Structural Attributes of Old-Growth and Partially Harvested Northern White-Cedar Stands in Northeastern North America


 ,
, 
Abstract
:1. Introduction
2. Methods
2.1. Study Site Selection
2.2. Site Description
2.3. Field Sampling and Calculations
2.4. Statistical Analysis
3. Results
Tree Species Composition and Forest Structure
4. Discussion
4.1. Tree Species Composition
4.2. Forest Structure
4.3. Management Implications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Forest Stewardship Council. FSC-US Forest Management Standard (v1.0); Forest Stewardship Council: Washington, DC, USA, 2010; Available online: https://us.fsc.org/preview.fsc-us-forest-management-standard-v1-0.a-188.pdf (accessed on 15 May 2016).
- McGee, G.G.; Leopold, D.J.; Nyland, R.D. Structural characteristics of old-growth, maturing, and partially cut northern hardwood forests. Ecol. Appl. 1999, 9, 1316–1329. [Google Scholar] [CrossRef]
- Whitman, A.A.; Hagan, J.M. An index to identify late-successional forest in temperate and boreal zones. For. Ecol. Manag. 2007, 246, 144–154. [Google Scholar] [CrossRef]
- Burrascano, S.; Keeton, W.S.; Sabatini, F.M.; Blasi, C. Commonality and variability in the structural attributes of moist temperate old-growth forests: A global review. For. Ecol. Manag. 2013, 291, 458–479. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Orwig, D.A.; Foster, D.R. The influence of successional processes and disturbance on the structure of Tsuga canadensis forests. Ecol. Appl. 2008, 18, 1182–1199. [Google Scholar] [CrossRef] [PubMed]
- Motta, R.; Garbarino, M.; Berretti, R.; Meloni, F.; Nosenzo, A.; Vacchiano, G. Development of old-growth characteristics in uneven-aged forests of the Italian Alps. Eur. J. For. Res. 2015, 134, 19–31. [Google Scholar] [CrossRef]
- Keeton, W.S. Managing for late-successional/old-growth characteristics in northern hardwood-conifer forests. For. Ecol. Manag. 2006, 235, 129–142. [Google Scholar] [CrossRef]
- Kimmins, J.P. Old-growth forest: An ancient and stable sylvan equilibrium, or a relatively transitory ecosystem condition that offers people a visual and emotional feast? Answer it depends. For. Chron. 2003, 79, 429–440. [Google Scholar] [CrossRef]
- Curtis, J.D. Preliminary observations on northern white-cedar in Maine. Ecology 1946, 27, 23–36. [Google Scholar] [CrossRef]
- Johnston, W. Thuja occidentalis L. Northern white-cedar. In Silvics of North America: Conifers; Burns, R., Honkalla, B., Eds.; USDA: Washington, DC, USA, 1990; Volume 1, pp. 1189–1209. ISBN 978-0160271458. [Google Scholar]
- Gawler, S.; Cutko, A. Natural Landscapes of Maine: A Guide to Natural Communities and Ecosystems; Maine Natural Areas Program, Maine Department of Conservation: Augusta, ME, USA, 2010; ISBN 978-0-615-34739-4.
- Fraver, S.; White, A.S.; Seymour, R.S. Natural disturbance in an old-growth landscape of northern Maine, USA. J. Ecol. 2009, 97, 289–298. [Google Scholar] [CrossRef]
- Larouche, C.; Kenefic, L.S.; Ruel, J.-C. Northern white-cedar regeneration dynamics on the Penobscot Experimental Forest in Maine: 40-year results. North. J. Appl. For. 2010, 27, 5–12. [Google Scholar]
- Selva, S.B. Using calicioid lichens and fungi to assess ecological continuity in the Acadian Forest Ecoregion of the Canadian Maritimes. For. Chron. 2003, 79, 550–558. [Google Scholar] [CrossRef]
- Tyrrell, L.E.; Nowacki, G.J.; Buckley, D.S.; Nauertz, E.A.; Niese, J.N.; Rollinger, J.L.; Zasada, J.C. Information about Old Growth for Selected Forest Type Groups in the Eastern United States; USDA Forest Service, General Technical Report NC-197; USDA: St. Paul, MN, USA, 1998.
