Population Genetic Diversity of Quercus ilex subsp. ballota (Desf.) Samp. Reveals Divergence in Recent and Evolutionary Migration Rates in the Spanish Dehesas


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Populations, Plant Material, and DNA Extraction
2.2. Nuclear and Chloroplast Microsatellite Analysis
2.3. Data Analysis
2.3.1. Chloroplast DNA
2.3.2. Nuclear DNA
2.3.3. Inferring Migration
3. Results
3.1. Chloroplast DNA
3.2. Nuclear DNA
3.3. Past Demographic Change
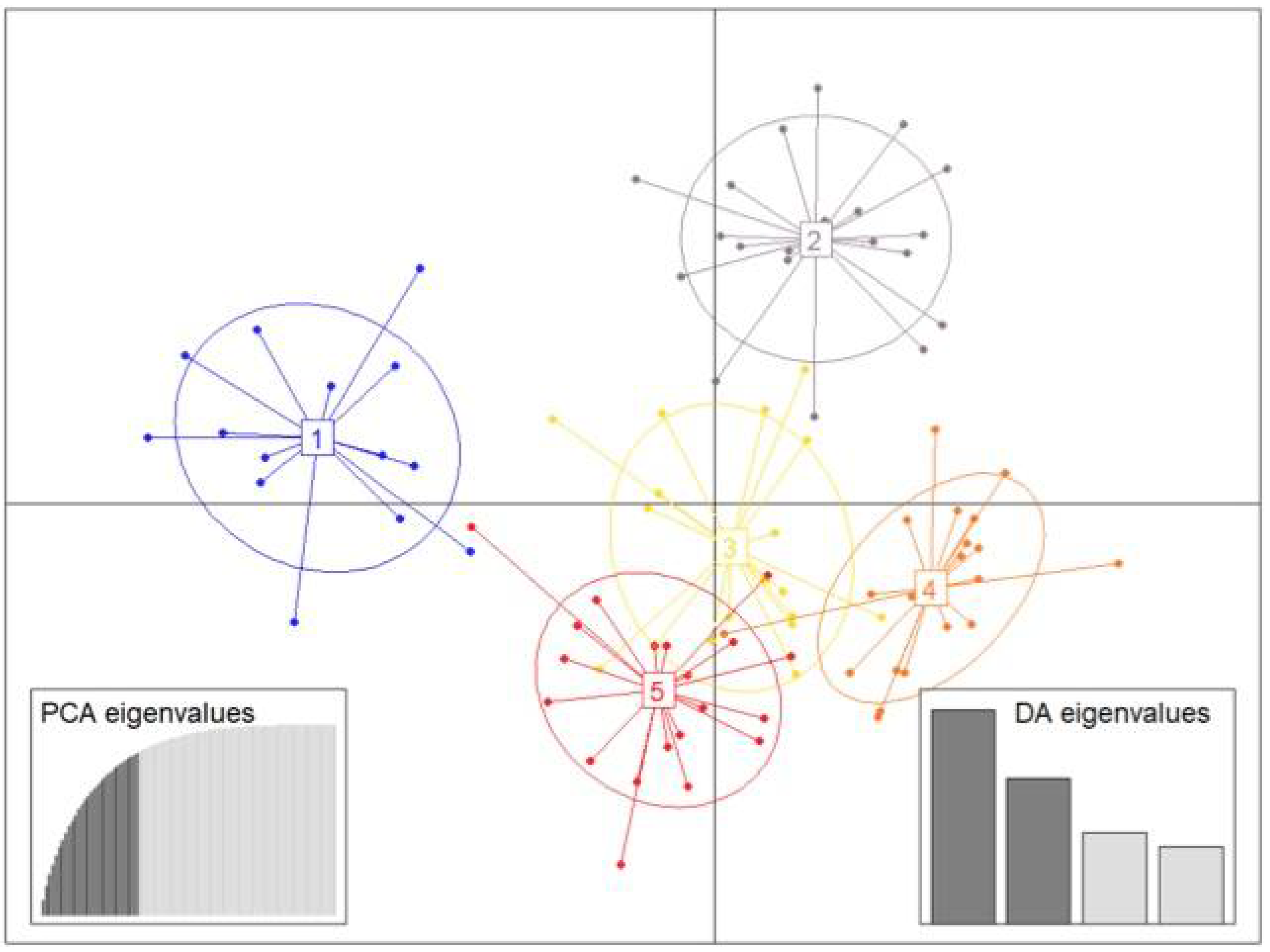
3.4. Population Structure
3.5. Migration
4. Discussion
4.1. Population Structure
4.2. Gene Flow
4.3. Association between Phenotypic Traits and Population Divergence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Moreno, G.; Pulido, F.J. The functioning, management and persistence of dehesas. In Agroforestry in Europe; Springer: Dordrecht, The Netherland, 2009; pp. 127–160. [Google Scholar]
- Brasier, C.M.; Robredo, F.; Ferraz, J.F.P. Evidence for Phytophthora cinnamomi involvement in Iberian oak decline. Plant Pathol. 1993, 42, 140–145. [Google Scholar] [CrossRef]
- Sghaier-Hammami, B.; Valero Galvàn, J.; Romero-Rodríguez, C.; Navarro-Cerrillo, R.M.; Abdelly, C.; Jorrín-Novo, J. Physiological and proteomics analyses of Holm oak Quercus ilex subsp. ballota [Desf.] Samp. responses to Phytophthora cinnamomi. Plant Physiol. Biochem. 2013, 71, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Pulido, F.J.; Díaz, M.; Hidalgo de Trucios, S.J. Size structure and regeneration of Spanish holm oak Quercus ilex forests and dehesas: Effects of agroforestry use on their long-term sustainability. For. Ecol. Manag. 2001, 146, 1–13. [Google Scholar] [CrossRef]
- Barbeta, A.; Peñuelas, J. Sequence of plant responses to droughts of different timescales: Lessons from holm oak Quercus ilex forests. Plant Ecol. Divers. 2016, 9, 321–338. [Google Scholar] [CrossRef]
- MAPA. Forestación de Tierras Agrícolas; Ministerio de Agricultura y Pesca: Madrid, Spain, 2006.
- Valero Galván, J.; González Fernandez, R.; Navarro Cerrillo, R.M.; Pelegrín, E.G.; Jorrín-Novo, J.V. Physiological and Proteomic Analyses of Drought Stress Response in Holm Oak Provenances. J. Proteome Res. 2013, 12, 5110–5123. [Google Scholar] [CrossRef] [PubMed]
- Corcobado, T.; Cubera, E.; Juárez, E.; Moreno, G.; Solla, A. Drought events determine performance of Quercus ilex seedlings and increase their susceptibility to Phytophthora cinnamomi. Agric. For. Meteorol. 2014, 192, 1–8. [Google Scholar] [CrossRef]
