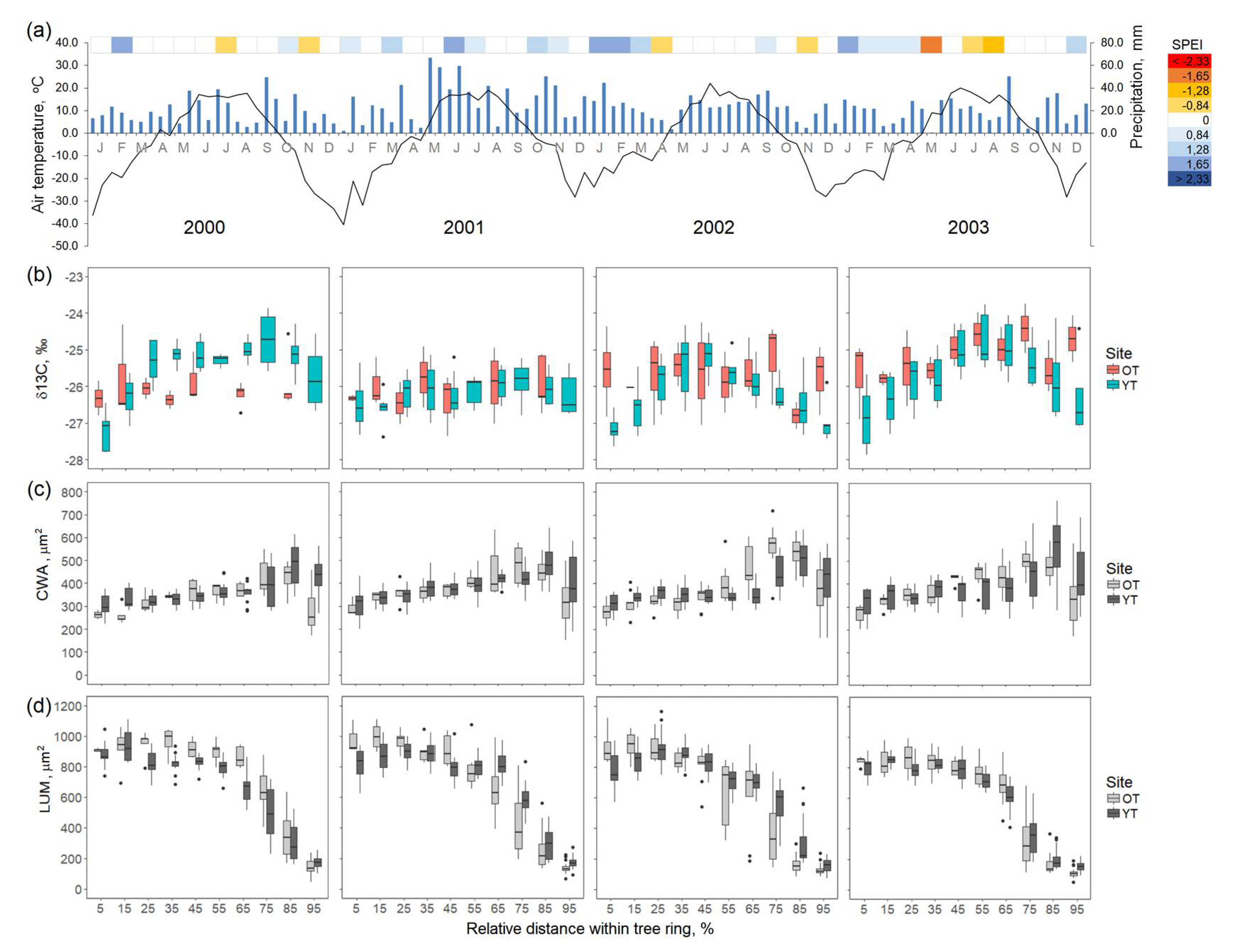
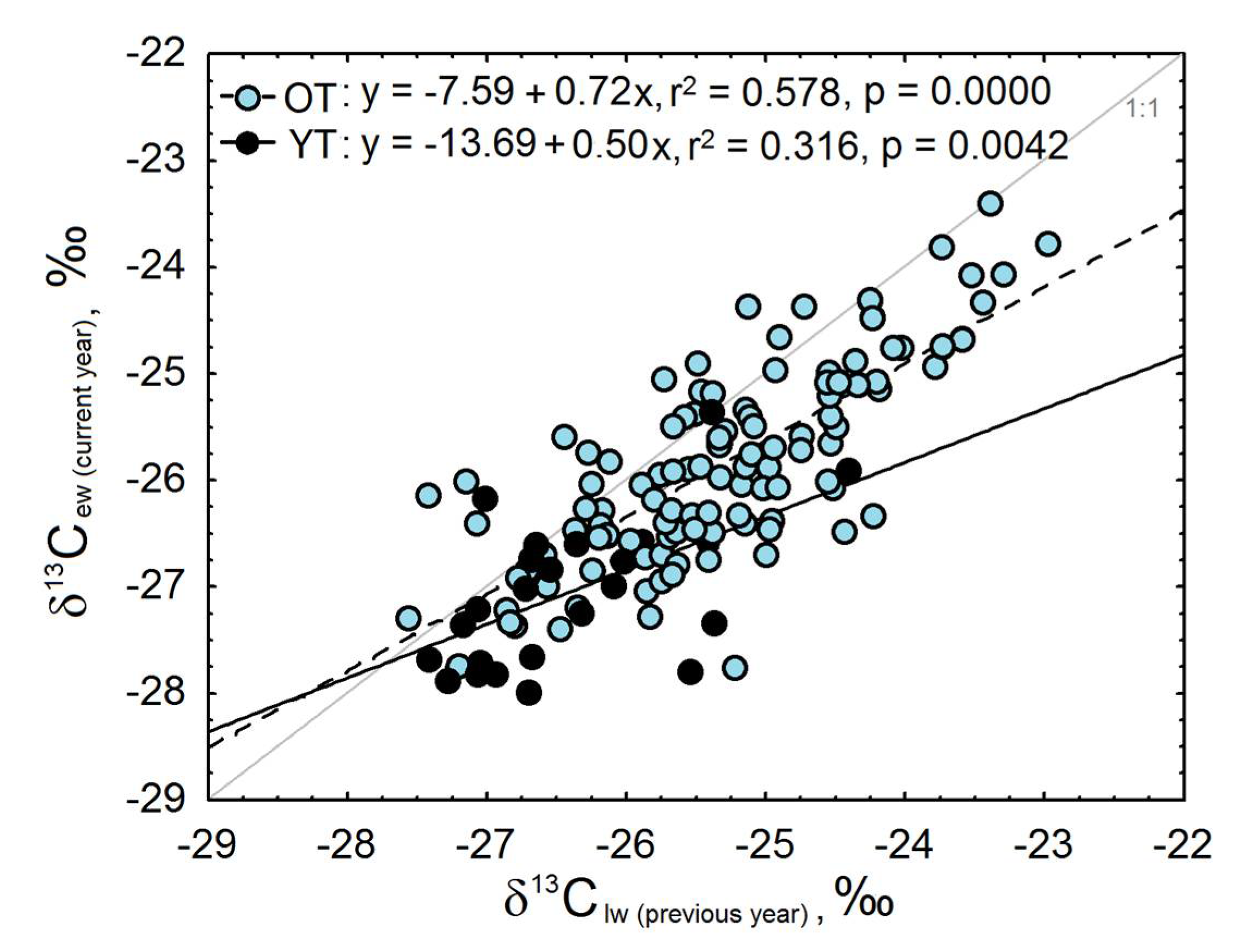
3.3. Intra-Ring Variability of Isotopic Composition and Anatomical Features of Young and Old Trees
3.4. Xylem Development and Intra-Annual δ13C
4. Discussion
4.1. Different Climatic Signal and Sensitivity between OT and YT
4.2. Growth Rate, Carbon Storage and Tree Size Might Explain the Differences
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Meinzer, F.C.; Lachenbruch, B.; Dawson, T.E. Size- and Age-Related Changes in Tree Structure and Function; Springer: Dordrecht, The Netherlands, 2011; Volume 4, 514p, ISBN 978-94-007-1241-6. [Google Scholar]
- Schweingruber, F.H. Tree Rings and Environment Dendroecology; Paul Haupt: Bern, Switzerland, 1996; 609p. [Google Scholar]
- Fritts, H. Tree Rings and Climate; Academic Press: London, UK; New York, NY, USA, 1976; 582p, ISBN 9780323145282. [Google Scholar]
- Brienen, R.J.W.; Gloor, E.; Clerici, S.; Newton, R.; Arppe, L.; Boom, A.; Bottrell, S.; Callaghan, M.; Heaton, T.; Helama, S.; et al. Tree height strongly affects estimates of water-use efficiency responses to climate and CO2 using isotopes. Nat. Commun. 2017, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology. Application in the Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; 394p, ISBN 978-0-7923-0586-6. [Google Scholar]
- Fonti, P.; Von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Battipaglia, G.; Brand, W.A.; Linke, P.; Saurer, M.; Aronne, G.; Cherubini, P. Discrete versus continuous analysis of anatomical and δ13C variability in tree rings with intra-annual density fluctuations. Trees Struct. Funct. 2012, 26, 513–524. [Google Scholar] [CrossRef]
- Rinne, K.T.; Saurer, M.; Kirdyanov, A.V.; Loader, N.; Bryukhanova, M.V.; Werner, R.; Siegwolf, R.T.W. The relationship between needle sugar carbon isotope ratios and tree rings of larch in Siberia. Tree Physiol. 2015, 35, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Fritts, H.C.; Vaganov, E.A.; Sviderskaya, I.V.; Shashkin, A.V. Climatic variation and tree-ring structure in conifers: Empirical and mechanistic models of tree-ring width, number of cells, cell size, cell-wall thickness and wood density. Clim. Res. 1991, 1, 97–116. [Google Scholar] [CrossRef]
- Pumijumnong, N.; Park, W.K. Vessel chronologies from teak in northern Thailand and their climatic signal. IAWA J. 1999, 20, 285–294. [Google Scholar] [CrossRef]
