1. Introduction
High quality and good physiological condition of cutting material are key aspects of successful and economically feasible short rotation coppice (SRC) plantations [
1]. In addition to an insufficient availability of nutrients, unequal plant growth and dieback of cuttings are found to be the main factors resulting in a decline of production during consecutive SRC harvests [
2]. These kinds of problems are most severe in
Salix L. SRC where very high planting densities are usually suggested [
3]. In addition to economic considerations, SRC plantations with variable plant densities and large numbers of gaps in the SRC vegetation cover can cause ecological problems if the SRC is used as a phytoremediation site or in terms of vegetation filter [
4].
It is especially important to ensure fast, equal sprouting of cuttings with consecutive rapid shoot growth occur during the first growing season of SRC, since this is the period where the largest seed bank of weed species in the plantation soil exists and competition between sprouting cuttings and weeds can be expected [
5,
6,
7]. Therefore, a lot of attention has been paid to the study of critical factors that may impact the growth potential of cuttings [
8]. Besides the selection of planting material (highly productive genotypes and vigorous hybrid cultivars), the quality of cutting material survival has been found to be influenced by pre-planting storage conditions [
1], the quality of soil preparation [
9,
10] and planting techniques [
11]. There are also a number of publications that demonstrate the benefits of horizontal planting instead of the more typical vertical planting method and include discussion of the reasonable length of the cutting [
12,
13,
14]. Moreover, the impact of cutting diameter has been found to be an important predictor of plant growth and hence when mechanically planting using long rods, the effectiveness of placing the thinnest top of one rod adjacent to the thickest bottom of another rod is considered to be arguable [
8]. The potential for decreasing the typical cutting length from between 18 and 25 cm to between 1 and 5 cm (micro-cuttings) in order to minimize plantation costs has also been studied [
15].
Another approach to enhancing the economic feasibility and sustainability of modern agriculture including SRC management is to promote the distribution of soil microorganisms (fungi or bacteria) that support nutrient uptake from growing substrate and thus enable a reduction in the use of mineral fertilizers [
16]. The rhizosphere microbiome is considered to be dependent on root exudates and to be species-specific. Some studies have revealed that
Salix can take advantage of specific fungal assemblages [
17,
18]. It has been demonstrated that planting of
Salix in former permanent grassland areas resulted with differences in the soil microbial community [
19]. Moreover, there is strong evidence that
Salix also has some endophytes in terms of symbionts for nitrogen uptake [
20], hence, the inoculation of planting material with specific microorganisms is suggested [
3].
There is information available about the physiological responses of different willow genotypes to variable water and nutrient conditions in pot experiments (e.g., [
21]). During the first growing season in some genotypes, even a short one-week period of water stress can lead to a significant decrease in plant biomass and height [
22]. Supplementation of nutrients in terms of mineral fertilizer or various organic residues from human activities (e.g., sewage sludge, wastewater, biogas digestate, etc.) may increase the biomass increment of young plants significantly [
23,
24]. Differences in biomass production can be observed only in cases where large differences exist between the “control” growing substrate and artificially created altered growing conditions [
4]. The maximal fertilization, however, must be carefully considered, since young cuttings cannot withstand such large doses of additional nutrients or utilize residues in the same way that mature SRC plants do [
23]. The duration of such greenhouse or pot-experiments however, is usually limited by the size of a growing area or difficulties in maintaining homogenous artificial growing conditions.
There are greenhouse data available on the leaf phenology of several
Salix genotypes that can be correlated with the wood yield of particular clones in SRC [
25,
26], little less is known about cutting biomass production in the absence of additional nutrients or growth substrates. Willow is a species that is easy to propagate vegetatively by chopping dormant stems into cuttings. New roots on the cutting sprout from latent root primordia that have been formed during stem development and are ready to grow as soon as the cutting is placed in a humid environment [
27]. However, studies have shown that
Salix species exhibit great variability in bud structure and therefore coppicing ability [
28]. Hence the rapid visual evaluation of bud number can be a useful indicator for predicting the early stage biomass production of cuttings.
While hydroponic experiments have been undertaken to study plant tolerance to a range of chemicals or to gather information about the synthesis of root exudates [
29,
30], information from hydroponic experiments for selecting energy crops, or even SRC planting material is very limited. However, this can add valuable information about particular clone preferences and shortages. Therefore, we decided to undertake a repeated experiment with the main aim of gaining an understanding of whether a quick, cheap, and simple to use method can be reasonably employed to obtain additional knowledge on those
Salix genotypes commonly grown in SRC.
The experiment was designed to test the following hypotheses:
- -
cutting genotype and diameter is a predictor of early stage biomass
- -
application of fertilizer and/or microorganisms originating from mature SRC promote the early stage biomass production of cuttings
- -
there are significant differences in the sprouting speed and early stage biomass production between different willow clones
- -
allocation of photosynthesis products between shoots and roots varies by clones and/or by applied substances
- -
early stage biomass production of a cutting can be predicted by counting the number of visible buds on the cutting.
2. Materials and Methods
The shoots of the willow clones studied, were harvested from the Kambja vegetation filter, South-Estonia (58°14′57′′ N, 26°42′39′′ E) on 15 April 2009 and 15 April 2010. The harvesting time was selected to avoid snow cover in SRC and start the experiment before the beginning of the growing period or any leaf bud burst in Estonian climatic conditions. In the first year, shoots of clones “Tora” and 78,183 (according to the Swedish clone numbering system) were studied. In the second year, material from clones 81,090 and “Gudrun” was added to the above clones in the study. Hence the clone selection represented both clones of Salix viminalis L. (78,183), S. dasyclados Wimm. (81,090) and the relatively new commercial hybrids of S. dasyclados “Helga” burjatica × S. viminalis (clone “Gudrun”) and S. schwerinii E. Wolf × S. viminalis (clone “Tora”). In the field the tallest (1.6–2.5 m) one-year-old, straight shoots from inside the plantation were selected to harvest the rods and cut at 5–10 cm height above the ground. Within a few hours the collected material was transported to the laboratory for further experiment preparation the following day.