- Hofmeyer, P.V.; Kenefic, L.S.; Seymour, R.S. Historical stem development of northern white-cedar (Thuja occidentalis L.). North. J. Appl. For. 2010, 27, 92–96. [Google Scholar]
- Frelich, L.E.; Reich, P.B. Perspectives on development of definitions and values related to old-growth forests. Environ. Rev. 2003, 11 (Suppl. S1), S9–S22. [Google Scholar] [CrossRef]
- Boulfroy, E.F.; Forget, E.; Hofmeyer, P.V.; Kenefic, L.S.; Larouche, C.; Lessard, G.; Lussier, J.-M.; Pinto, F.; Ruel, J.-C.; Weiskittel, A. Silvicultural Guide for Northern White-Cedar (Eastern White Cedar); USDA Forest Service, General Technical Report NRS-98; USDA: Newtown Square, PA, USA, 2012.
- PRISM Climate Group, Oregon State University. Available online: http://prism.oregonstate.edu (accessed on 6 June 2016).
- Web Soil Survey, Natural Resources Conservation Service, United States Department of Agriculture. Available online: http://websoilsurvey.nrcs.usda.gov/ (accessed on 6 June 2016).
- Brown, J.K. A planar intersect method for sampling fuel volume and surface area. For. Sci. 1971, 17, 96–102. [Google Scholar]
- Sollins, P.; Cline, S.P.; Verhoeven, T.; Sachs, D.; Spycher, G. Patterns of log decay in old-growth Douglas-fir forests. Can. J. For. Res. 1987, 17, 1585–1595. [Google Scholar] [CrossRef]
- Van Wagner, C.E. The line intersect method in forest fuel sampling. For. Sci. 1968, 14, 20–26. [Google Scholar]
- Fraver, S.; Milo, A.M.; Bradford, J.B.; D’Amato, A.W.; Kenefic, L.; Palik, B.J.; Woodall, C.W.; Brissette, J. Woody debris volume depletion through decay: Implications for biomass and carbon accounting. Ecosystems 2013, 16, 1262–1272. [Google Scholar] [CrossRef]
- Wesely, N.J. Old-Growth Characteristics of Northern White-Cedar Stands. Master’s Thesis, University of Maine, Orono, ME, USA, 2016. [Google Scholar]
- Pommerening, A. Approaches to quantifying forest structures. Forestry 2002, 75, 305–324. [Google Scholar] [CrossRef] [Green Version]
- Motz, K.; Sterba, H.; Pommerening, A. Sampling measures of tree diversity. For. Ecol. Manag. 2010, 260, 1985–1996. [Google Scholar] [CrossRef]
- Hui, G.; Zhao, X.; Zhao, Z.; von Gadow, K. Evaluating tree species spatial diversity based on neighborhood relationships. For. Sci. 2011, 57, 292–300. [Google Scholar]
- Peck, J.E.; Zenner, E.K.; Brang, P.; Zingg, A. Tree size distribution and abundance explain structural complexity differentially within stands of even-aged and uneven-aged structure types. Eur. J. For. Res. 2014, 133, 335–346. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities, 3rd ed.; MjM Software: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Genuer, R.; Poggi, J-M.; Tuleau-Malot, C. Variable Selection Using Random Forests. R Package Version 1.0.3. 2016. Available online: https://cran.r-project.org/web/packages/VSURF (accessed on 15 May 2016).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.r-project.org/ (accessed on 1 January 2016).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.1-8. 2015. Available online: http://cran.r-project.org/package=lme4 (accessed on 15 May 2016).