- Panyushkina, I.P.; Hughes, M.K.; Vaganov, E.A.; Munro, M.A.R. Summer temperature in northeastern Siberia since 1642 reconstructed from tracheid dimensions and cell numbers of Larix cajanderi. Can. J. For. Res. 2003, 33, 1905–1914. [Google Scholar] [CrossRef]
- Bryukhanova, M.V.; Fonti, P.; Kirdyanov, A.V.; Siegwolf, R.T.W.; Saurer, M.; Pochebyt, N.P.; Churakova (Sidorova), O.V.; Prokushkin, A.S. The response of δ13C, δ18O and cell anatomy of Larix gmelinii tree rings to differing soil active layer depths. Dendrochronologia 2015, 34, 51–59. [Google Scholar] [CrossRef]
- Fonti, P.; Babushkina, E.A. Tracheid anatomical responses to climate in a forest-steppe in Southern Siberia. Dendrochronologia 2016, 39, 32–41. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Xylogenesis of Pinus pinaster under a Mediterranean climate. Ann. For. Sci. 2014, 71, 71–80. [Google Scholar] [CrossRef]
- Steppe, K.; Sterck, F.; Deslauriers, A. Diel growth dynamics in tree stems: Linking anatomy and ecophysiology. Trends Plant Sci. 2015, 20, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Vaganov, E.A.; Hughes, M.K.; Shashkin, A.V. Growth Dynamics of Conifer Tree Rings: Images of Past and Future Environments; Springer: Berlin/Heidelberg, Germany, 2006; Volume 183, 354p, ISBN 978-3-540-26086-8. [Google Scholar]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Mäkinen, H.; Prislan, P.; Rossi, S.; Martinez del Castillo, E.; Campelo, F.; Vavrčík, H.; et al. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, A.; Sugimoto, A.; Maximov, T.C. Seasonal course of translocation, storage and remobilization of 13CO2 pulse-labelling photoassimilate in naturally growing Larix gmelini saplings. New Phytol. 2006, 171, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Skomarkova, M.V.; Vaganov, E.A.; Mund, M.; Knohl, A.; Linke, P.; Boerner, A.; Schulze, E.-D. Inter-annual and seasonal variability of radial growth, wood density and carbon isotope ratios 13С/12С in tree rings of beech (Fagus sylvatica) growing in Germany and Italy. Trees Struct. Funct. 2006, 20, 571–586. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Schulze, Е.