The experiment (between 16 April and 14 May both in 2009 and 2010) was carried out in a laboratory of the Estonian University of Life Sciences at a room temperature of between 18–23 °C. The experiment room had large windows facing in a southerly direction and the natural light on windowsills was considered to be equal for all studied materials. At the start of the experiment the rods were divided into cuttings of 20 ± 0.1 cm in length as long as their diameter was more than 7 mm and their location along the rod in terms of position from the ground (1–8), number of visible buds on the bark and diameters of the apical cutting part were recorded and the cutting was marked using a waterproof pencil. In total, 120 cuttings per clone in 2009 and 60 cuttings in 2010 were made. After this procedure, 15 cuttings originating from the same clone were placed in a similar orientation vertically into a specially designed aquarium-like glass box of 8 × 40 × 25 cm and placed on the windowsill to get the maximum natural light available. The open glass boxes with different clones were located on the windowsills alternately and filled to a 15 cm height with water which originated from an open well near to the Kambja SRC that met all the criteria for drinking water. Since the evaporation from the boxes was small and no significant algae intervention occurred, we did not add any water or disturb the glass boxes in any other way during the experiment. The cuttings were placed along the longer edge (40 cm) of the box and hence the distance between cuttings was more than 2 cm to avoid any competition for space between young plants during the experiment (
Figure S1).
In 2009, every second glass box received 150 g of soil collected from the upper 10 cm of the Kambja vegetation filter control plots (sandy loam soil with pH 5.5) or/and 10 g of typical solid NPK (nitrogen, phosphorus and potassium) fertilizer (17.0:3.1:11.6) was added to the ca 4.8 L of water in the boxes, thus creating four different treatment options (control, fertilizer, soil, and fertilizer + soil) (
Table 1,
Figure S2). The application of soil decreased water transparency significantly. In 2010, only the addition of mineral fertilizer was repeated. Since the space with homogeneous natural light conditions was limited, this change allowed us to keep the number of cuttings in one treatment constant (
n = 30 replicated into two boxes) but to increase the number of clones studied in 2010.
Two weeks after the start of the experiment (30.04 in both years), each second cutting from each box/treatment/replication box was harvested. The cutting was taken out of the box and all sprouted shoots and roots were carefully removed from the cutting with the help of scissors or a razor blade. All harvested biomass was separately collected into paper envelopes and dried for at least 48 h at 85 °C to constant weight. After four weeks of the experiment, the same procedure was carried out with the remainder of the cuttings.
For statistical analyses the software package SAS System v. 9.4 (SAS Institute Inc., Cary, NC, USA) was used. In all cases, General Linear Models were exploited, and the significance of all factors was tested with SAS GLM (generalized linear model). The regressions between cutting diameter and biomass production were found with Linear Regression Model, the differences between average values of various datasets were detected with ANOVA or MANOVA procedures Ryan-Einot-Gabriel-Welsh Multiple Range Test (REGW) and the covariation of different factor combinations analyzed with the Least Square Means test (LSM). In all cases the confidence level of tests was set to 0.05.
From previous studies we assumed that there is a strong linear correlation between the cutting diameter and its biomass [
8]. In our experiment the studied shoots were younger and the range of diameters in each data series was variable. Therefore, we found it useful to normalize the biomass data over the diameter values and used the equation of the GLM linear regression fit for all collected data (Equation (1)):
where biomass production is the measured dry biomass (g) of a particular cutting and diameter is the measured upper diameter of the same cutting in the start date of the experiment.
The slope coefficient from this equation (a), further labelled as ”relative biomass production“, was calculated separately for each cutting data and used as the dependent variable instead of the actual biomass production data to enable a more detailed statistical analysis on a range of experiment factors. In practice this slope indicates the influence of cutting diameter (mm) on the production of new biomass (g) from the cutting.