- Corcobado, T.; Miranda-Torres, J.J.; Martín-García, J.; Jung, T.; Solla, A. Early survival of Quercus ilex subspecies from different populations after infections and coinfections by multiple Phytophthora species. Plant Pathol. 2017, 665, 792–804. [Google Scholar] [CrossRef]
- Valero Galván, J.; Valledor, L.; Cerrillo, R.M.N.; Pelegrín, E.G.; Jorrín-Novo, J.V. Studies of variability in Holm oak Quercus ilex subsp. ballota [Desf.] Samp. through acorn protein profile analysis. J. Proteom. 2011, 74, 1244–1255. [Google Scholar] [CrossRef] [PubMed]
- Valero Galván, J.; Jorrín-Novo, J.; Cabrera, A.; Ariza, D.; García-Olmo, J.; Cerrillo, R. Population variability based on the morphometry and chemical composition of the acorn in Holm oak Quercus ilex subsp. ballota [Desf.] Samp. Eur. J. For. Res. 2012, 131, 893–904. [Google Scholar] [CrossRef]
- Valero Galván, J.; Valledor, L.; González Fernandez, R.; Navarro Cerrillo, R.M.; Jorrín-Novo, J.V. Proteomic analysis of Holm oak Quercus ilex subsp. ballota [Desf.] Samp. pollen. J. Proteom. 2012, 75, 2736–2744. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, B.; Rodríguez-López, C.M.; Forrest, A.; Cano, E.; Vargas, P. Protected areas of Spain preserve the neutral genetic diversity of Quercus ilex L. irrespective of glacial refugia. Tree Genet. Genom. 2015, 11, 124. [Google Scholar] [CrossRef]
- Vitelli, M.; Vessella, F.; Cardoni, S.; Pollegioni, P.; Denk, T.; Grimm, G.W.; Simeone, M.C. Phylogeographic structuring of plastome diversity in Mediterranean oaks (Quercus Group Ilex, Fagaceae). Tree Genet. Genom. 2017, 13, 3. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. B 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.G.; Cosson, J.F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Ortego, J.; Bonal, R.; Muñoz, A. Genetic consequences of habitat fragmentation in long-lived tree species: The case of the Mediterranean Holm Oak (Quercus ilex, L.). J. Hered. 2010, 101, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Lunt, D.H. Refugia within refugia: Patterns of phylogeographic concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia: Evolutionary Perspectives on the Origins and Conservation of European Biodiversity; Weiss, S., Ferrand, N., Eds.; Springer: Berlin, Germany, 2007; pp. 155–188. [Google Scholar]
- Sinclair, W.T.; Morman, J.D.; Ennos, R.A. The postglacial history of Scots pine (Pinus sylvestris L.) in Western Europe: Evidence from mitochondrial DNA variation. Mol. Ecol. 1999, 8, 83–88. [Google Scholar] [CrossRef]