-D.; Skomarkova, M.V.; Knohl, A.; Brand, W.A.; Roscher, C. Intra-annual variability of anatomical structure and δ13С values within tree rings of spruce and pine in alpine, temperate and boreal Europe. Oecologia 2009, 161, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Krepkowski, J.; Gebrekirstos, A.; Shibistova, O.; Bräuning, A. Stable carbon isotope labeling reveals different carry-over effects between functional types of tropical trees in an Ethiopian mountain forest. New Phytol. 2013, 199, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campelo, F.; Vieira, J.; Battipaglia, G.; de Luis, M.; Nabais, C.; Freitas, H.; Cherubini, P. Which matters most for the formation of intra-annual density fluctuations in Pinus pinaster: Age or size? Trees Struct. Funct. 2015, 29, 237–245. [Google Scholar] [CrossRef]
- Connor, K.F.; Lanner, R.M. Effects of tree age on secondary xylem and phloem anatomy in stems of Great Basin bristlecone pine (Pinus longaeuai). Am. J. Bot. 1990, 77, 1070–1077. [Google Scholar] [CrossRef]
- Briand, C.H.; Posluszny, U.; Larson, D.W. Influence of age and growth rate on radial anatomy of annual rings of Thuja occidentalis L. (eastern white cedar). Int. J. Plant Sci. 1993, 154, 406–411. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carrer, M. Age-dependent xylogenesis in timberline conifers. New Phytol. 2008, 177, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Bryukhanova, M.V.; Kirdyanov, A.V.; Prokushkin, A.S.; Silkin, P.P. Specific features of xylogenesis in Dahurian larch, Larix gmelinii (Rupr.) Rupr., growing on permafrost soils in Middle Siberia. Russ. J. Ecol. 2013, 44, 361–366. [Google Scholar] [CrossRef]
- Montwé, D.; Spiecker, H.; Hamann, A. An experimentally controlled extreme drought in a Norway spruce forest reveals fast hydraulic response and subsequent recovery of growth rates. Trees Struct. Funct. 2014, 28, 891–900. [Google Scholar] [CrossRef]
- Gessler, A.; Ferrio, J.P.; Hommel, R.; Treydte, K.; Werner, R.A.; Monson, R.K. Stable isotopes in tree rings: Towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 2014, 34, 796–818. [Google Scholar] [CrossRef] [PubMed]
- Prendin, A.L.; Petit, G.; Fonti, P.; Rixen, C.; Dawes, M.A.; von Arx, G. Axial xylem architecture of Larix decidua exposed to CO2 enrichment and soil warming at the treeline. Funct. Ecol. 2018, 32, 273–287. [Google Scholar] [CrossRef]
- Furyaev, V.V.; Vaganov, E.A.; Tchebakova, N.M.; Valendik, E.N. Effects of fire and climate on succession and structural changes in the Siberian boreal forest. Eur. J. For. Res. 2001, 2, 1–15. [Google Scholar]