3. Results
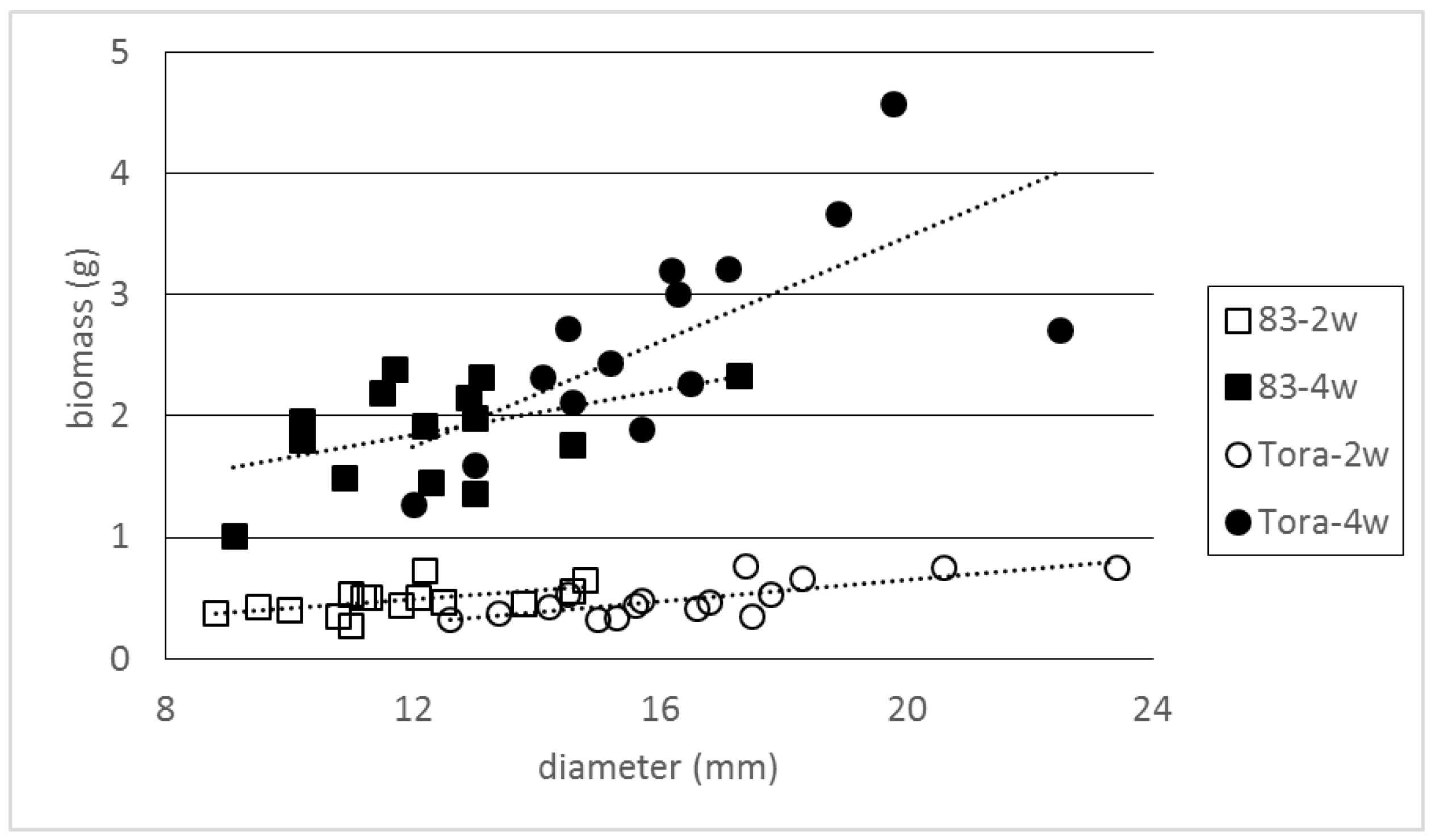
The general model, that included all experiment data, revealed that the total new biomass production of cuttings was significantly dependent on study year, on treatment, on clone and on cutting diameter, but the replication did not add any significant information to the model (
Table 2). Hence, further normalization was necessary as the range of cutting diameters under investigation varied by clones (e.g.,
Figure 1). The intercept value of the general linear regression model (
R2 = 0.83
p < 0.001) Equation (1) was −0.270173.
The 2009 data show that the relative biomass production was significantly (
p < 0.05) affected by harvesting date, treatment and clone. The other factors studied (the replication and the cutting position) were found to be insignificant and removed from further analyses. The detailed analysis was carried out separately for both harvesting dates (
Table 3). In both cases the treatment and clone were significant factors having an impact on relative biomass production. The interaction between these two factors was found to be insignificant in most cases and hence we assumed that cuttings from both clones reacted to the treatment in a similar way.