- Soranzo, N.; Alia, R.; Provan, J.; Powell, W. Patterns of variation at a mitochondrial sequence-tagged-site locus provides new insights into the postglacial history of European Pinus sylvestris populations. Mol. Ecol. 2000, 9, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Csaikl, U.; Bordacs, S.; Burg, K.; Coart, E.; Cottrell, J.; Van Dam, B.; Deans, J.; Dumolin-Lapegue, S.; Fineschi, S.; et al. Chloroplast DNA variation in European white oaks: Phylogeography and patterns of diversity based on data from over 2600 populations. For. Ecol. Manag. 2002, 156, 5–26. [Google Scholar] [CrossRef]
- González-Martinez, S.C.; Gómez, A.; Carrión, J.S.; Agúndez, D.; Alía, R.; Gil, L. Spatial genetic structure of an explicit glacial refugium of maritime pine Pinus pinaster Aiton in southeastern Spain. In Phylogeography of Southern European Refugia: Evolutionary Perspectives on the Origins and Conservation of European Biodiversity; Weiss, S., Ferrand, N., Eds.; Springer: Berlin, Germany, 2007; pp. 257–269. [Google Scholar]
- Lumaret, R.; Mir, C.; Michaud, H.; Raynal, V. Phylogeographical variation of chloroplast DNA in Holm Oak (Quercus ilex L.). Mol. Ecol. 2002, 11, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.; Finlayson, C.; Fa, D.; Finlayson, G. Southern Iberia as a refuge for the last Neanderthal populations. J. Biogeogr. 2011, 38, 1873–1885. [Google Scholar] [CrossRef]
- Jorrín-Novo, J.V.; Pascual, J.; Lucas, R.S.; Romero-Rodriguez, C.; Ortega, M.R.; Lenz, C.; Valledor, L. Fourteen years of plant proteomics reflected in “Proteomics”: Moving from model species and 2-DE based approaches to orphan species and gel-free platforms. Proteomics 2015, 15, 1089–1112. [Google Scholar] [CrossRef] [PubMed]
- Echevarria-Zomeno, S.; Ariza, D.; Jorge, I.; Lenz, C.; Del Campo, A.; Jorrin, J.V.; Navarro, R.M. Changes in the protein profile of Quercus ilex leaves in response to drought stress and recovery. J. Plant Physiol. 2009, 166, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polzin, T.; Daneschmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Schroeder, K.B.; Rosenberg, N.A. A maximum-likelihood method to correct for allelic dropout in microsatellite data with no replicate genotypes. Genetics 2012, 192, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Goudet, J. FSTAT: A Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3.2). 2002. Available online: https://www.scienceopen.com/document?vid=79097bb4-ec3c-47c3-94a1-47085d721e6b (accessed on 15 January 2018).