- Wirth, C.; Schulze, E.-D.; Schulze, W.; von Stunzner-Karbe, D.; Ziegler, W.; Miljukowa, I.M.; Sogatchev, A.; Varlagin, A.B.; Panvyorov, M.; Grigoriev, S.; et al. Above-ground biomassand structure of pristine Siberian Scots pine forests as controlled by competition and fire. Oecologia 1999, 121, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Wirth, C.; Schulze, E.-D.; Luhker, B.; Grigoriev, S.; Siry, M.; Hardes, G.; Ziegler, W.; Backor, M.; Bauer, G.; Vygodskaya, N.N. Fire and site type effects on the long-term carbon and nitrogen balance in pristine Siberian Scots pine forests. Plant Soil 2002, 242, 41–63. [Google Scholar] [CrossRef]
- Panov, A.V.; Onuchin, A.A.; Zrazhevskaya, G.K.; Shibistova, O.B. Structure and dynamics of organic matter pools in clearings in the lichen pine forests of middle taiga subzone of Yenisei Siberia. Izvestiya Akademii Nauk Seriya Biologicheskaya 2012, 6, 658–666. (In Russian) [Google Scholar] [CrossRef]
- Glebov, F.Z. Bogs and Wetlands of the Forest Zone of the Yenisei Left-Bank; Nauka: Moscow, Russia, 1969; 132p. (In Russian) [Google Scholar]
- Walter, H.; Lieth, H. Klimadiagramm Weltatlas; Fischer: Jena, Germany, 1967. [Google Scholar]
- Rinn, F. Tsap V 3.6 Reference Manual: Computer Program for Tree-Ring Analysis and Presentation; Bierhelder weg 20, D-69126; RINNTECH: Heidelberg, Germany, 1996; 263p. [Google Scholar]
- Cook, E.R.; Peters, K. The smoothing spline: A new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree Ring Bull. 1981, 41, 45–53. [Google Scholar]
- Holmes, R.L. Program COFECHA: Version 3; The University of Arizona: Tucson, AZ, USA, 1992. [Google Scholar]
- Furst, G.G. Methods of Anatomical and Histochemical Research of Plant Tissue; Nauka: Moscow, Russia, 1979; 156p. (In Russian) [Google Scholar]
- Vaganov, E.A.; Shashkin, A.V.; Sviderskaya, I.V.; Vysotskaya, L.G. Histometric Analysis of Woody Plant Growth; Nauka: Novosibirsk, Russia, 1985; 108p. (In Russian) [Google Scholar]
- Munro, M.A.R.; Brown, P.M.; Hughes, M.K.; Garcia, E.M.R. Image analysis of tracheid dimensions for dendrochronological use. In Tree Rings, Environment and Humanity, Proceedings of the International Conference, Tucson, Arizona, 17–21 May 1994; Dean, J.S., Meko, D.M., Swetnam, T.W., Eds.; Radiocarbon: Tucson, AZ, USA, 1996; pp. 843–852. [Google Scholar]