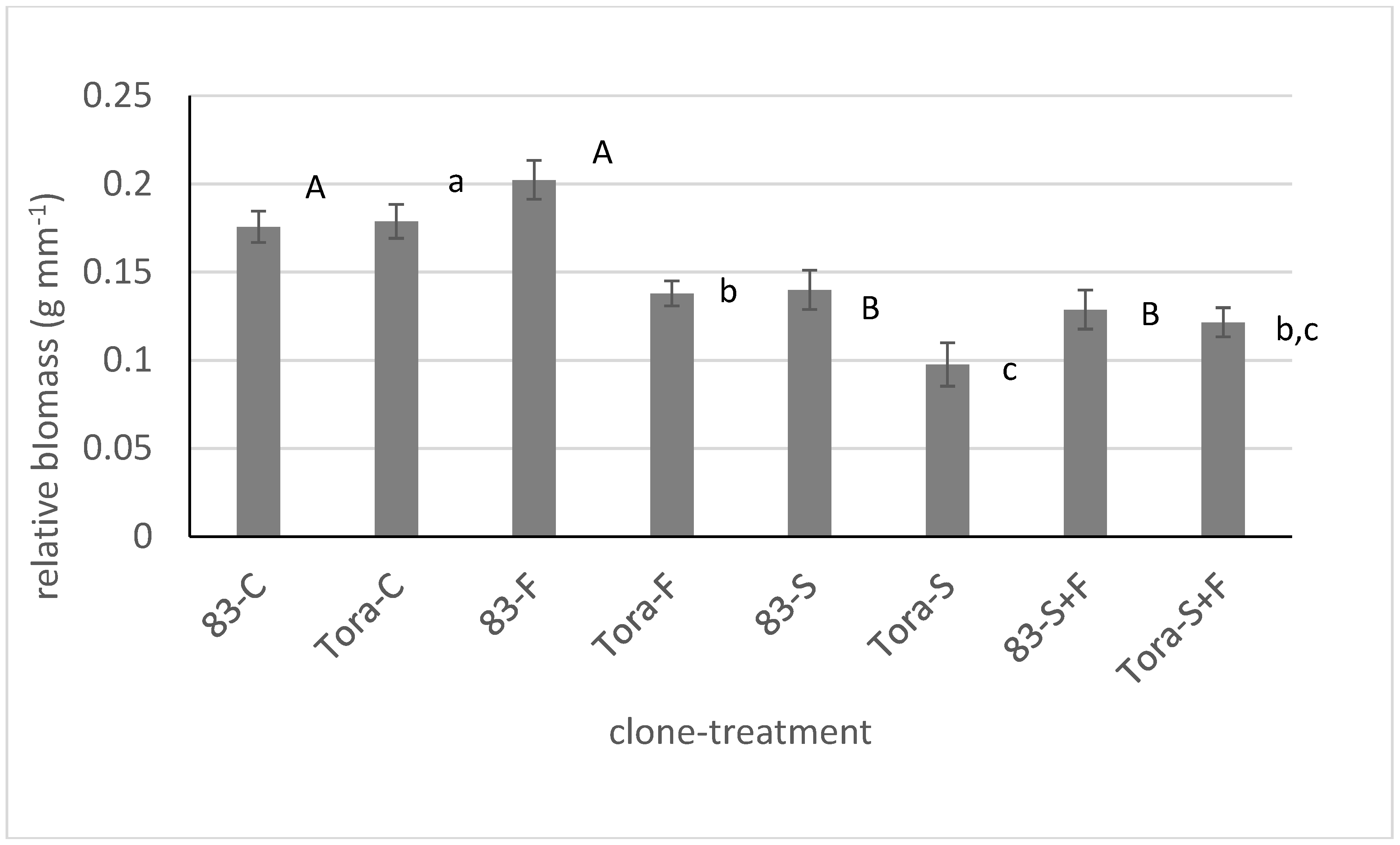
The statistical analyses of pooled data from both clones revealed that the application of different materials to the water boxes did not have any positive effect on the biomass production of the cuttings after four weeks of treatment when compared to the control dataset in 2009 (
Figure 2). In contrast, soil (S) supply decreased the production alone and in combination with fertilizer (S+F) both when the biomass production was measured after two weeks or after four weeks from the start of the experiment. At the end of the experiment a negative impact of the fertilizer application was also detected on clone 83, but this pattern was not found to be statistically significant, when the clone factor was excluded from the analysis.
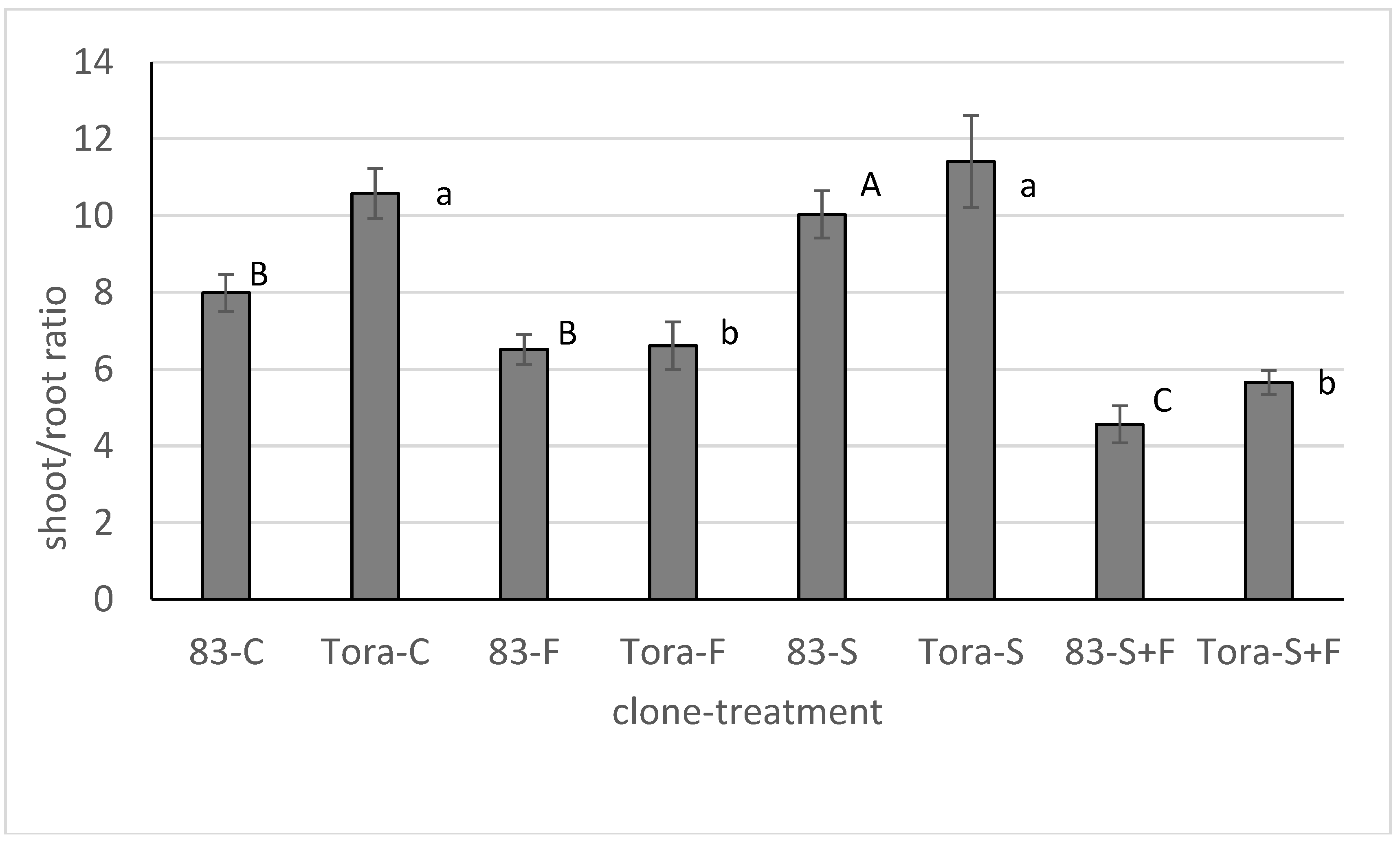
In order to better understand the impact of the treatments on cutting biomass production we also studied the ratio between shoot and root biomass production. At the end of the experiment the cuttings which were treated with fertilizer alone or in combination with soil tended to have a smaller shoot to root ratio than cuttings from other treatments (
Figure 3).