- Weiskittel, A.R.; Wagner, R.G.; Seymour, R.S. Refinement of the Forest Vegetation Simulator, Northeastern Variant growth and yield model: Phase 2. In 2010 Annual Report; Mercier, W.J., Nelson, A.S., Eds.; University of Maine, Cooperative Forestry Research Unit: Orono, ME, USA, 2010; pp. 23–30. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice-Hall, Inc.: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Wirth, C.; Messier, C.; Bergeron, Y.; Frank, D.; Fankhanel, A. Old-growth forest definitions: A pragmatic view. In Old-Growth Forests: Function, Fate, and Value; Wirth, C., Gleixner, G., Heiman, M., Eds.; Springer: Berlin, Germany, 2009; pp. 11–33. ISBN 978-3-540-92706-8. [Google Scholar]
- Mosseler, A.; Lynds, J.A.; Major, J.E. Old-growth forests of the Acadian Forest Region. Environ. Rev. 2003, 11, S47–S77. [Google Scholar] [CrossRef]
- Oliver, C.; Larson, B. Forest Stand Dynamics; John Wiley & Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Keyes, C.R.; Teraoka, E.K. Structure and composition of old-growth and unmanaged second-growth riparian forests at Redwood National Park, USA. Forests 2014, 5, 256–268. [Google Scholar] [CrossRef]
- Ruel, J.-C.; Lussier, J.-M.; Morissette, S.; Ricodeau, N. Growth response of northern white-cedar (Thuja occidentalis) to natural disturbances and partial cuts in mixedwood stands of Quebec, Canada. Forests 2014, 5, 1194–1211. [Google Scholar]
- Fraver, S.; White, A.S. Disturbance dynamics of old-growth Picea rubens forests of northern Maine. J. Veg. Sci. 2005, 16, 597–610. [Google Scholar] [CrossRef]
- Hanson, J.J.; Lorimer, C.G. Forest structure and light regimes following moderate wind storms: Implications for multi-cohort management. Ecol. Appl. 2007, 17, 1325–1340. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, P.; Briggs, S.V.; Ayers, D.A.; Doyle, S.; Seddon, J.; McElhinny, C.; Jones, N.; Sims, R.; Doody, J.S. Rapidly quantifying reference conditions in modified landscapes. Biol. Conserv. 2008, 141, 2483–2493. [Google Scholar] [CrossRef]
