Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Analysis
3. Results
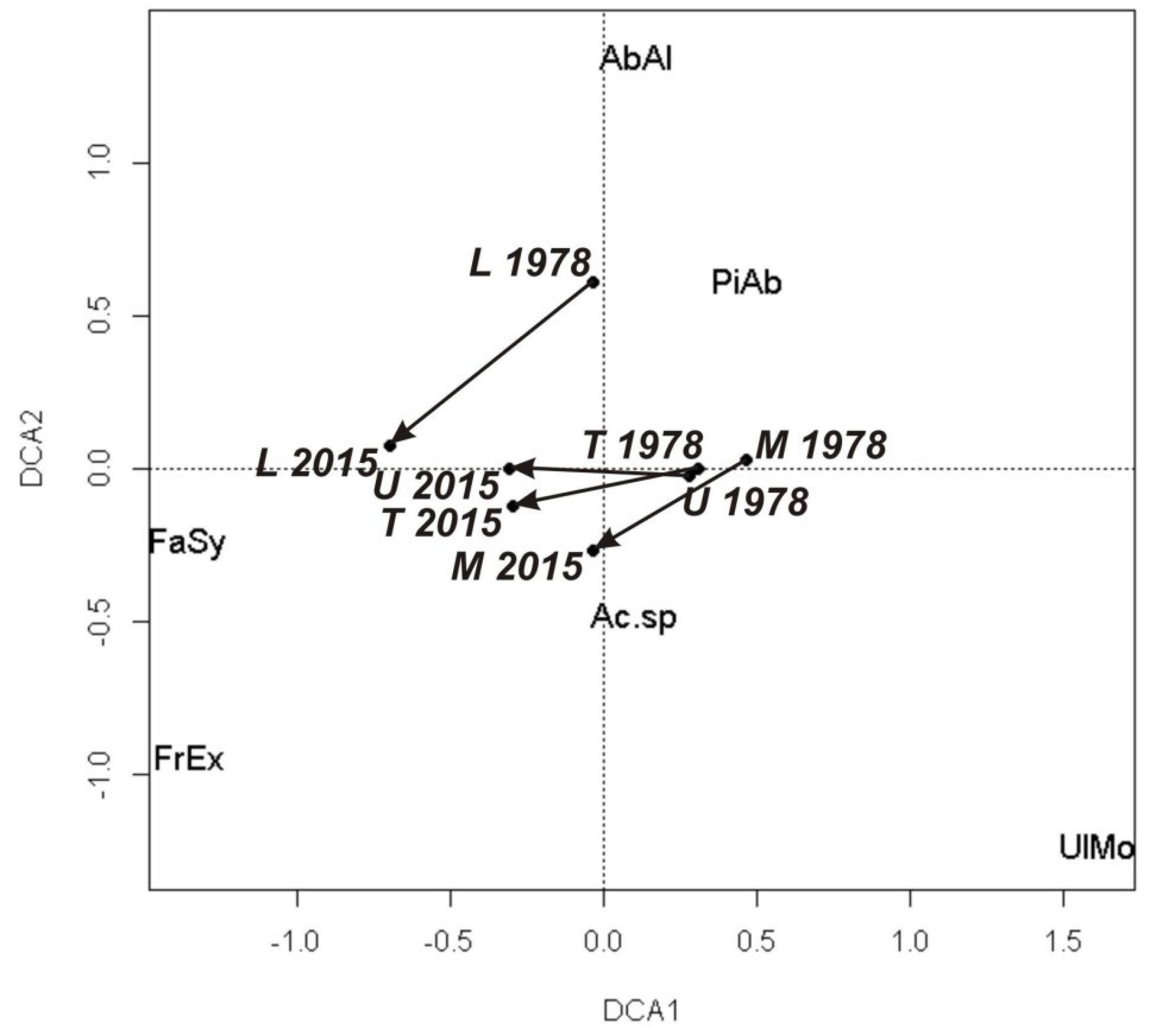
3.1. Changes in Tree Species Composition
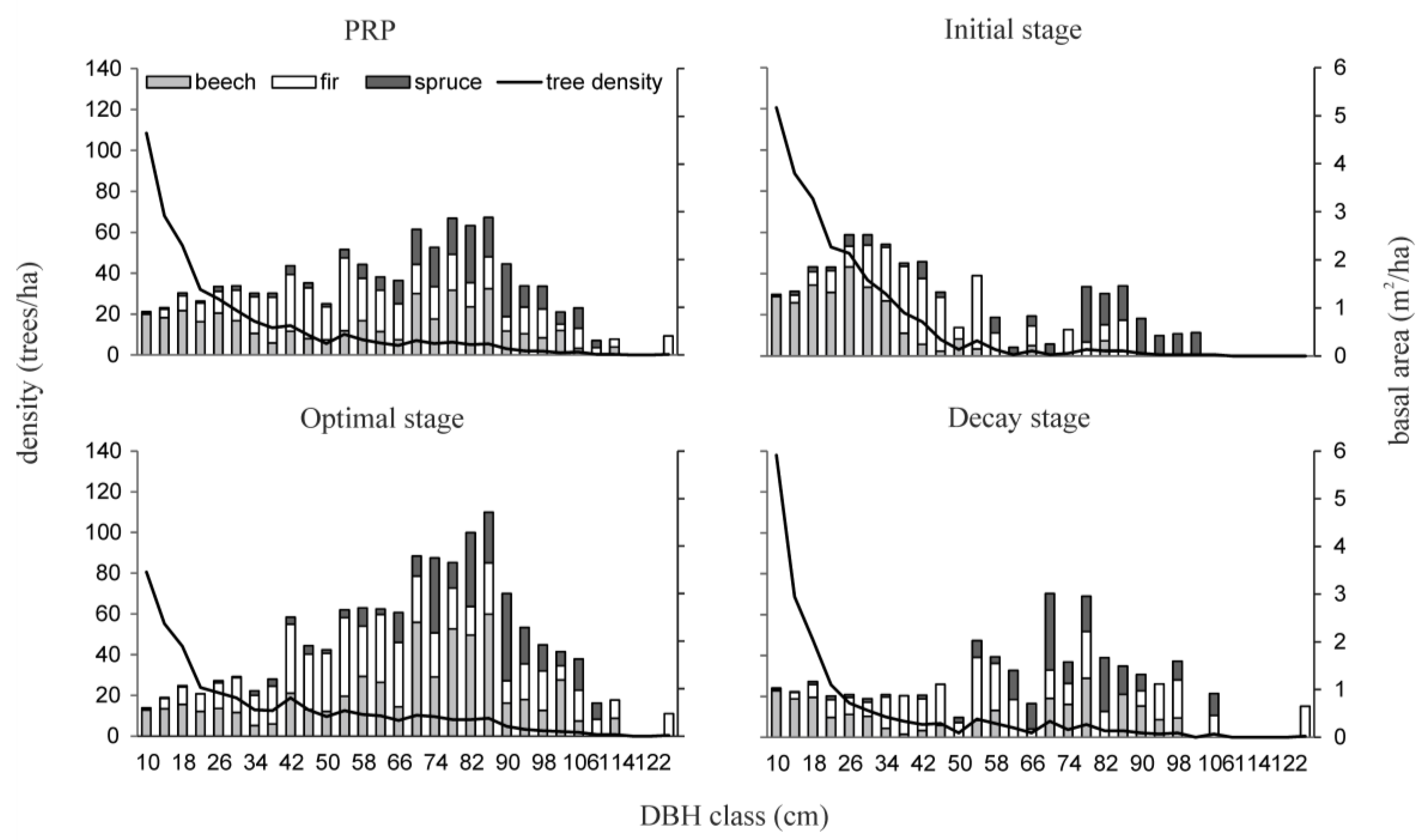
3.2. Attributes of the Old-Growth Forest Structure
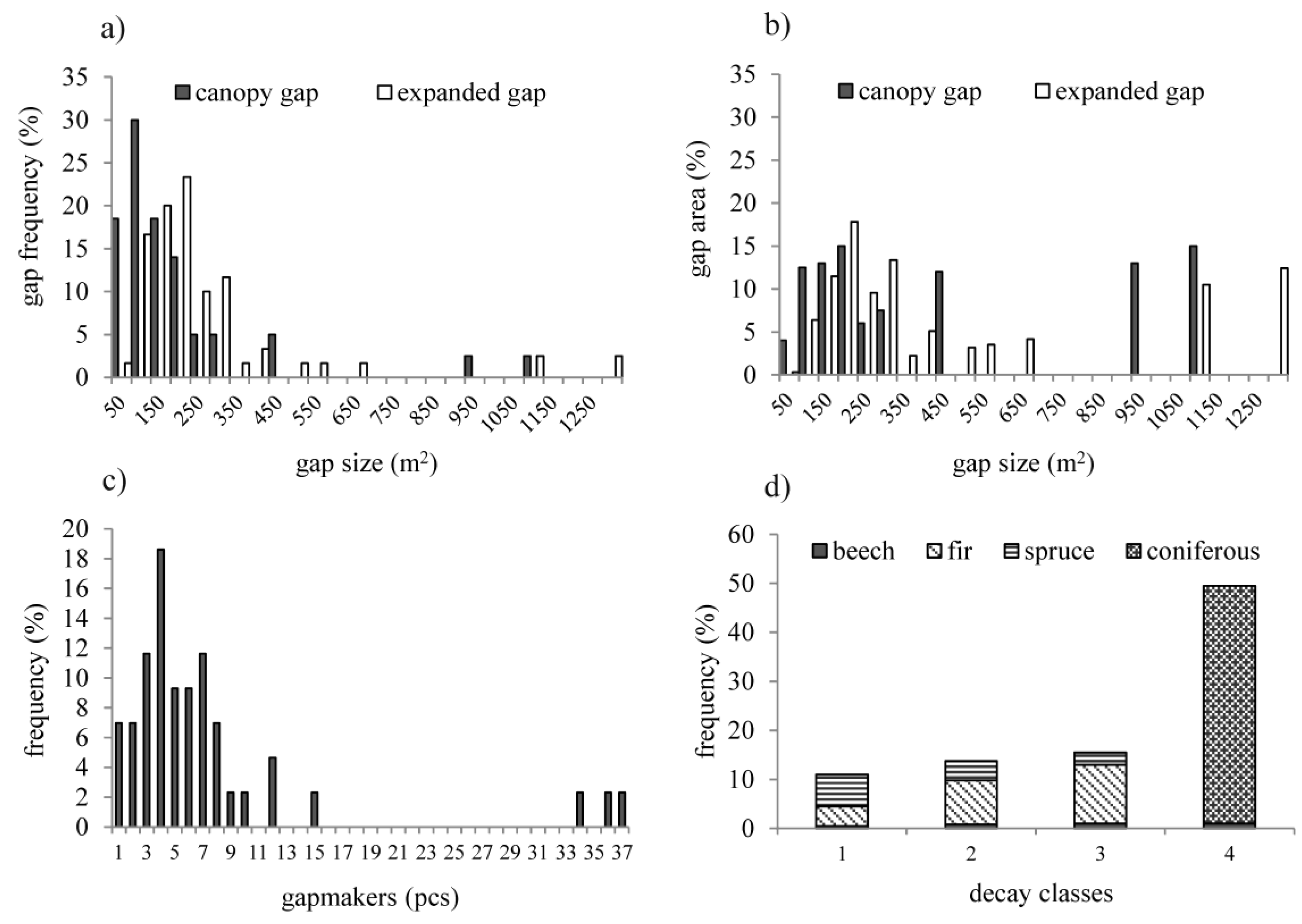
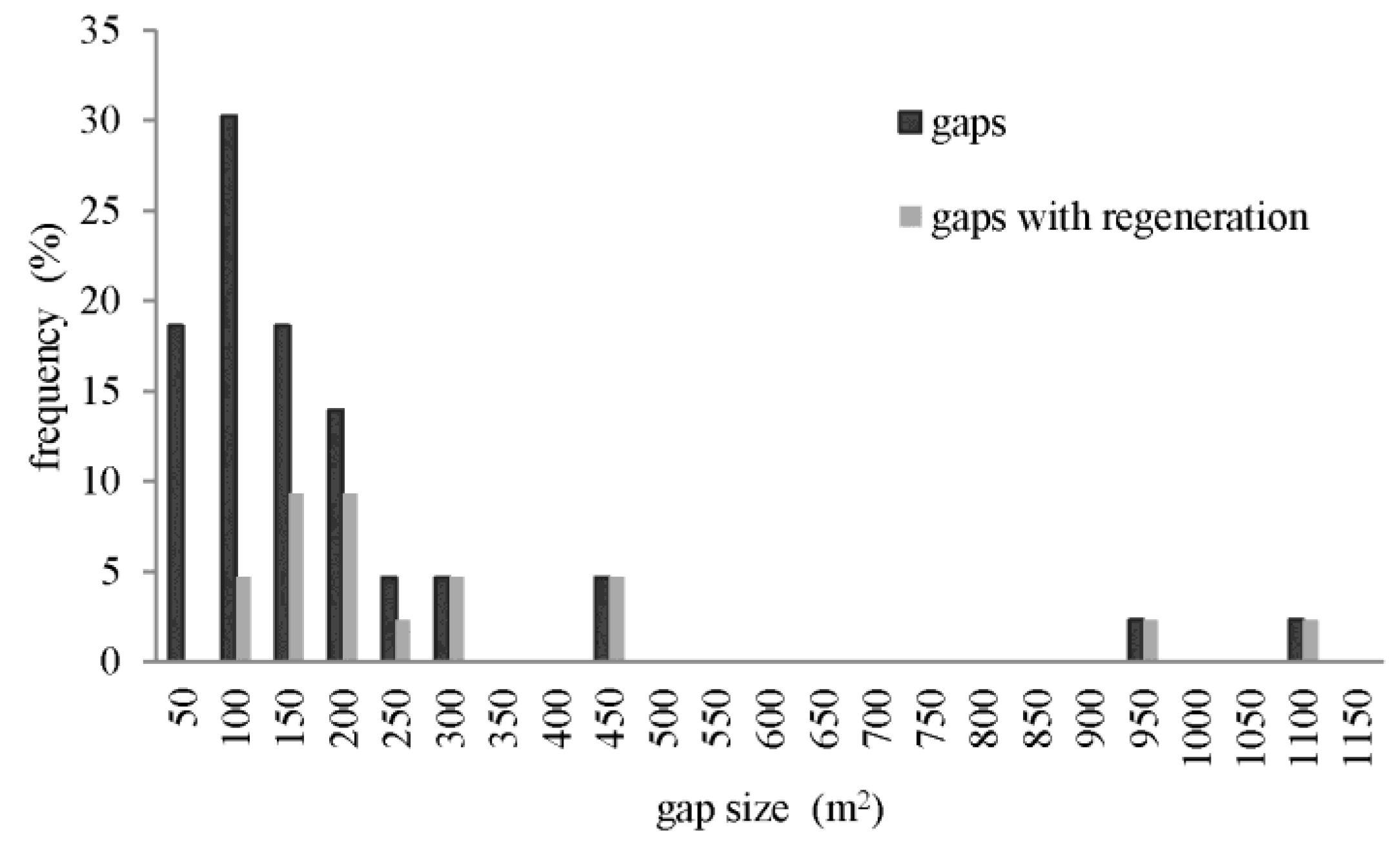
3.3. Structure of Canopy Gaps
4. Discussion
5. Summary
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Name | Main Tree Species | Size (ha) | Elevation (m) | Temperature (°C) | Precipitation (mm) | Forest Plant Community | Source of Data |
|---|---|---|---|---|---|---|---|
| Slovakia | |||||||
| Dobročský prales | F, B, S | 101.8 | 840–940 | 5.2 | 890 | Dentario enneaphylli-Fagetum | [42] |
| Ľubietovský Vepor | B, F, S | 236.9 | 950–1277 | 4 | 950 | Dentario enneaphylli-Fagetum | [84] |
| Klenovský Vepor | B, S, F | 122.1 | 1100–1338 | 4 | 950 | Dentario enneaphylli-Fagetum | [85] |
| Poland | |||||||
| Oszast | S, B, F | 43.3 | 950–1147 | 3 | 1300 | Dentario glandulosae-Fagetum | [86] |
| Zarnowka | B, F, S | - | 940–1010 | 4 | 1350 | Dentario glandulosae-Fagetum | [39] |
| Baniska | B, F, S | 142.0 | 775–1035 | 5 | 1200 | Dentario glandulosae-Fagetum | [39] |
| Czech Republic | |||||||
| Polom | S, B, F | 20.2 | 546–625 | 6 | 786 | Dentario enneaphylli-Fagetum Dentario glandulosae-Fagetum | [87] |
| Razula | F, B, S | 22.8 | 600–812 | 6.2 | 1088 | Dentario enneaphylli-Fagetum Dentario glandulosae-Fagetum | [14] |
| Salajka | B, F, S | 22.9 | 715–820 | 5.4 | 1144 | Dentario enneaphylli-Fagetum Dentario glandulosae-Fagetum | [14] |
| Criterion | Initial Stage | Optimal Stage | Decay Stage |
|---|---|---|---|
| General age estimation | Young | Adult | Old |
| Tree density | High | Medium | Low |
| Greatest proportion of trees by size | In small and medium sizes | In medium and large sizes | In large sizes |
| Canopy dominated by | Lower and middle layer | Upper and middle layer | Upper layer |
| Deadwood volume | Medium | Low | High |
| Decay stage | Higher decay classes | Various decay classes | Lower decay classes |
| Gaps | Absent | Absent | Present |
| Regeneration | >8 cm, clustered in former gaps | Little, scattered over the whole area | <8 cm, clustered in gaps |

References
- Mayer, H.; Ott, E. Gebirgswaldbau—Schutzwaldpflege; Gustav Fischer Verlag: Stuttgart, Germany, 1991. [Google Scholar]
- Korpeľ, Š. Die Urwälder der Westkarpaten; Gustav Fischer Verlag: New York, NY, USA, 1995. [Google Scholar]
- Meyer, P.; Tabaku, V.; Lüpke, B. Die Struktur albanischer Rotbuchen Urwäldern—Ableitung für eine naturnahe Buchenwirtschaft. Forstwiss. Cent. 2003, 122, 47–58. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Akhavan, R.; Sagheb-Talebi, K.; Zenner, E.K. Spatial patterns in different forest development stages of an intact old-growth Oriental beech forest in the Caspian region of Iran. Eur. J. For. Res. 2012, 131, 1355–1366. [Google Scholar] [CrossRef]
- Diaci, J.; Rozenbergar, D.; Mikac, S.; Anić, I.; Hartman, T.; Boncina, A. Long-term changes in tree species composition in old-growth Dinaric beech-fir forest. Glas. Šumske Pokuse 2008, 42, 13–27. [Google Scholar]