- Piry, S.; Luikart, G.; Cornuet, J.M. Bottleneck: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, N.M.; Mayze, J.L.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, A. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Manni, F.; Guérard, E.; Heyer, E. Geographic patterns of genetic, morphologic, linguistic variation: How barriers can be detected by “Monmonier’s algorithm”. Hum. Biol. 2004, 762, 173–190. [Google Scholar] [CrossRef]
- Dieringer, D.; Schlötterer, C. Microsatellite analyser MSA: A platform independent analysis tool for large microsatellite data sets. Mol. Ecol. Notes 2003, 3, 167–169. [Google Scholar] [CrossRef]
- Wilson, G.A.; Rannala, B. Bayesian Inference of Recent Migration Rates Using Multilocus Genotypes. Genetics 2003, 163, 1177–1191. [Google Scholar] [PubMed]
- Rambaut, A.; Drummond, A.J. Tracer v1.4. 2007. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 15 January 2018).
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [PubMed]
- Gómez, J.M. Spatial patterns in long distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography 2003, 26, 573–584. [Google Scholar] [CrossRef]
- Gómez, J.M.; Valladares, F.; Puerta-Piñero, C. Differences between structural and functional environmental heterogeneity caused by seed dispersal. Funct. Ecol. 2004, 18, 787–792. [Google Scholar] [CrossRef] [Green Version]
- Soto, A.; Lorenzo, Z.; Gil, L. Differences in fine-scale genetic structure and dispersal in Quercus ilex L. and Q. suber L.: Consequences for regeneration of Mediterranean open woods. Heredity 2007, 99, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Smit, C.; Díaz, M.; Jansen, P. Establishment limitation of holm oak Quercus ilex subsp. ballota Desf. Samp. in a Mediterranean savanna–forest ecosystem. Ann. For. Sci. 2009, 66, 1–7. [Google Scholar] [CrossRef]
- García-Nogales, A.; Linares, J.C.; Laureano, R.G.; Seco, J.I.; Merino, J. Range wide variation in life history phenotypes: Spatiotemporal plasticity across the latitudinal gradient of the evergreen oak Quercus ilex. J. Biogeogr. 2016, 4312, 2366–2379. [Google Scholar] [CrossRef]
- Le Corre, V.; Machon, N.; Petit, R.J.; Kremer, A. Colonization with long-distance seed dispersal and genetic structure of maternally inherited genes in forest trees: A simulation study. Genet. Res. 1997, 69, 117–125. [Google Scholar] [CrossRef]
- Grivet, D.; Smouse, P.E.; Sork, V.L. A novel approach to an old problem: Tracking dispersed seeds. Mol. Ecol. 2005, 14, 3585–3595. [Google Scholar] [CrossRef] [PubMed]
- Dodd, R.S.; Afzal-Rafii, Z.; Mayer, W. Molecular markers show how pollen and seed dispersal affect population genetic structure in coast live oak Quercus agrifolia Née. In Proceedings of the Sixth Symposium on Oak Woodlands: Today’s Challenges, Tomorrow’s Opportunities; Merenlender, A., McCreary, D., Purcell, K.L., Eds.; PSW-GTR-217; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2008; pp. 485–495. [Google Scholar]
- Nettel, A.; Dodd, R.S.; Afzal-Rafii, Z. Genetic diversity, structure, and demographic change in tanoak, Lithocarpus densiflorus Fagaceae, the most susceptible host to the Sudden Oak Death disease in California. Am. J. Bot. 2009, 96, 2224–2233. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, A. Descriptive analysis of the ‘relictual’Mediterranean landscape in the Guadalquivir River valley southern Spain: A baseline for scientific research and the development of conservation action plans. Biodivers. Conserv. 2008, 17, 2219–2232. [Google Scholar] [CrossRef]