- Meyer, P.; Schmidt, M. Accumulation of dead wood in abandoned beech (Fagus sylvatica L.) forests in northwestern Germany. For. Ecol. Manag. 2011, 261, 342–352. [Google Scholar] [CrossRef]
- Liira, J.; Sepp, T. Indicators of structural and habitat natural quality in boreo-nemoral forests along the management gradient. Ann. Bot. Fenn. 2009, 46, 308–325. [Google Scholar] [CrossRef]
- Keeton, W.S.; Kraft, C.E.; Warren, D.R. Mature and old-growth riparian forests: Structure, dynamics, and effects on Adirondack stream habitats. Ecol. Appl. 2007, 17, 852–868. [Google Scholar] [CrossRef] [PubMed]
- Uotila, A.; Kouki, J.; Kontkanen, H.; Pulkkinen, P. Assessing the naturalness of boreal forests in eastern Fennoscandia. For. Ecol. Manag. 2002, 161, 257–277. [Google Scholar] [CrossRef]
- Di Filippo, A.; Biondi, F.; Piovesan, G.; Ziaco, E. Tree ring-based metrics for assessing old-growth forest naturalness. J. Appl. Ecol. 2017, 54, 737–749. [Google Scholar] [CrossRef]
- Winter, S.; Fischer, H.S.; Fischer, A. Relative quantitative reference approach for naturalness assessments of forests. For. Ecol. Manag. 2010, 259, 1624–1632. [Google Scholar] [CrossRef]
- Junninen, K.; Similä, M.; Kouki, J.; Kotiranta, H. Assemblages of wood-inhabiting fungi along the gradients of succession and naturalness in boreal pine-dominated forests in Fennoscandia. Ecography 2006, 29, 75–83. [Google Scholar] [CrossRef]
- Brumelis, G.; Jonsson, B.G.; Kouki, J.; Kuuluvainen, T.; Shorohova, E. Forest naturalness in northern Europe: Perspectives on processes, structures and species diversity. Silva Fenn. 2011, 45, 807–821. [Google Scholar] [CrossRef]
- Gradel, A.; Mühlenberg, M. Spatial characteristics of near-natural Mongolian forests at the southern edge of the taiga. Allg. Forst Jagdztg. 2011, 182, 40–52. [Google Scholar]
- Angers, V.A.; Messier, C.; Beaudet, M.; Leduc, A. Comparing composition and structure in old-growth and harvested (selection and diameter-limit cuts) northern hardwood stands of Quebec. For. Ecol. Manag. 2005, 217, 275–293. [Google Scholar] [CrossRef] [Green Version]