- Denne, M.P. Definition of latewood according to Mork (1928). IAWA J. 1989, 10, 59–62. [Google Scholar] [CrossRef]
- Vaganov, E.A. The traheidogram method in tree-ring analysis and its application. In Methods of Dendrochronology. Application in Environmental Sciences; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1990; pp. 63–75. ISBN 978-0-7923-0586-6. [Google Scholar]
- Astrahantseva, N.V.; Cherkashin, V.P.; Stasova, V.V.; Antonova, G.F. The structure and development of secondary xylem and secondary phloem in the stems of Pinus sylvestris (Pinaceae) trees with different growth rates. Russ. J. Bot. (Botanicheskiy zhurnal) 2010, 95, 190–202. (In Russian) [Google Scholar]
- Schulze, B.; Wirth, C.; Linke, P.; Brand, W.A.; Kuhlmann, I.; Horna, V.; Schulze, E.-D. Laser-Ablation-Combustion-GC-IRMS—A new method for online analysis of intra-annual variation of δ¹³С in tree ring. Tree Physiol. 2004, 24, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Guerrieri, R.; Jennings, K.; Belmecheri, S.; Asbjornsen, H.; Ollinger, S. Evaluating climate signal recorded in tree-ring δ13C and δ18O values from bulk wood and α-cellulose for six species across four sites in the northeastern US. Rapid Commun. Mass Spectrom. 2017, 31, 2081–2091. [Google Scholar] [CrossRef] [PubMed]
- Benkova, V.E.; Shashkin, A.V.; Naurzbaev, M.M.; Prokushkin, A.S.; Simanko, V.V. The importance of microecological conditions for growth of Larix gmelinii at the timberline on Taimyr Peninsula. Russ. J. For. (Lesovedenie) 2012, 5, 59–70. (In Russian) [Google Scholar]
- Fonti, P.; Bryukhanova, M.V.; Myglan, V.S.; Kirdyanov, A.V.; Naumova, O.V.; Vaganov, E.A. Temperature-induced responses of xylem structure of Larix Sibirica Ldb. (Pinaceae) from Russian Altay. Am. J. Bot. 2013, 100, 1332–1343. [Google Scholar] [CrossRef] [PubMed]
- Vincente-Serrano, M.S.; Begueria, S.; Lopez-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Anfodillo, T.; Petit, G.; Crivellaro, A. Axial conduit widening in woody species: A still neglected anatomical pattern. IAWA J. 2013, 34, 352–364. [Google Scholar] [CrossRef]