- Diaci, J.; Rozenbergar, D.; Anic, I.; Mikac, S.; Saniga, M.; Kucbel, S.; Visnjic, C.; Ballian, D. Structural dynamics and synchronous silver fir decline in mixed oldgrowth mountain forests in Eastern and Southeastern Europe. Forestry 2011, 84, 479–491. [Google Scholar] [CrossRef]
- Vrška, T.; Adam, D.; Hort, L.; Kolař, T.; Janik, D. European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) rotation in the Carpathians—A developmental cycle or a linear trend induced by man? For. Ecol. Manag. 2009, 258, 347–356. [Google Scholar] [CrossRef]
- Ficko, A.; Boncina, A. Silver fir (Abies alba Mill.) distribution in Slovenian forests. Res. Rep. For. Wood Sci. Technol. 2006, 79, 19–35. [Google Scholar]
- Klopcic, M.; Jerina, K.; Boncina, A. Long-Term Changes of Structure and Tree Species Composition in Dinaric Uneven-Aged Forests: Are Red Deer an Important Factor? Eur. J. For. Res. 2010, 129, 277–288. [Google Scholar] [CrossRef]
- Marini, L.; Okland, B.; Jonsson, A.M.; Bentz, B.; Carroll, A.; Forster, B.; Gregoire, J.C.; Hurling, R.; Nageleisen, L.M.; Netherer, S.; et al. Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 2017, 40, 1426–1435. [Google Scholar] [CrossRef]
- Keren, S.; Motta, R.; Govedar, Z.; Lucic, R.; Medarevic, M.; Diaci, J. Comparative structural dynamics of the Janj mixed old-growth mountain forest in Bosnia and Herzegovina: Are conifers in a long-term decline? Forests 2014, 5, 1243–1266. [Google Scholar] [CrossRef]
- Jaloviar, P.; Saniga, M.; Kucbel, S.; Pittner, J.; Vencurik, J.; Dovciak, M. Seven decades of change in a European old-growth forest following a stand-replacing wind disturbance: A long-term case study. For. Ecol. Manag. 2017, 399, 197–205. [Google Scholar] [CrossRef]
- Janík, D.; Adam, D.; Hort, L.; Král, K.; Šamonil, P.; Unar, P.; Vrška, T. Tree spatial patterns of Abies alba and Fagus sylvatica in the Western Carpathians over 30 years. Eur. J. For. Res. 2014, 133, 1015–1028. [Google Scholar] [CrossRef]
- Bouriaud, O.; Popa, I. Comparative dendroclimatic study of Scots pine, Norway spruce, and silver fir in the Vrancea Range, Eastern Carpathian Mountains. Trees 2009, 23, 95–106. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct effects of climate warming on populations of silver fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef]
- Nagel, T.A.; Levanic, T.; Diaci, J. A dendroecological reconstruction of disturbance in an old-growth Fagus-Abies forest in Slovenia. Ann. For. Sci. 2007, 64, 891–897. [Google Scholar] [CrossRef]
- Kucbel, S.; Jaloviar, P.; Saniga, M.; Vencurik, J.; Klimaš, V. Canopy gaps in an old-growth fir–beech forest remnant of Western Carpathians. Eur. J. For. Res. 2010, 129, 249–259. [Google Scholar] [CrossRef]
- Sefidi, K.; Marvie Mohadjer, M.R.; Mosandl, R.; Copenheaver, C.A. Canopy gaps and regeneration in old-growth Oriental beech (Fagus orientalis Lipsky) stands, northern Iran. For. Ecol. Manag. 2011, 262, 1094–1099. [Google Scholar] [CrossRef]