- Siitonen, J.; Martikainen, P.; Punttila, P.; Rauh, J. Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For. Ecol. Manag. 2000, 128, 211–225. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Lassauce, A.; Paillet, Y.; Jactel, H.; Bouget, C. Deadwood as a surrogate for forest biodiversity: Meta-analysis of correlations between deadwood volume and species richness of saproxylic organisms. Ecol. Indic. 2011, 11, 1027–1039. [Google Scholar] [CrossRef]
- Jönsson, M.T.; Fraver, S.; Jonsson, B.G. Forest history and the development of old-growth characteristics in fragmented boreal forests. J. Veg. Sci. 2009, 20, 91–106. [Google Scholar] [CrossRef]
- Morris, P.I.; Ingram, J.; Larkin, G.; Laks, P. Field tests of naturally durable species. For. Prod. J. 2011, 61, 344–351. [Google Scholar] [CrossRef]
- Ducey, M.; Gunn, J.; Whitman, A. Late-successional and old-growth forests in the Northeastern United States: Structure, dynamics, and prospects for restoration. Forests 2013, 4, 1055–1086. [Google Scholar] [CrossRef]
- Lee, P.; Hanus, S.; Grover, B. Criteria for estimating old growth in boreal mixedwoods from standard timber inventory data. For. Ecol. Manag. 2000, 129, 25–30. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F.; Franklin, J.F. Global decline in large old trees. Science 2012, 338, 1305–1306. [Google Scholar] [CrossRef] [PubMed]
- Goodburn, J.M.; Lorimer, C.G. Cavity trees and coarse woody debris in old-growth and managed northern hardwood forests in Wisconsin and Michigan. Can. J. For. Res. 1998, 28, 427–438. [Google Scholar] [CrossRef]
- McGee, G.G.; Kimmerer, R.W. Size of Acer saccharum hosts does not influence growth of mature bryophyte gametophytes in Adirondack northern hardwood forests. Bryologist 2004, 107, 302–311. [Google Scholar] [CrossRef]
- Zenner, E.K.; Peck, J.E. Characterizing structural conditions in mature managed red pine: Spatial dependency of metrics and adequacy of plot size. For. Ecol. Manag. 2009, 257, 311–320. [Google Scholar] [CrossRef]