- Carrer, M.; Von Arx, G.; Castagneri, D.; Petit, G. Distilling allometric and environmental information from time series of conduit size: The standardization issue and its relationship to tree hydraulic architecture. Tree Physiol. 2015, 35, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin Biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niinemets, Ü. Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiol. 2002, 22, 515–535. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Steppe, K.; Niinemets, Ü.; Teskey, R.O. Tree Size- and Age-Related Changes in Leaf Physiology and Their Influence on Carbon Gain. In Size- and Age-Related Changes in Tree Structure and Function; Meinzer, F.C., Lachenbruch, B., Dawson, T.E., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 4, pp. 235–253. ISBN 978-94-007-1241-6. [Google Scholar]
- Danis, P.-A.; Hatté, C.; Misson, L.; Guiot, J. MAIDENiso: A multiproxy biophysical model of tree-ring width and oxygen and carbon isotopes. Can. J. For. 2012, 42, 1697–1713. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Young Trees | Old Trees | ||||
|---|---|---|---|---|---|---|
| Ring | EW | LW | Ring | EW | LW | |
| Width, mm | 3.09 | 2.62 | 0.47 | 0.52 | 0.39 | 0.13 |
| Cell number | 73 | 55 | 18 | 15 | 10 | 5 |
| D, μm | 33.3 | 36.6 | 21.7 | 30.9 | 37.6 | 16.7 |
| CWT, μm | 3.7 | 3.1 | 5.7 | 3.7 | 3.3 | 4.7 |
| CWA, μm2 | 395.3 | 375.3 | 464.3 | 377.2 | 386.2 | 358.9 |
| LUM, μm2 | 603.0 | 723.7 | 187.6 | 550.3 | 744.8 | 150.5 |
| δ13С, ‰ | −25.93 | −25.89 | −26.04 | −25.50 | −25.61 | −25.19 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonti, M.V.; Vaganov, E.A.; Wirth, C.; Shashkin, A.V.; Astrakhantseva, N.V.; Schulze, Е.-D. Age-Effect on Intra-Annual δ13C-Variability within Scots Pine Tree-Rings from Central Siberia. Forests 2018, 9, 364. https://doi.org/10.3390/f9060364
Fonti MV, Vaganov EA, Wirth C, Shashkin AV, Astrakhantseva NV, Schulze Е-D. Age-Effect on Intra-Annual δ13C-Variability within Scots Pine Tree-Rings from Central Siberia. Forests. 2018; 9(6):364. https://doi.org/10.3390/f9060364
Chicago/Turabian StyleFonti, Marina V., Eugene A. Vaganov, Christian Wirth, Alexander V. Shashkin, Natalya V. Astrakhantseva, and Еrnst-Detlef Schulze. 2018. "Age-Effect on Intra-Annual δ13C-Variability within Scots Pine Tree-Rings from Central Siberia" Forests 9, no. 6: 364. https://doi.org/10.3390/f9060364
APA StyleFonti, M. V., Vaganov, E. A., Wirth, C., Shashkin, A. V., Astrakhantseva, N. V., & Schulze, Е. -D. (2018). Age-Effect on Intra-Annual δ13C-Variability within Scots Pine Tree-Rings from Central Siberia. Forests, 9(6), 364. https://doi.org/10.3390/f9060364