- Trotsiuk, V.; Hobi, M.L.; Commarmot, B. Age structure and disturbance dynamics of the relic virgin beech forest Uholka (Ukrainian Carpathians). For. Ecol. Manag. 2012, 265, 181–190. [Google Scholar] [CrossRef]
- Fischer, A.; Marshall, P.; Camp, A. Disturbances in deciduous temperate forest ecosystems of the Northern Hemisphere: Their effects on both recent and future development. Biodivers. Conserv. 2013, 22, 863–898. [Google Scholar] [CrossRef]
- Balanda, M.; Saniga, M.; Jaloviar, P.; Pittner, J. Structure, Production and Regeneration Processes of Natural Forests in NPR Hrončecký Grúň; Technical University in Zvolen: Zvolen, Slovakia, 2015; ISBN 978-80-228-2729-4. [Google Scholar]
- Elling, W.; Dittmar, C.; Pfaffelmoser, K.; Rötzer, T. Dendroecological Assessment of the Complex Causes of Decline and Recovery of the Growth of Silver Fir (Abies alba Mill.) in Southern Germany. For. Ecol. Manag. 2009, 257, 1175–1187. [Google Scholar] [CrossRef]
- Bošeľa, M.; Petráš, R.; Sitková, Z.; Priwitzer, T.; Pajtík, J.; Hlavatá, H.; Sedmák, R.; Tobin, B. Possible causes of the recent rapid increase in the radial increment of silver fir in the Western Carpathians. Environ. Pollut. 2014, 184, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.A.; Svoboda, M.; Rugani, T.; Diaci, J. Gap Regeneration and replacement patterns in an old-growth Fagus-Abies forest of Bosnia–Herzegovina. Plant Ecol. 2010, 208, 307–318. [Google Scholar] [CrossRef]
- Watt, A.S. Pattern and process in the plant community. J. Ecol. 1947, 35, 1–22. [Google Scholar] [CrossRef]
- Leibundgut, H. Europäische Urwälder: Wegweiser zur Naturnahen Waldwirtschaft; Verlag Paul Haupt: Bern und Stuttgart, Germany, 1993. [Google Scholar]
- Meyer, P. Determination of development phases and diversity of forest texture. AFJZ 1999, 170, 203–211. [Google Scholar]
- Petritan, I.C.; Commarmot, B.; Hobi, M.L.; Petritan, A.M.; Bigler, C.H.; Abrudan, I.V.; Rigling, A. Structural patterns of beech and silver fir suggest stability and resilience of the virgin forest Sinca in the Southern Carpathians, Romania. For. Ecol. Manag. 2015, 356, 184–195. [Google Scholar] [CrossRef]
- Zenner, E.K.; Hibbs, D.E. A new method for modeling the heterogeneity of forest structure. For. Ecol. Manag. 2000, 129, 75–87. [Google Scholar] [CrossRef]
- Pommerening, A. Approaches to quantifying forest structures. Forestry 2002, 75, 305–324. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. Zum Einfluss des Baumverteilungsmusters auf den Bestandeszuwachs. AFJZ 1995, 166, 190–201. [Google Scholar]
- Emborg, J. Understorey light condition and regeneration with respect to the structural dynamics of near-natural temperate deciduous forest in Denmark. For. Ecol. Manag. 1998, 106, 83–95. [Google Scholar] [CrossRef]
- Graz, F.P. The behaviour of the species mingling index Msp in relation to species dominance and dispersion. Eur. J. For. Res. 2004, 123, 87–92. [Google Scholar] [CrossRef]
- Drössler, L.; Meyer, P. Waldentwicklungsphasen in zwei Buchen-Urwaldreservaten in der Slowakei. Forstarchiv 2006, 77, 155–161. [Google Scholar]