- Kuehne, C.; Weiskittel, A.R.; Fraver, S.; Puettmann, K.J. Effects of thinning-induced changes in structural heterogeneity on growth, ingrowth, and mortality in secondary coastal Douglas-fir forests. Can. J. For. Res. 2015, 45, 1448–1461. [Google Scholar] [CrossRef]
- Donato, D.C.; Campbell, J.L.; Franklin, J.F. Multiple successional pathways and precocity in forest development: Can some forests be born complex? J. Veg. Sci. 2012, 23, 576–584. [Google Scholar] [CrossRef]
- Zhao, Z.; Hui, G.; Hu, Y.; Wang, H.; Zhang, G.; von Gadow, K. Testing the significance of different tree spatial distribution patterns based on the uniform angle index. Can. J. For. Res. 2014, 44, 1419–1425. [Google Scholar] [CrossRef]
- Fahey, R.T.; Fotis, A.T.; Woods, K.D. Quantifying canopy complexity and effects on productivity and resilience in late-successional hemlock-hardwood forests. Ecol. Appl. 2015, 25, 834–847. [Google Scholar] [CrossRef] [PubMed]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agric. For. Meterol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Cohen, W.B.; Acker, S.A.; Parker, G.G.; Spies, T.A.; Harding, D. Lidar remote sensing of the canopy structure and biophysical properties of Douglas-fir western hemlock forests. Remote Sens. Environ. 1999, 70, 339–361. [Google Scholar] [CrossRef]
- Eberhard, L.L.; Thomas, J.M. Designing environmental field studies. Ecol. Monogr. 1991, 61, 53–73. [Google Scholar] [CrossRef]
- Seymour, R.S.; Hunter, M.L., Jr. Principles of ecological forestry. In Managing Biodiversity in Forest Ecosystems; Hunter, M.L., Jr., Ed.; Cambridge University Press: Cambridge, UK, 1999; pp. 21–61. [Google Scholar]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, A.W.; Catanzaro, P.F.; Fletcher, L.S. Early regeneration and structural responses to patch selection and structural retention in second-growth northern hardwoods. For. Sci. 2015, 61, 183–189. [Google Scholar] [CrossRef]