- Carrión-Marco, Y.; Ntinou, M.; Badal, E. Neolithic woodland in the north Mediterranean basin: A review on Olea europaea L. In Proceedings of the Fourth International Meeting of Anthracology; Damblon, F., Ed.; Series 2486; Archaeo Press: Oxford, UK, 2013. [Google Scholar]
- Fortuna, M.A.; Albaladejo, R.G.; Fernández, L.; Aparicio, A.; Bascompte, J. Networks of spatial genetic variation across species. Proc. Natl. Acad. USA 2009, 106, 19044–19049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casimiro-Soriguer, R.; Talavera, M.; Balao, F.; Terrab, A.; Herrera, J.; Talavera, S. Phylogeny and genetic structure of Erophaca (Leguminosae), a East–West Mediterranean disjunct genus from the Tertiary. Mol. Phylogenet. Evol. 2010, 56, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.Á.; Tremetsberger, K.; Terrab, A.; Stuessy, T.F.; García-Castaño, J.L.; Urtubey, E.; Baeza, C.M.; Ruas, C.F.; Gibbs, P.E.; Talavera, S. Phylogeography of the invasive weed Hypochaeris radicata (Asteraceae): From Moroccan origin to worldwide introduced populations. Mol. Ecol. 2008, 17, 3654–3667. [Google Scholar] [CrossRef] [PubMed]
- Carrión, J.S.; Sánchez-Gómez, P.; Mota, J.F.; Yll, R.; Chaín, C. Holocene vegetation dynamics, fire and grazing in the Sierra de Gádor, southern Spain. Holocene 2003, 13, 839–849. [Google Scholar] [CrossRef]
- Carrion, J.S.; Yll, E.I.; Walker, M.J.; Legaz, A.J.; Chain, C.; Lopez, A. Glacial refugia of temperate, Mediterranean and Ibero North African flora in south-eastern Spain: New evidence from cave pollen at two Neanderthal man sites. Glob. Ecol. Biogeogr. 2003, 12, 119–129. [Google Scholar] [CrossRef]
- Carrión, J.S.; Fernández, S.; Jiménez-Moreno, G.; Fauquette, S.; Gil-Romera, G.; González-Sampériz, P.; Finlayson, C. The historical origins of aridity and vegetation degradation in southeastern Spain. J. Arid Environ. 2010, 74, 731–736. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S.; Fernández, S.; González-Sampériz, P.; Gil-Romera, G.; Badal, E.; Carrión-Marco, Y.; López-Merino, L.; López-Sáez, J.A.; Fierro, E.; Burjachs, F. Expected trends and surprises in the Lateglacial and Holocene vegetation history of the Iberian Peninsula and Balearic Islands. Rev. Palaeobot. Palynol. 2010, 162, 458–475. [Google Scholar] [CrossRef] [Green Version]
- Gil-Romera, G.; Carrion, J.S.; Pausas, J.G.; Seville-Callejo, M.; Lamb, H.F.; Burjachs, F. Holocene fire activity and vegetation response in South-Eastern Iberia. Quat. Sci. Rev. 2010, 29, 1082–1092. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Lees, K.; Montgomery, M.E.; England, P.R.; Lowe, E.; Briscoe, D.A. Do population size bottlenecks reduce evolutionary potential? Anim. Conserv. 1999, 2, 255–260. [Google Scholar] [CrossRef]
- Plieninger, T.; Pulido, F.J.; Schaich, H. Effects of land-use and landscape structure on holm oak recruitment and regeneration at farm level in Quercus ilex L. dehesas. J. Arid Environ. 2004, 57, 345–364. [Google Scholar] [CrossRef]
- Geri, F.; Amici, V.; Rocchini, D. Human activity impact on the heterogeneity of a Mediterranean landscape. Appl. Geogr. 2010, 30, 370–379. [Google Scholar] [CrossRef]
- Beerli, P. Migrate Version 3.0: A Máximum Likelihood and Bayesian Estimator of Gene Flow Using the Coalescent. 2008. Available online: http://popgen.scs.edu/migrate.html (accessed on 15 January 2018).