- Král, K.; Vrška, T.; Hort, L.; Adam, D.; Šamonil, P. Development phases in a temperate natural spruce-fir-beech forest: Determination by a supervised classification method. Eur. J. For. Res. 2010, 129, 339–351. [Google Scholar] [CrossRef]
- Zenner, E.K.; Sagheb-Talebi, K.; Akhavan, R.; Peck, J.E. Integration of small-scale canopy dynamics smoothes live-tree structural complexity across development stages in old-growth Oriental beech (Fagus orientalis Lipsky) forests at the multi-gap scale. For. Ecol. Manag. 2015, 335, 26–36. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M. Gap disturbance regime in an old-growth Fagus-Abies forest in the Dinaric Mountains, Bosnia–Herzegovina. Can. J. For. Res. 2008, 38, 2728–2737. [Google Scholar] [CrossRef]
- Paluch, J.G.; Kołodziej, Z.; Pach, M.; Jastrzębski, R. Spatial variability of close-to-primeval Fagus-Abies-Picea forests in the Western Carpathians (Central Europe): A step towards a generalized pattern. Eur. J. For. Res. 2015, 134, 235–246. [Google Scholar] [CrossRef]
- Peck, J.E.; Commarmot, B.; Hobi, M.L.; Zenner, E.K. Should reference conditions be drawn from a single 10 ha plot? Assessing representativeness in a 10,000 ha old-growth European beech forest. Restor. Ecol. 2015, 23, 927–935. [Google Scholar] [CrossRef]
- Gömöryová, E.; Ujházy, K.; Martinák, M.; Gömöry, D. Soil microbial community response to variation in vegetation and abiotic environment in a temperate old-growth forest. Appl. Soil Ecol. 2013, 68, 10–19. [Google Scholar] [CrossRef]
- Ujházy, K.; Krížová, E.; Glončák, P.; Benčaťová, B.; Nič, J. Tree Species and Management Effect on Herb Layer Species Composition in Mountain Fir-Beech Forests of the Western Carpathians. In The Carpathians: Integrating Nature and Society Towards Sustainability; Kozak, J., Ostapowicz, K., Bytnerowicz, A., Wyzga, B., Eds.; Springer: Berlin, Germany, 2013; pp. 239–255. [Google Scholar]
- Tabaku, V. Structur von Buchen-Urwäldern in Albanien im Vergleich mit Deutchen Buchen-Naturwaldreservaten und -Wirtschaftswäldern. Ph.D. Thesis, Der Georg-August-Universität, Göttingen, Germany, November 1999. [Google Scholar]
- Albrecht, L. Grundlagen, Ziele und Methodik der Waldökologischen Forschung in Naturwaldreservaten; Schriftenreihe des Bayerischen Staatsministeriums für Ernährung, Landwirtschaft und Forsten gemeinsam mit dem Lehrstuhl für Landschaftstechnik: München, Germany, 1990; p. 221. [Google Scholar]
- Petráš, R.; Pajtík, J. Sústava česko–slovenských objemových tabuliek drevín. Lesn. Čas. 1991, 37, 49–56. [Google Scholar]
- Avery, T.E.; Burkhart, H.E. Forest Measurements, 5th ed.; McGraw-Hill: New York, NY, USA, 2002; ISBN 978-0-07-366176-6. [Google Scholar]
- Runkle, J.R. Patterns of disturbance in some old-growth mesic forests of Eastern North America. Ecology 1982, 63, 1533–1546. [Google Scholar] [CrossRef]
- Latham, P.A.; Zuuring, H.R.; Coble, D.W. A method for quantifying vertical forest structure. For. Ecol. Manag. 1998, 104, 157–170. [Google Scholar] [CrossRef]