The variability of relative biomass production by clones was studied in detail on the basis of the data from 2010. In this year the covariation of clone and treatment factors were insignificant both in two-week harvest and four-week harvest data (
Table 4). These results enabled pooling of the data excluding the fertilization factor during this year and to increase the sample number for further analysis. We found strong evidence that the relative biomass production was clone-dependent. Biomass production for all the clones studied was approximately the same during the first two weeks and the final two weeks of the experiment. Clone 90 had significantly lower relative biomass production per cutting diameter than the rest of the clones studied for both the two-week and four-week harvest dates (
Figure 4).
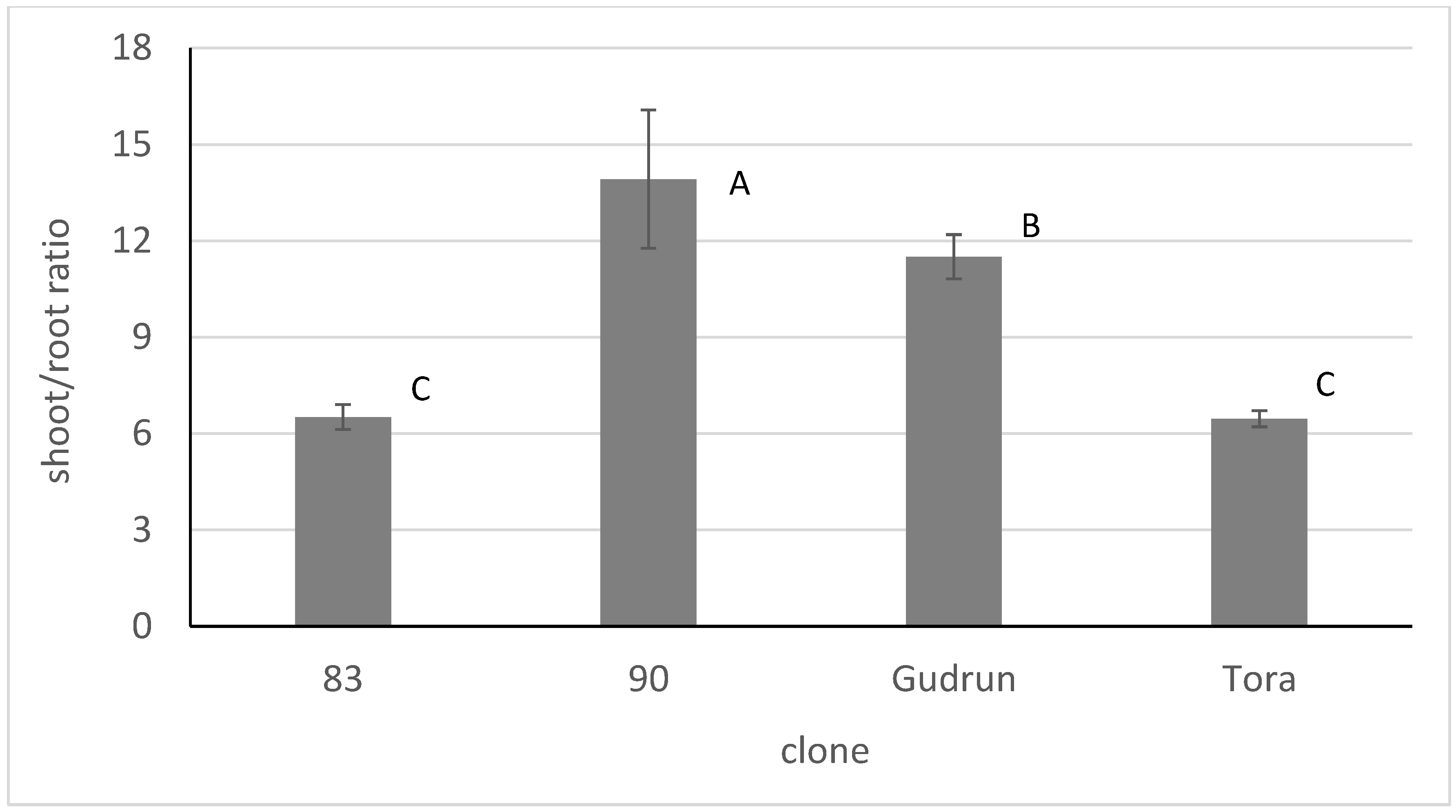
The lower relative biomass growth of clone 90 was accompanied by the smaller biomass of sprouted roots: The shoot/root biomass ratio of this clone was significantly larger than that of other clones (
Figure 5). Clone Gudrun also had a larger shoot/root production compared with clones 83 and Tora.