- Cornett, M.W.; Reich, P.B.; Puettmann, K.J.; Frelich, L.E. Seedbed and moisture availability determine safe sites for early Thuja occidentalis (Cupressaceae) regeneration. Am. J. Bot. 2000, 87, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Verme, L.J.; Johnston, W.F. Regeneration of northern white cedar deeryards in upper Michigan. J. Wildl. Manag. 1986, 50, 307–313. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Stand Variable | Old-Growth (n = 16) | Partial Harvest (n = 17) | ||
|---|---|---|---|---|
| Mean (SD) | Range | Mean (SD) | Range | |
| Living trees (DBH ≥ 10 cm) | ||||
| Basal area (m2·ha−1) | 51.8 (17.9) | 26.6–94.2 | 54.5 (11.1) | 37.3–76.3 |
| Trees density (No. ha−1) | 765 (220) | 300–1130 | 1051 (98) | 560–1680 |
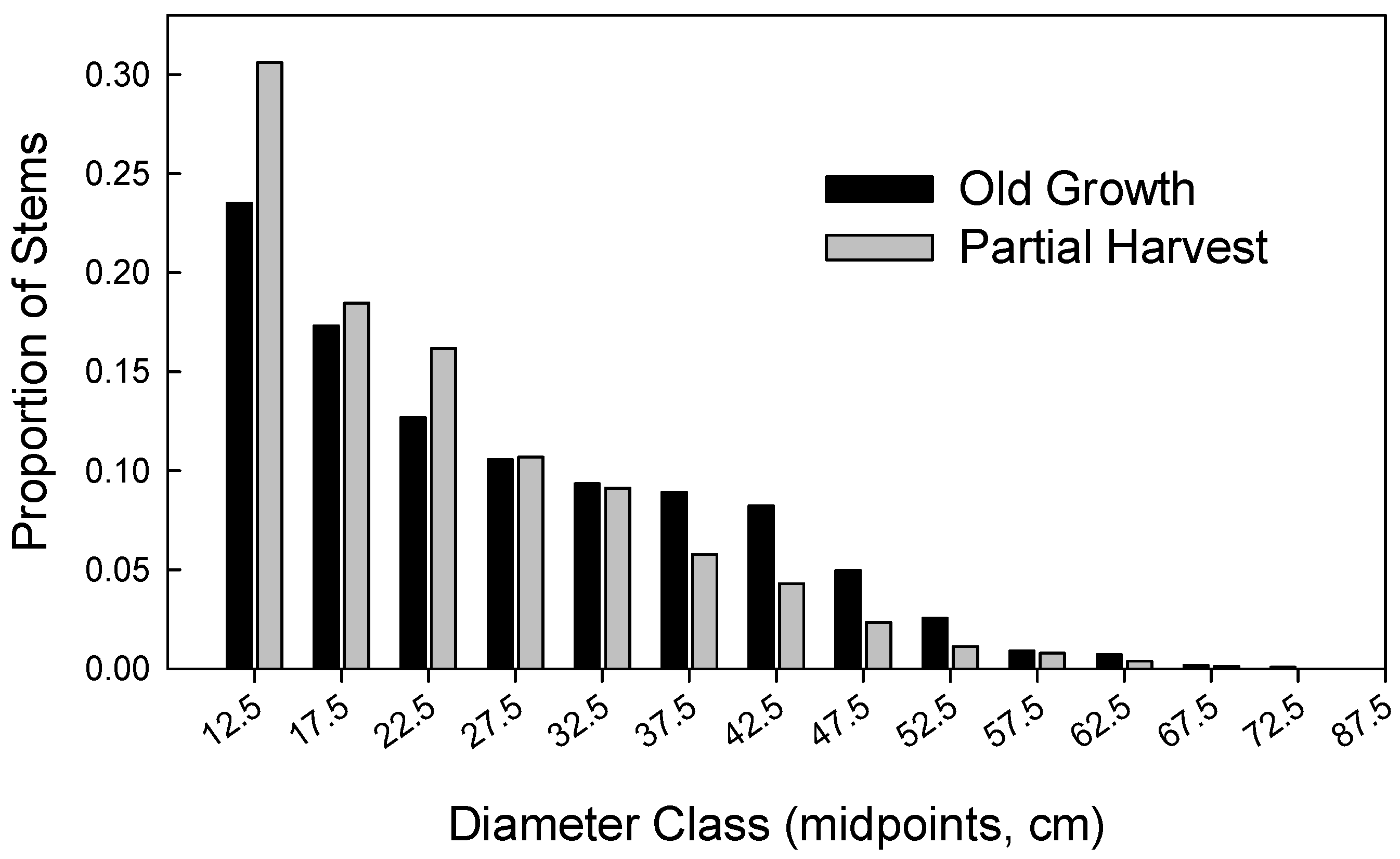
| Quadratic Mean Diameter (cm) | 29.4 (3.4) | 24.2–34.4 | 26.3 (4.8) | 19.8–34.9 |
| Large trees (≥40 cm DBH) (No. ha−1) | 135 (67) | 70–280 | 97 (68) | 0–210 |
| Standing dead trees (DBH ≥ 10 cm) | ||||
| Basal area (m2·ha−1) | 12.7 (10.0) | 3.2–31.9 | 7.5 (2.5) | 2.5–12.0 |
| Trees per ha (No. ha−1) | 164 (63) | 70–280 | 182 (79) | 90–350 |
| Quadratic Mean Diameter (cm) | 26.7 (4.4) | 18.7–36.2 | 23.3 (3.1) | 18.9–27.6 |
| Large trees (≥40 cm DBH) (No. ha−1) | 34 (42) | 0–110 | 10 (7) | 0–20 |
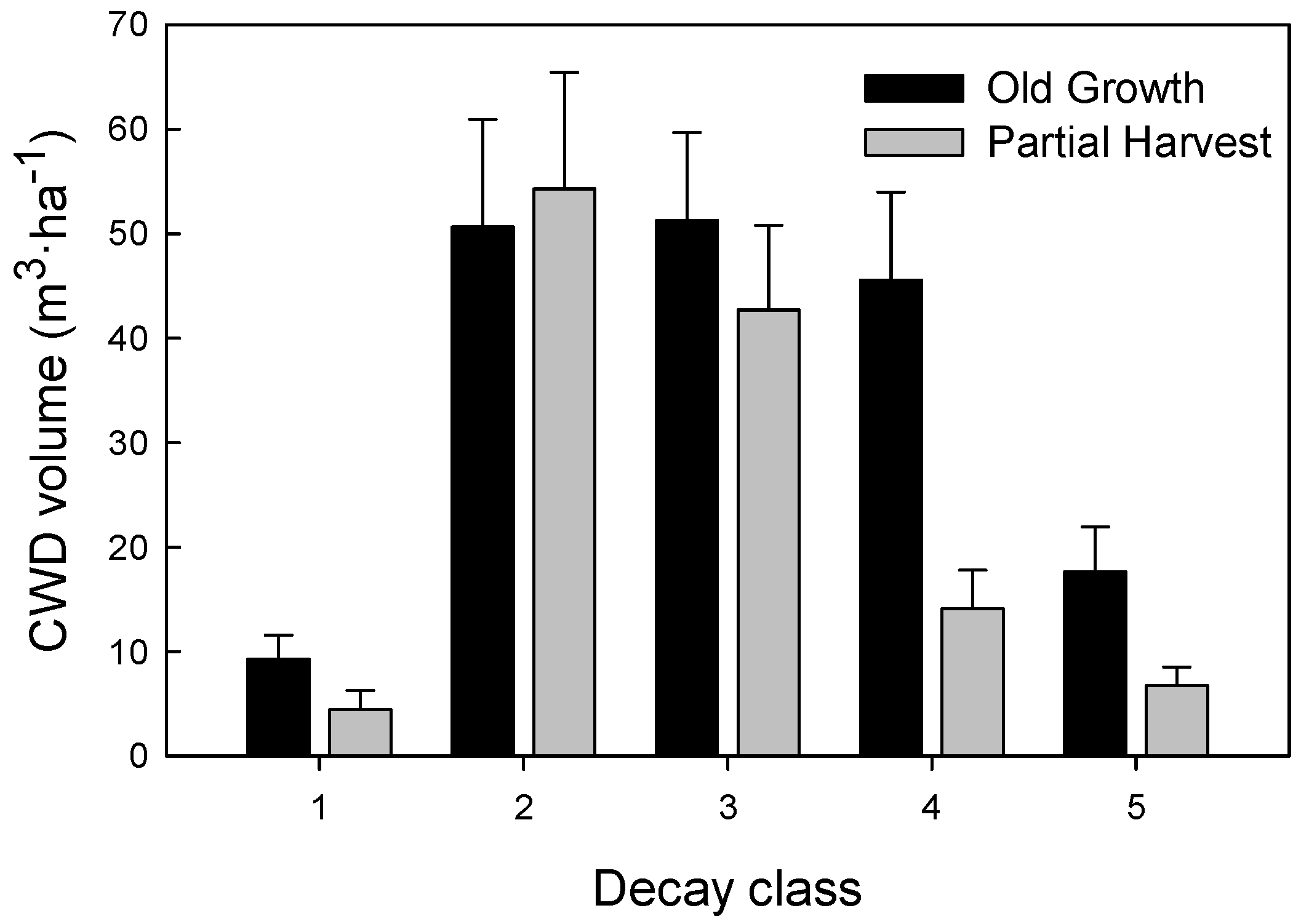
| Coarse woody debris (≥10 cm diam.) | ||||
| Total volume (m3·ha−1) | 168.1 (49.3) | 74.1–240.5 | 125.1 (80.2) | 36.4–314.1 |
| Advanced decay volume (m3·ha−1) | 60.6 (40.5) | 2.5–147.7 | 20.8 (21.1) | 0–70.2 |
| Cut stumps | ||||
| Density (No. ha−1) | 0 | 0 | 151 (89) | 30–320 |
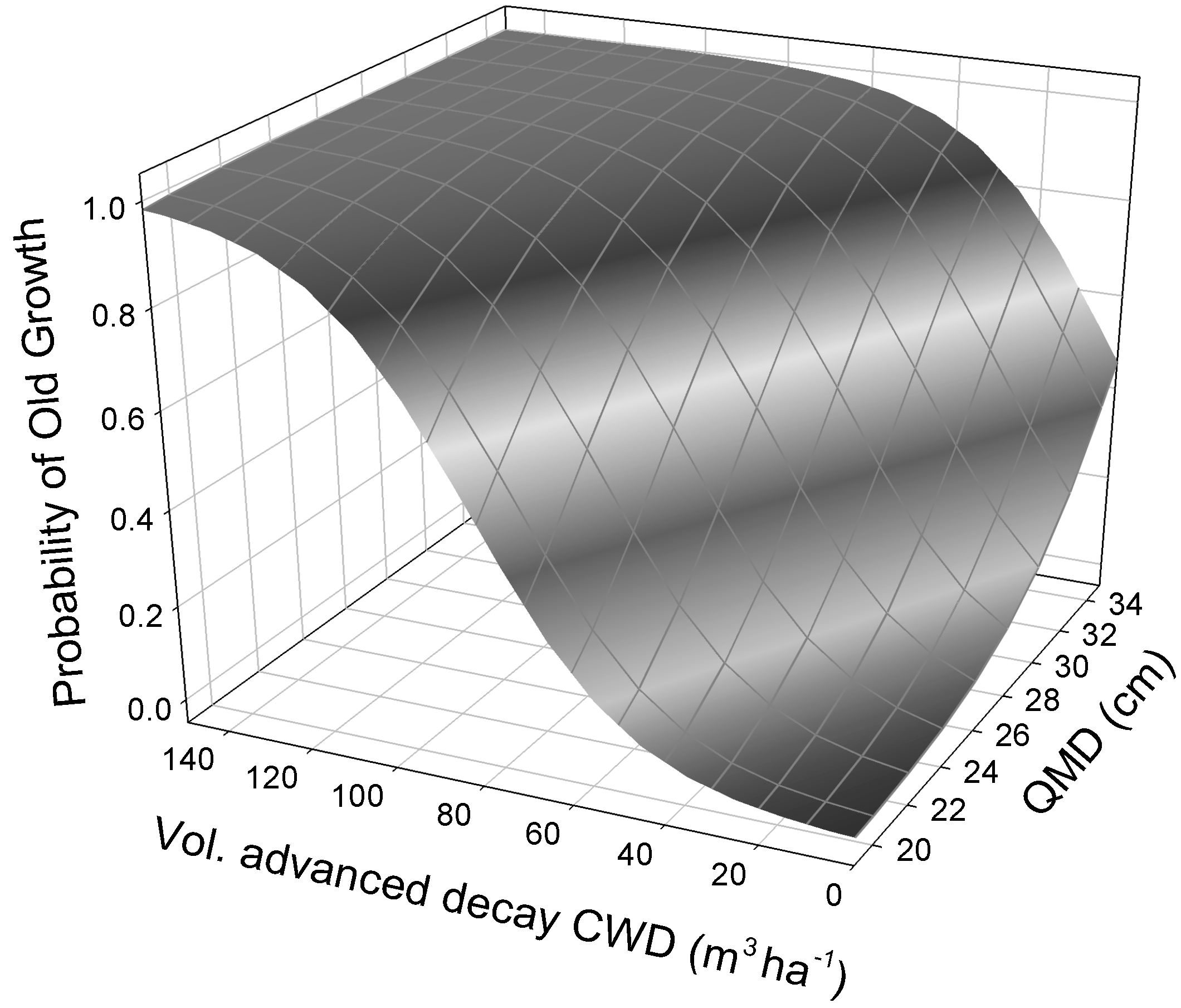
| Model Predictors | k | AICc | ΔAICc | AICc wt. | AUC | R2 |
|---|---|---|---|---|---|---|
| CWDADV * + QMD * | 4 | 37.8 | 0.0 | 0.62 | 0.84 | 0.69 |
| CWDADV * + QMD * + CWDTOT | 5 | 39.3 | 1.6 | 0.28 | 0.75 | 0.77 |
| CWDADV * + QMD + CWDTOT + TPH | 6 | 42.0 | 4.2 | 0.08 | 0.71 | 0.79 |
| CWDADV * + QMD + CWDTOT + TPH + SN_QMD | 7 | 45.2 | 7.5 | 0.01 | 0.70 | 0.79 |
| Variable | Parameter | Parameter Value (Standard Error) | p-Value |