- Shanon, C.E. The mathematical theory of communication. In The Mathematical Theory of Communication; Shannon, C.E., Weaver, W., Eds.; University of Illinois Press: Urbana, IL, USA, 1948; pp. 3–91. [Google Scholar]
- Camino, R. Zur Bestimmung der Bestandeshomogenität. Allg. For. Jagdztg. 1976, 147, 54–58. [Google Scholar]
- Reineke, L.H. Perfecting a stand density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Clark, P.J.; Evans, F.C. Distance to nearest neighbour as a measure of spatial relationships in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Donnelly, K. Simulations to determine the variance and edge effect of total nearest neighbour distance. In Simulation Methods in Archaeology; Hodder, J.R., Ed.; Cambridge University Press: London, UK, 1978; pp. 91–95. [Google Scholar]
- Füldner, K. Zur Strukturbeschreibung in Mischbeständen. Forstarchiv 1995, 66, 235–240. [Google Scholar]
- Hill, M.O.; Gauch, H.G. Detrended corespondence analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M.; Diaci, J. Regeneration patterns after intermediate wind disturbance in an old-growth Fagus–Abies forest in southeastern Slovenia. For. Ecol. Manag. 2006, 226, 268–278. [Google Scholar] [CrossRef]
- Klopcic, M.; Boncina, A. Recruitment of tree species in mixed selection and irregular shelterwood forest stands. Ann. For. Sci. 2012, 69, 915–925. [Google Scholar] [CrossRef] [Green Version]
- Keren, S.; Diaci, J.; Motta, R.; Govedar, Z. Stand structural complexity of mixed old-growth and adjacent selection forests in the Dinaric Mountains of Bosnia and Herzegovina. For. Ecol. Manag. 2017, 400, 531–541. [Google Scholar] [CrossRef]
- Parobeková, Z.; Saniga, M.; Pittner, J.; Kucbel, S.; Jaloviar, P. The Structure, Growing Stock Distribution, Disturbance Regime and Regeneration Processes of Virgin Forest Dobroč (Time Study); Technical University in Zvolen: Zvolen, Slovakia, 2016; ISBN 978-80-228-2853-6. [Google Scholar]
- Allen, M.S.; Thapa, V.; Arévalo, J.R.; Palmer, M.W. Windstorm damage and forest recovery: Accelerated succession, stand structure, and spatial pattern over 25 years in two Minnesota forests. Plant Ecol. 2012, 213, 1833–1842. [Google Scholar] [CrossRef]
- Büntgen, U.; Tegel, W.; Kaplan, J.O.; Schaub, M.; Hagedorn, F.; Bürgi, M.; Brázdil, R.; Helle, G.; Career, M.; Heussner, K.U.; et al. Placing unprecedented recent fir growth in a European-wide and Holocene-long context. Front. Ecol. Environ. 2014, 12, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Boden, S.; Kahle, H.P.; Wilpert, K.; Spiecker, H. Resilience of Norway spruce (Picea abies (L.) Karst) growth to changing climatic conditions in Southwest Germany. For. Ecol. Manag. 2014, 315, 12–21. [Google Scholar] [CrossRef]
- Bošeľa, M.; Sedmák, R.; Sedmáková, D.; Marušák, R.; Kulla, L. Temporal shifts of climate–growth relationships of Norway spruce as an indicator of health decline in the Beskids, Slovakia. For. Ecol. Manag. 2014, 325, 108–117. [Google Scholar] [CrossRef]