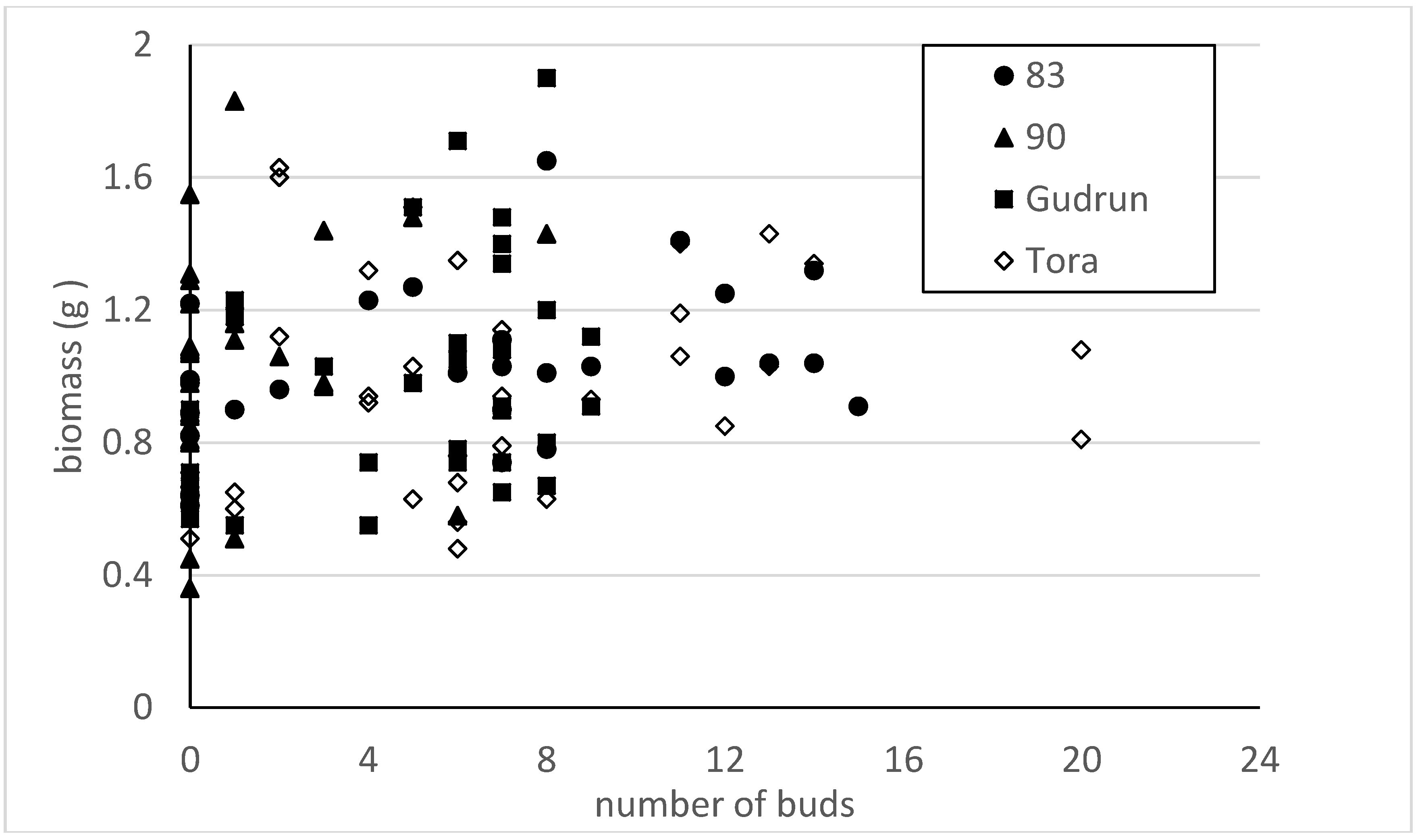
Analysis of the 2010 data also shows that the number of visible buds on cuttings was an unreliable parameter for predicting absolute biomass production when harvested after the first four weeks. No significant relationship was found between the number of visible buds and cutting biomass production by clones (
R2 < 0.15 in all cases) either (
Figure 6). However, the General Linear Model revealed that both the cutting diameter and clone were significant factors for predicting the number of buds on the cutting (
p < 0.0001 and
p < 0.05, respectively), but the model reliability was rather low (
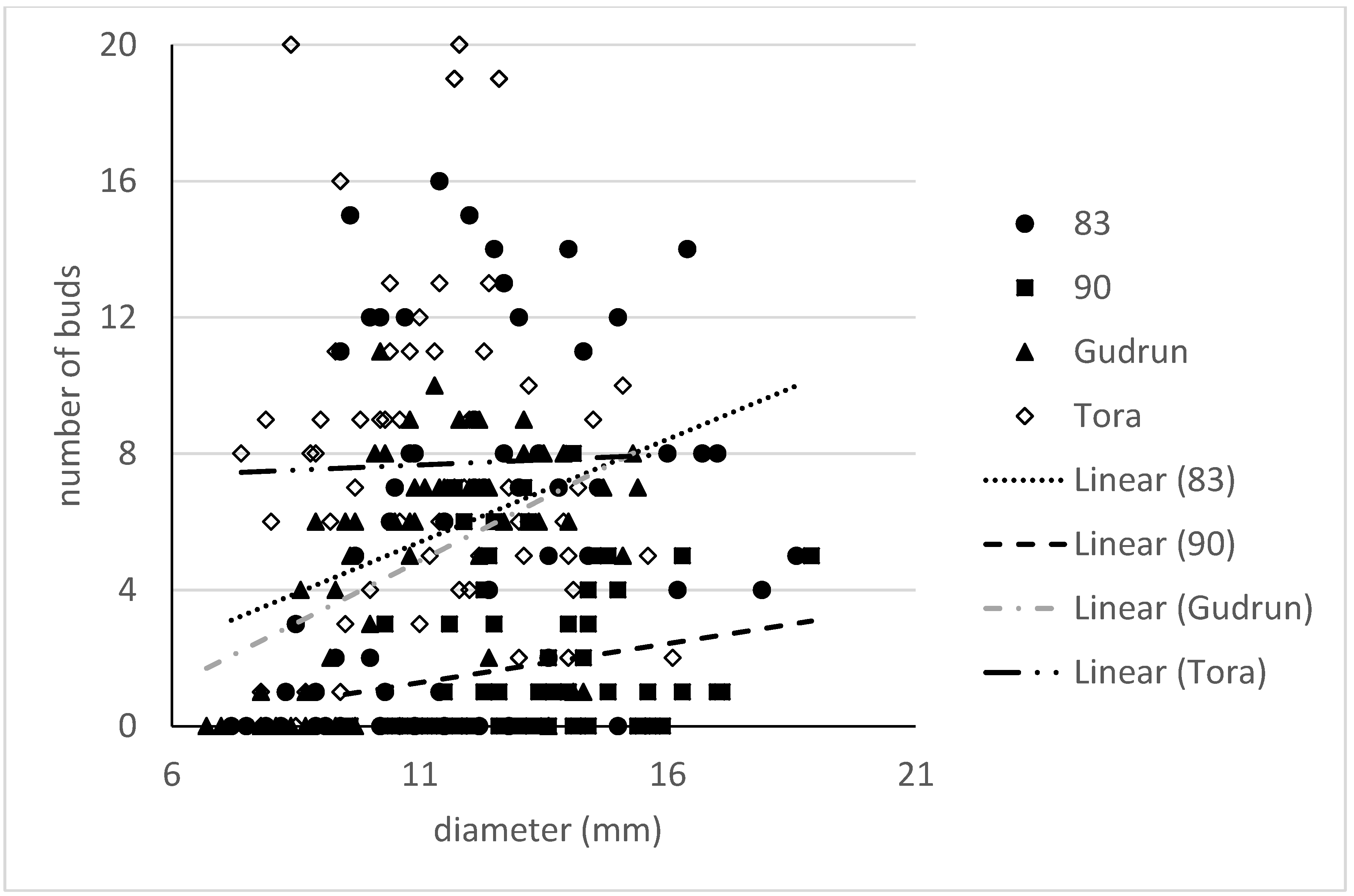
R2 = 0.29). Inclusion of data about the origin of the cutting from the donor rod did not improve the model neither was this factor significant. The detailed analysis demonstrated that the pattern between cutting diameter and number of visible buds varied by clones (
Figure 7). For clone 83 there was a tendency to positive correlation (
R2 = 0.10) between cutting diameter and the number of buds. At the same time clone Tora tended to have a similar number of buds regardless of the cutting diameter. In many cases no visible buds were detected on cuttings and it was especially difficult to detect buds in clone 90, where 28 cuttings out of 60 were marked with 0 visible buds. The number of cuttings from clone Tora with no visible buds was tenfold smaller than that of clone 90 in our experiment. Hence, the average numbers per cutting were 7.7; 6.0; 5.0 and 1.8 for Tora, 83, Gudrun and 90, respectively.
4. Discussion
Our lab-scale experiment with Salix cuttings revealed that there are some patterns in the biomass production and allocation between roots and shoots that can be linked with studied factors. In general, hydroponic conditions differ from the natural growing environment by both unlimited water supply and more anaerobic conditions of rhizosphere. However, to our knowledge the studied Salix species in nature prefer wet growing conditions. Therefore we assume that both their roots and potential symbiotic microorganisms behaved as usual in our laboratory conditions. In this case our study results do not contain only purely new scientific information but can be extrapolated to the practical SRC establishment as well.