|---|---|---|---|
| Intercept | a | −10.44 (4.60) | 0.023 |
| Advanced-decay CWD volume | b | 0.060 (0.02) | 0.013 |
| Quadratic mean diameter (QMD) | c | 0.294 (0.14) | 0.039 |
| Indices | Old-Growth (n = 16) | Partial Harvest (n = 17) | ||
|---|---|---|---|---|
| Mean (SD) | Range | Mean (SD) | Range | |
| Diameter differentiation | 0.35 (0.04) | 0.28–0.44 | 0.33 (0.03) | 0.27–0.39 |
| Mingling | 0.50 (0.19) | 0.18–0.77 | 0.43 (0.15) | 0.17–0.64 |
| Clark-Evans | 0.30 (0.02) | 0.27–0.34 | 0.29 (0.03) | 0.24–0.33 |
| Shannon-Weaver (diameters) | 1.08 (0.44) | 0.38–1.64 | 0.86 (0.30) | 0.40–1.35 |
| Mean directional | 1.86 (0.12) | 1.56–2.04 | 1.94 (0.09) | 1.78–2.15 |
| Gini (diameters) | 0.26 (0.03) | 0.22–0.29 | 0.24 (0.04) | 0.18–0.29 |
| Simpson (diameters) | 0.54 (0.20) | 0.19–0.77 | 0.45 (0.16) | 0.19–0.70 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wesely, N.; Fraver, S.; Kenefic, L.S.; Weiskittel, A.R.; Ruel, J.-C.; Thompson, M.E.; White, A.S. Structural Attributes of Old-Growth and Partially Harvested Northern White-Cedar Stands in Northeastern North America. Forests 2018, 9, 376. https://doi.org/10.3390/f9070376
Wesely N, Fraver S, Kenefic LS, Weiskittel AR, Ruel J-C, Thompson ME, White AS. Structural Attributes of Old-Growth and Partially Harvested Northern White-Cedar Stands in Northeastern North America. Forests. 2018; 9(7):376. https://doi.org/10.3390/f9070376
Chicago/Turabian StyleWesely, Nathan, Shawn Fraver, Laura S. Kenefic, Aaron R. Weiskittel, Jean-Claude Ruel, Michael E. Thompson, and Alan S. White. 2018. "Structural Attributes of Old-Growth and Partially Harvested Northern White-Cedar Stands in Northeastern North America" Forests 9, no. 7: 376. https://doi.org/10.3390/f9070376
APA StyleWesely, N., Fraver, S., Kenefic, L. S., Weiskittel, A. R., Ruel, J. -C., Thompson, M. E., & White, A. S. (2018). Structural Attributes of Old-Growth and Partially Harvested Northern White-Cedar Stands in Northeastern North America. Forests, 9(7), 376. https://doi.org/10.3390/f9070376