- Schütz, J.P.; Götz, M.; Schmid, W.; Mandallaz, D. Vulnerability of spruce (Picea abies) and beech (Fagus sylvatica) forest stands to storms and consequences for silviculture. Eur. J. For. Res. 2006, 125, 291–302. [Google Scholar] [CrossRef]
- Podlaski, R. A development cycle of the forest with fir (Abies alba Mill.) and beech (Fagus sylvatica L.) and its species composition in the Swietokrzyski National Park. J. For. Sci. 2004, 50, 55–66. [Google Scholar] [CrossRef]
- Heiri, C.; Wolf, A.; Rohreb, L.; Bugmann, H. Forty years of natural dynamics in Swiss beech forests: Structure, composition, and the influence of former management. Ecol. Appl. 2009, 19, 1920–1934. [Google Scholar] [CrossRef] [PubMed]
- Šamonil, P.; Antolik, L.; Svoboda, M.; Adam, D. Dynamics of windthrow events in a natural fir–beech forest in the Carpathian Mountains. For. Ecol. Manag. 2009, 257, 1148–1156. [Google Scholar] [CrossRef]
- Christensen, M.; Hahn, K.; Mountford, E.P.; Odor, P.; Standovar, T.; Rozenbergar, D.; Diaci, J.; Wijdeven, S.; Meyer, P.; Winter, S.; et al. Dead wood in European beech (Fagus sylvatica) forest reserves. For. Ecol. Manag. 2005, 210, 267–282. [Google Scholar] [CrossRef]
- Korpeľ, Š. Pralesy Slovenska; Veda: Bratislava, Slovensko, 1989. [Google Scholar]
- Saniga, M. Structure, Production and Regeneration Processes of the Dobroč Virgin Forest; Technical University in Zvolen: Zvolen, Slovakia, 1999. [Google Scholar]
- Motta, R.; Garbarino, M.; Berretti, R.; Meloni, F.; Vacchiano, G. Development of old-growth characteristics in uneven-aged forests of the Italian Alps. Eur. J. For. Res. 2015, 134, 19–31. [Google Scholar] [CrossRef]
- Lombardi, F.; Cherubini, P.; Tognetti, R.; Cocozza, C.; Lasserre, B.; Marchetti, M. Investigating biochemical processes to assess deadwood decay of beech and silver fir in Mediterranean mountain forests. Ann. For. Sci. 2013, 70, 101–111. [Google Scholar] [CrossRef]
- Balanda, M. Spatio-temporal structure of natural forest: A structural index approach. Beskydy 2012, 5, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Zeibig, A.; Diaci, J.; Wagner, S. Gap disturbance patterns of a Fagus sylvatica virgin forest remnant in the mountain vegetation belt of Slovenia. For. Snow Landsc. Res. 2005, 79, 69–80. [Google Scholar]
- Drössler, L.; Luepke, B. Canopy gaps in two virgin beech forest reserves in Slovakia. J. For. Sci. 2005, 51, 446–457. [Google Scholar] [CrossRef]
- Kenderes, K.; Král, K.; Vrška, T.; Standovár, T. Natural gap dynamics in a Central European mixed beech–spruce–fir oldgrowth forest. Ecoscience 2009, 16, 39–47. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics, 2nd ed.; John Wiley & Sons Inc.: New York, NY, USA, 1996; 520p. [Google Scholar]
- Vencurik, J.; Kucbel, S.; Saniga, M.; Jaloviar, P.; Parobeková, Z.; Sedmáková, D.; Pittner, J. Stand structure, light microclimate and distribution of lower layer individuals in selection forest in two localities of Slovakia. Zprávy Lesn. Výzkumu 2016, 61, 230–237. [Google Scholar]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schütz, J.-P. Silvicultural tools to develop irregular and diverse forest structures. Forestry 2002, 75, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Mohr, C.; Schori, C. Femelschlag oder Plenterung—Ein Vergleich aus betriebswirtschaftlicher Sicht. Schweiz. Z. Forstwes. 1999, 150, 49–55. [Google Scholar] [CrossRef]