The strong linear regression between cutting diameter and the biomass of newly formed roots and shoots detected in these experiments has also been reported by other scientists [
8,
31]. This phenomenon is most probably caused by the larger volume of products from photosynthesis stored in cuttings of a greater diameter, thus larger volumes of energy were available for new biomass growth. The intercept reciprocal value of the regression model (−0.270173 in our case) indicates the minimal hypothetical value of the cutting diameter that is still able to produce new biomass. The crucial numeric value of this parameter for each clone that marks the lower diameter limit for vigorous cuttings which are capable of sprouting can be calculated by dividing this constant with the relative biomass production value for particular data series. These critical diameters are much lower than the cutting diameters that are currently used for practical SRC establishment in order to avoid cutting debarking or breaking. However, this information can contain useful additional information for planning manual planting, operating with micro-cuttings or assuming the reasonable rod length for horizontal planting of
Salix shoots [
14].
Differences in the available cutting diameter range for various clones was the reason for using the normalized biomass production number in our analysis instead of absolute values. This correction evened out the differences in the cutting biomass production of clones 93 and Tora in 2009. Moreover, this analysis gave a negative result to our hypothesis, that the original position of cuttings on the rod may have an impact on the biomass production of cuttings of the same diameter. Cuttings with a given diameter from apical parts of rods are reported to have a larger biomass production than cuttings originating from basal parts, but this result was not confirmed by our study with other Salix clones.