- Rodríguez-Ariza, M.O. Vegetation evolution and use during prehistory in the upper Guadalquivir. In MENGA 02. Journal of Andalusian Prehistory; Junta de Andalucia: Malaga, Spain, 2011. [Google Scholar]
- Garza, J.C.; Williamson, E.G. Detection of reduction in population size using data from microsatellite loci. Mol. Ecol. 2001, 10, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Chiucchi, J.E.; Gibbs, H.L. Similarity of contemporary and historical gene flow among highly-fragmented populations of an endangered rattlesnake. Mol Ecol. 2010, 19, 5345–5348. [Google Scholar] [CrossRef] [PubMed]
- Vicente, A.M.; Alés, R.F. Long term persistence of dehesas. Evidences from history. Agrofor. Syst. 2006, 67, 19–28. [Google Scholar] [CrossRef]
- Gómez-Casero, M.T.; Galán, C.; Domínguez-Vilches, E. Flowering phenology of Mediterranean Quercus species in different locations Córdoba, SW Iberian Peninsula. Acta Bot. Malacit. 2007, 32, 127–146. [Google Scholar]
- Felicísimo, A.M.; Muñóz, J.; Mateo, R.; Villalba, C.J. Vulnerabilidad de la flora y vegetación españolas ante el cambio climático. Ecosistemas 2012, 21, 1–6. [Google Scholar]
- Sáez, J.L.; García, P.L.; Sánchez, M.M. Análisis palinológico del yacimiento arqueológico de Pocito Chico El Puerto de Santa María: El paisaje prehistórico y protohistórico durante el Holoceno reciente en las Marismas de Cádiz. Cuaternario y Geomorfología 2001, 15, 45–59. [Google Scholar]
- Pérez-Jordà, G.; Peña-Chocarro, L.; Fernández, M.G.; Rodríguez, J.V. The beginnings of fruit tree cultivation in the Iberian Peninsula: Plant remains from the city of Huelva southern Spain. Veg. History Archaeobot. 2017, 26, 527–538. [Google Scholar] [CrossRef]
- Dow, B.D.; Ashley, M.V.; Howe, H.F. Characterization of highly variable (GA/CT)n microsatellites in the bur oak, Quercus macrocarpa. Theor. Appl. Genet. 1995, 91, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, S.; Lexer, C.; Glöss, J.; Steinkellner, H. Characterization of (GA)n microsatellite loci from Quercus robur. Hereditas 1998, 129, 183–186. [Google Scholar] [CrossRef]
- Steinkellner, H.; Fluch, S.; Turetscheck, E.; Lexer, C.; Streiff, R.; Kremer, A.; Burg, K.; Gloss, J. Identification and characterization of (GA/CT)—Microsatellite loci from Quercus petraea. Plant Mol. Biol. 1997, 33, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, L.; Scebba, F.; Tognetti, R. Heavy metal accumulation and growth responses in poplar clones Eridano (Populus deltoides × maximowiczii) and I.-214 (P. × euramericana) exposed to industrial waste. Environ. Exp. 2004, 52, 79–88. [Google Scholar] [CrossRef]
- Deguilloux, M.F.; Pemonge, M.H.; Petit, R.J. Use of chloroplast microsatellites to differentiate oak populations. Ann. For. Sci. 2004, 61, 825–830. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Code | Sample Size | Latitude (°N) | Longitude (°W) | Altitude (m) | Annual Precipitation (mm) | Mean Winter Precipitation (mm) | Mean Autumn Precipitation (mm) | Mean Monthly Maximum Temperature (°C) | Mean Monthly Minimum Temperature (°C) |
|---|---|---|---|---|---|---|---|---|---|---|
| Cádiz | BCA | 15 | 36.756628 | 5.452958 | 649 | 1263.6 | 375.1 | 408.9 | 24.9 | 9.8 |