- Bončina, A.; Gasperšič, F.; Diaci, J. Long-term changes in tree species composition in the Dinaric mountain forests of Slovenia. For. Chron. 2003, 79, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Saniga, M.; Balanda, M.; Kucbel, S.; Jaloviar, P. Cyclic changes in tree species composition of mixed-species forest in Western Carpathians: Role of disturbance and tree regeneration. Pol. J. Ecol. 2011, 59, 381–389. [Google Scholar]
- Ujházy, K.; Hederová, L.; Máliš, F.; Ujházyová, M.; Bošeľa, M.; Čiliak, M. Overstorey dynamics controls plant diversity in age-class temperate forests. For. Ecol. Manag. 2017, 391, 96–105. [Google Scholar] [CrossRef]
- Miadok, D. Beitrag zur Kenntnis der Buchenwälder im Gebirge Klenovský Vepor. Acta Fac. Rerum Nat. Univ. Comenianae 1971, 17, 99–115. [Google Scholar]
- Jaworski, A.; Pach, M. A comparison of lower montane natural forest (Abies, Fagus, Picea) in Oszast Reserve and spruce monocultures in the Zywiecki Beskid and Slaski Beskid. For. Res. Pap. 2014, 75, 13–32. [Google Scholar] [CrossRef] [Green Version]
- Vrška, T.; Hort, L.; Adam, D.; Odehnalová, P.; Horal, D. Developmental Dynamics of Virgin Forest Reserves in the Czech Republic I; Academia: Prague, Czech Republic, 2002; p. 213. [Google Scholar]





| Tree Species | 1978 | 2015 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stem Density | Basal Area | Growing Stock | Stem Density | Basal Area | Growing Stock | |||||||
| N·ha−1 | % | m2·ha−1 | % | m3·ha−1 | % | N·ha−1 | % | m2·ha−1 | % | m3·ha−1 | % | |
| Fir | 139 | 38 | 17.6 | 41 | 261 | 38 | 81 | 20 | 17.8 | 39 | 280 | 38 |
| Beech | 125 | 35 | 9.2 | 22 | 161 | 24 | 257 | 65 | 15.6 | 34 | 258 | 35 |
| Spruce | 90 | 25 | 14.7 | 34 | 238 | 35 | 49 | 12 | 10.5 | 23 | 174 | 24 |
| Others * | 8 | 2 | 1.2 | 3 | 21 | 3 | 9 | 2 | 1.4 | 3 | 25 | 3 |
| Total | 362 | 100 | 42.7 | 100 | 682 | 100 | 396 | 100 | 45.3 | 100 | 737 | 100 |
| Category | Beech | Fir | Spruce | Maples | Ash | Total |
|---|---|---|---|---|---|---|
| ≤20 cm | 2143 | 9190 | 164 | 3435 | 730 | 15,661 |
| 21–50 cm | 573 | 16 | 3 | 525 | 676 | 1793 |
| 51–130 cm | 114 | 0 | 0 | 77 | 79 | 270 |
| >130 cm * | 497 | 2 | 0 | 0 | 2 | 502 |
| Total | 3327 | 9208 | 167 | 4037 | 1487 | 18,226 |
| Stand Characteristics | Initial Stage | Optimal Stage | Decay Stage | PRP | |
|---|---|---|---|---|---|
| Spatial proportion | (%) | 26 | 44 | 30 | 100 |
| Tree density ≥ 8 cm | (N·ha−1) | 531 ± 61 b | 413 ± 131 a | 398 ± 125 a | 413 ± 113 |
| Basal area | (m2·ha−1) | 32.2 ± 6.8 a | 60.2 ± 10.1 b | 33.3 ± 11.2 a | 44.9 ± 6.2 |
| Growing stock | (m3·ha−1) | 459 ± 133 a | 1005 ± 196 b | 529 ± 203 a | 731 ± 149 |
| Fir | (%) | 38 ± 24 a | 35 ± 23 a | 41 ± 23 a | 40 ± 23 |
| Beech | (%) | 31 ± 14 a | 39 ± 26 a | 29 ± 24 a | 33 ± 23 |
| Spruce | (%) | 26 ± 22 a | 22 ± 25 a | 27 ± 29 a | 22 ± 25 |