The lack of statistical relevance for replication/box number in our general model revealed that both boxes with particular clone/treatment cuttings were located in equal environmental conditions in the laboratory. Differences in both absolute and relative biomass production by study years are most probably caused by differences in the available direct sunlight by years that could also have a slight impact on the water temperature. After four weeks of experiment in 2010 both Tora and 83 control series produced significantly less biomass per cutting diameter than in 2009. According to the Estonian Weather Service database (I.Saaremäe, personal communication) in the closest weather station to our experiment location there was 15% less direct sunshine in April 2010 than in the previous year. Hence, we conclude that light conditions during sprouting are important and the results obtained in different weather conditions cannot be compared in detail.
We did not detect any leaf discoloration or any other symptom of nutrient deficit during the experiment even in the control treatment without any nutrient supply. Application of soil originating from the local SRC had no positive effect on the relative biomass production of cuttings both alone and in combination with fertilizer. There can be two explanations for such a result. First, we cannot be sure if we got sufficient useful microorganisms from the SRC soil to promote
Salix growth or if they were able to survive in anaerobic hydroponic conditions. Second, it is possible that in the sprouting stage no support from microorganisms is available or required. Even more interesting however, is that the harvested biomass of soil-treated cuttings was significantly lower both at two and four weeks from the experiment start and with the ratio of shoots and roots biomass remaining similar with the control series. Since all growing conditions, with the exclusion of water transparency, remained the same, we have to assume that cutting bark is sensitive to light capture. As this phenomenon appeared throughout the experiment, light availability does not only trigger cuttings’ sprouting but most probably also has a positive impact on biomass production without changing the resource allocation between roots and shoots. The phenomenon of carbohydrate refixation by plant’s green parts, other than leaves, is reported to occur in
Populus tremula L. [
32]. This hypothesis is also supported by the report by S. Edelfeldt et al that shoots emerge from the ground more quickly from shallower horizontally planted cuttings than those from deeper planted [
12], as in this case some differences in the light availability can also be expected.
For practical SRC establishment it is usually recommended that cuttings are planted in the ground with no more than 1–2 cm of soil on top to avoid drought impacting cuttings before root formation [
33]. However, if light availability is important to promoting the speed of cutting biomass production in both above and below ground parts, it is perhaps useful to cover the upper part with a material which could avoid evaporation yet allow light penetration to promote fast sprouting of cuttings followed by rapid growth and thus avoid competition with the weeds.
Application of the fertilizer in 2009 did not have any significant impact on cuttings’ relative biomass production and changes were noticeable with only one clone. Repeating the experiment with the same fertilizer a year later did not confirm any significant differences either and therefore we conclude that additional nutrient availability is not important for cutting biomass production at this stage of growth or the fertilizer concentration in our experiments was too low to have any impact on the cutting productivity. However, in 2009 we found that newly formed biomass in treatments with fertilizer tended to be allocated towards the roots. This indicates that the cuttings were sensitive to nutrients in the hydroponic conditions and allocated their resources to get access to them. Such a plant strategy may be disadvantageous for practical SRC establishment into fertile soil. In this case the young Salix plants that allocate more of their resources to root production and less to shoot production suffer more from competition for light with weeds that have an opposite strategy.
It has been claimed that some of the new commercialized
Salix clones have an earlier bud burst than others [
8,
25]. Our experiment in 2010 which included four
Salix clones did not reveal any differences in cutting sprouting speed that could be detected by the biomass production during the first two weeks of the growing period. Biomass production during the final two weeks of the experiment was almost the same as that of the first two weeks, indicating similar growing speeds for all studied clones in the early growing stage. Clone 90 (
S. dasyclados), had the lowest relative biomass production at the two-week stage and its cuttings also performed the poorest at the end of the experiment. This result concurs with our data from the cuttings original SRC where clone 90 also had the lowest biomass production per plant during the first rotation cycle [
34]. This confirms the statement from our Swedish colleagues that pot-experiment results enable the prediction of performance of the plants in the field [
35]. Clone 90 also had a significantly higher shoot/root production ratio than the other clones. Higher shoot production compared to root production than in the rest of the clones studied, was also detected in clone Gudrun. This hybrid clone also has some
S dasyclados genes from its ancestors and therefore species-specific strategy can be speculated. The probability of gaining an advantage in practice from this strategy as well as from any other pattern described in the current experiment depends on the abiotic conditions in SRC. The most crucial factor here is a sufficient water supply that allows the cuttings to use their full potential and suppress the other stress factors. Moreover, the biomass production of cuttings is not a linear function in time and the first produced grams of biomass cannot be multiplied over the SRC lifetime to predict the wood yield.
Visible number of buds on a cutting was in correlation with the cutting diameter and clone but the statistical model only explained a relatively small proportion of the variability. There was strong evidence that the number of buds is larger on thicker cuttings. However, on clone Tora the number of visible buds was similar despite the cutting diameter. Although quite a number of cuttings were marked with no buds, they performed well in our biomass production experiment. The number of such cuttings was especially large among clone 90 cuttings. Hence, we must conclude that it is difficult to accurately measure the bud number on shoots in the dormant stage and that perhaps bud counting should take place after the cuttings have been in hydroponic conditions for some time and the vigorous buds are better detectable. However, such a technique would not be suitable as an express method in the field. Moreover, we are not sure that the counting of buds enables us to predict total biomass production since the relationship between the counted visible number of buds and absolute biomass production without any normalization with cutting diameter does not demonstrate any promising tendencies. Additionally, horizontal planting density must be based on other cutting characters than the number of visible buds on the rod. Most probably, during vegetative propagation the availability of other plant tissues including latent root primordia is more important for sprout formation than that of buds which are primarily grown to form new leaves in the future.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}