| Almería | SAA | 20 | 36.858483 | 2.478264 | 1241 | 277.9 | 248.5 | 103.4 | 25.2 | 8.9 |
| Huelva | CTH | 19 | 37.919931 | 6.475167 | 364 | 845.6 | 310.2 | 348.4 | 26.3 | 9.5 |
| Córdoba | PCO | 20 | 38.324489 | 4.827869 | 618 | 612.6 | 211.7 | 229.9 | 26.8 | 8.1 |
| Seville | APS | 20 | 37.875678 | 6.474586 | 482 | 722.1 | 78.9 | 307.6 | 26.4 | 9.5 |
| Population | Chloroplast | Nuclear | |||
|---|---|---|---|---|---|
| No. Haplotypes | He | Mean No. Alleles per Locus | AR15 per Locus | FIS | |
| Cádiz | 7 | 0.71 (0.07) | 6.2 (0.9) | 6.2 (0.9) | −0.11 (0.10) |
| Almería | 7 | 0.71 (0.07) | 6.8 (1.1) | 6.4 (1.0) | −0.11 (0.12) |
| Huelva | 5 | 0.72 (0.07) | 7.8 (1.5) | 7.3 (1.4) | −0.03 (0.05) |
| Córdoba | 9 | 0.68 (0.07) | 5.8 (1.0) | 5.5 (0.9) | −0.05 (0.09) |
| Seville | 5 | 0.72 (0.07) | 7.4 (1.1) | 7.0 (1.0) | −0.05 (0.08) |
| Total | 30 | 0.74 (0.07) | 11.2 (1.6) | 7.8 (1.2) | −0.02 (0.07) |
| From | Cádiz | Almería | Huelva | Córdoba | Seville | |
|---|---|---|---|---|---|---|
| In | ||||||
| Cádiz | BA | 0.91 | 0.02 | 0.02 | 0.02 | 0.03 |
| M | — | 27.2 (0.14) | 26.1 (0.13) | 21.9 (0.11) | 18.7 (0.09) | |
| Almería | BA | 0.03 | 0.68 | 0.02 | 0.26 | 0.02 |
| M | 23.9 (0.12) | — | 22.5 (0.11) | 23.9 (0.12) | 34.8 (0.17) | |
| FST | 0.07 | — | — | — | — | |
| Huelva | BA | 0.02 | 0.02 | 0.68 | 0.26 | 0.02 |
| M | 26.5 (0.13) | 26.5 (0.13) | — | 20.4 (0.10) | 105.5 (0.53) | |
| FST | 0.06 | 0.05 | — | — | — | |
| Córdoba | BA | 0.05 | 0.02 | 0.02 | 0.89 | 0.02 |
| M | 1.2 (0.01) | 1.0 (0.01) | 4.2 (0.02) | — | 4.8 (0.02) | |
| FST | 0.07 | 0.06 | 0.04 | — | — | |
| Seville | BA | 0.06 | 0.02 | 0.02 | 0.12 | 0.79 |
| M | 37.2 (0.19) | 30.2 (0.15) | 29.7 (0.15) | 23.0 (0.12) | — | |
| FST | 0.07 | 0.06 | 0.04 | 0.04 | — |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández i Marti, A.; Romero-Rodríguez, C.; Navarro-Cerrillo, R.M.; Abril, N.; Jorrín-Novo, J.V.; Dodd, R.S. Population Genetic Diversity of Quercus ilex subsp. ballota (Desf.) Samp. Reveals Divergence in Recent and Evolutionary Migration Rates in the Spanish Dehesas. Forests 2018, 9, 337. https://doi.org/10.3390/f9060337
Fernández i Marti A, Romero-Rodríguez C, Navarro-Cerrillo RM, Abril N, Jorrín-Novo JV, Dodd RS. Population Genetic Diversity of Quercus ilex subsp. ballota (Desf.) Samp. Reveals Divergence in Recent and Evolutionary Migration Rates in the Spanish Dehesas. Forests. 2018; 9(6):337. https://doi.org/10.3390/f9060337
Chicago/Turabian StyleFernández i Marti, Angel, Cristina Romero-Rodríguez, Rafael M. Navarro-Cerrillo, Nieves Abril, Jesús V. Jorrín-Novo, and Richard S. Dodd. 2018. "Population Genetic Diversity of Quercus ilex subsp. ballota (Desf.) Samp. Reveals Divergence in Recent and Evolutionary Migration Rates in the Spanish Dehesas" Forests 9, no. 6: 337. https://doi.org/10.3390/f9060337
APA StyleFernández i Marti, A., Romero-Rodríguez, C., Navarro-Cerrillo, R. M., Abril, N., Jorrín-Novo, J. V., & Dodd, R. S. (2018). Population Genetic Diversity of Quercus ilex subsp. ballota (Desf.) Samp. Reveals Divergence in Recent and Evolutionary Migration Rates in the Spanish Dehesas. Forests, 9(6), 337. https://doi.org/10.3390/f9060337