| Others * | (%) | 9 ± 17 b | 4 ± 5 ab | 2 ± 4 a | 5 ± 10 |
| Snags | (m3·ha−1) | 26 ± 39 a | 49 ± 73 ab | 80 ± 59 b | 53 ± 64 |
| Logs | (m3·ha−1) | 264 ± 171 ab | 185 ± 136 a | 353 ± 232 b | 260 ± 191 |
| Crown engagement ratio | 1.5 ± 0.16 b | 1.7 ± 0.41 c | 1.0 ± 0.30 a | 1.4 ± 0.4 | |
| Growth space utilisation | 0.3 ± 0.09 a | 0.4 ± 0.10 b | 0.2 ± 0.09 a | 0.3 ± 0.1 |
| Index | Stand Layer | Initial | Optimal | Decay | PRP |
|---|---|---|---|---|---|
| H′N | lower | 0.08 ± 0.07 a | 0.22 ± 0.15 ab | 0.29 ± 0.26 ab | 0.27 |
| medium | 0.42 ± 0.15 b | 0.48 ± 0.27 b | 0.26 ± 0.38 ab | 0.69 | |
| upper | 0.88 ± 0.24 c | 0.77 ± 0.26 c | 0.76 ± 0.36 c | 1.19 | |
| total | 0.51 ± 0.10 | 0.53 ± 0.18 | 0.65 ± 0.12 | 0.62 | |
| H′BA | lower | 0.28 ± 0.23 a | 0.48 ± 0.27 abc | 0.38 ± 0.29 ab | 0.63 |
| medium | 0.58 ± 0.18 abc | 0.51 ± 0.27 abc | 0.33 ± 0.47 ab | 0.84 | |
| upper | 0.81 ± 0.31 c | 0.63 ± 0.36 abc | 0.66 ± 0.41 bc | 1.16 | |
| total | 0.99 ± 0.16 | 0.83 ± 0.24 | 0.83 ± 0.33 | 1.16 | |
| R | lower | 0.96 ± 0.13 | 0.93 ± 0.11 | 0.90 ± 0.16 | 0.90 *** |
| medium | 1.10 ± 0.15 | 1.05 ± 0.15 | 1.02 ± 0.02 | 1.03 | |
| upper | 1.05 ± 0.28 | 1.07 ± 0.20 | 1.15 ± 0.19 | 1.04 | |
| total | 1.03 ± 0.09 | 1.08 ± 0.06 | 1.05 ± 0.06 | 1.03 | |
| H | 1.41 ± 0.12 | 1.51 ± 0.26 | 1.40 ± 0.17 | 2.00 | |
| EN | 0.45 ± 0.08 a | 0.50 ± 0.13 a | 0.69 ± 0.20 b | 0.39 | |
| EBA | 0.87 ± 0.09 | 0.79 ± 0.15 | 0.81 ± 0.13 | 0.72 | |
| G | 0.38 ± 0.03 | 0.36 ± 0.06 | 0.41 ± 0.06 | 0.38 | |
| T | 0.55 ± 0.03 b | 0.50 ± 0.04 a | 0.56 ± 0.02 b | 0.51 | |
| SCId | 10.22 ± 1.31 | 9.95 ± 1.83 | 11.29 ± 1.96 | 9.90 | |
| SCIh | 6.37 ± 0.53 | 6.21 ± 0.68 | 6.61 ± 0.79 | 6.15 |
| Category | Beech | Maples | Ash | Total | ||
|---|---|---|---|---|---|---|
| m2·ha−1 | m2·ha−1 | m2·ha−1 | m2·ha−1 | % | ||
| ≤50 cm | 107 | 89 | 196 | 13 | ||
| 51–130 cm | 18 | 102 | 120 | 8 | ||
| >130 cm * | 508 | 10 | 15 | 533 | 36 | |
| dbh 2–8 cm | 646 | 646 | 43 | |||
| total | m2·ha−1 | 1154 | 135 | 206 | 1495 | 100 |
| % | 77 | 9 | 14 | 100 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parobeková, Z.; Pittner, J.; Kucbel, S.; Saniga, M.; Filípek, M.; Sedmáková, D.; Vencurik, J.; Jaloviar, P. Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians. Forests 2018, 9, 379. https://doi.org/10.3390/f9070379
Parobeková Z, Pittner J, Kucbel S, Saniga M, Filípek M, Sedmáková D, Vencurik J, Jaloviar P. Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians. Forests. 2018; 9(7):379. https://doi.org/10.3390/f9070379
Chicago/Turabian StyleParobeková, Zuzana, Ján Pittner, Stanislav Kucbel, Milan Saniga, Michal Filípek, Denisa Sedmáková, Jaroslav Vencurik, and Peter Jaloviar. 2018. "Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians" Forests 9, no. 7: 379. https://doi.org/10.3390/f9070379
APA StyleParobeková, Z., Pittner, J., Kucbel, S., Saniga, M., Filípek, M., Sedmáková, D., Vencurik, J., & Jaloviar, P. (2018). Structural Diversity in a Mixed Spruce-Fir-Beech Old-Growth Forest Remnant of the Western Carpathians. Forests, 9(7), 379. https://doi.org/10.3390/f9070379