Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment


Abstract
:1. Introduction
2. Materials and Methods
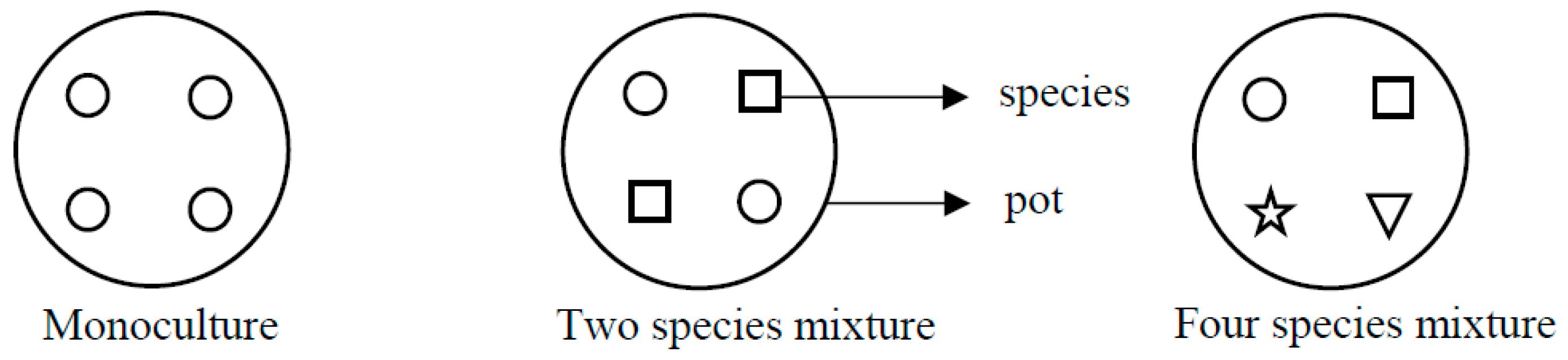
2.1. Experimental Design
2.2. Measurements
2.3. Data Analysis
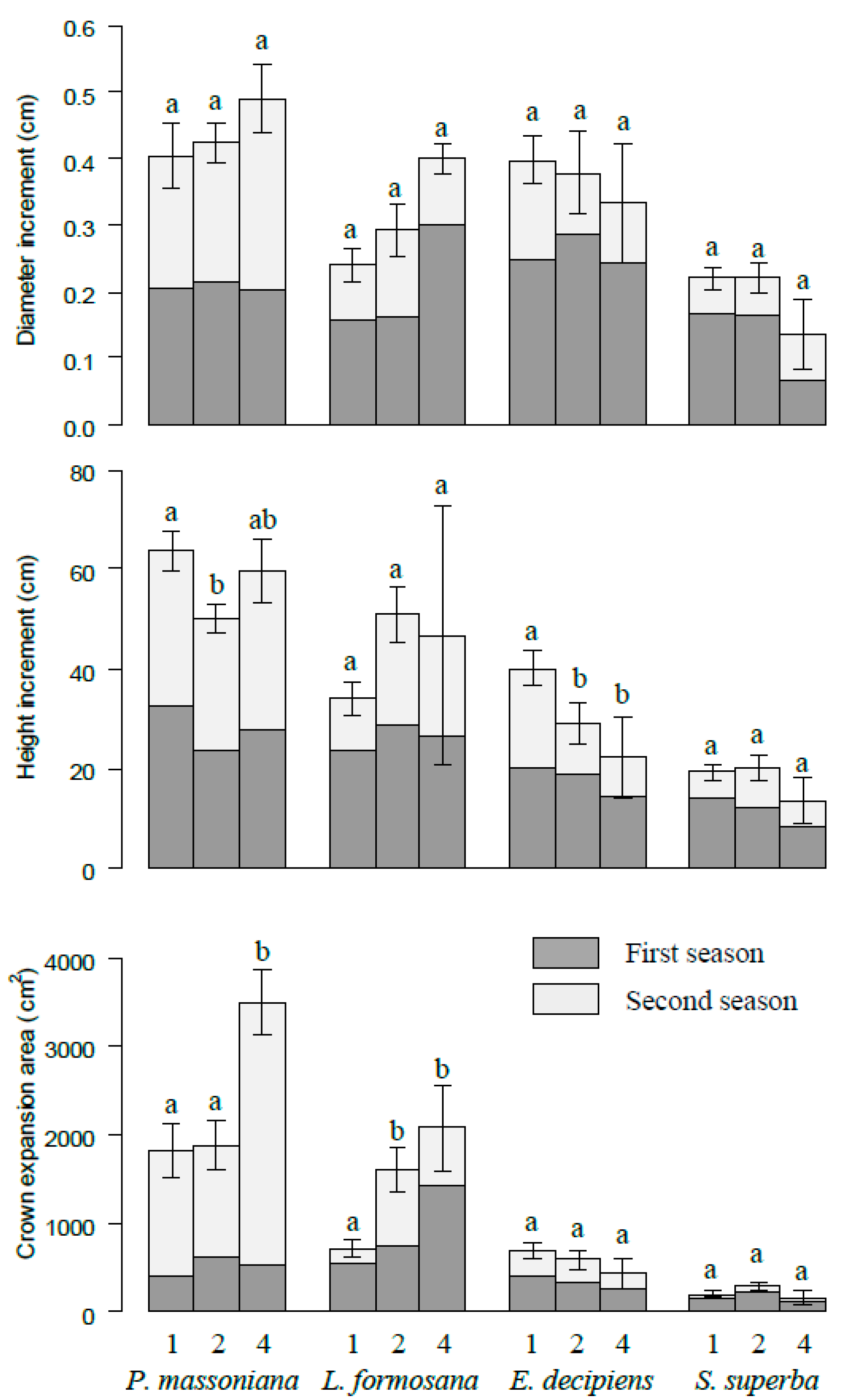
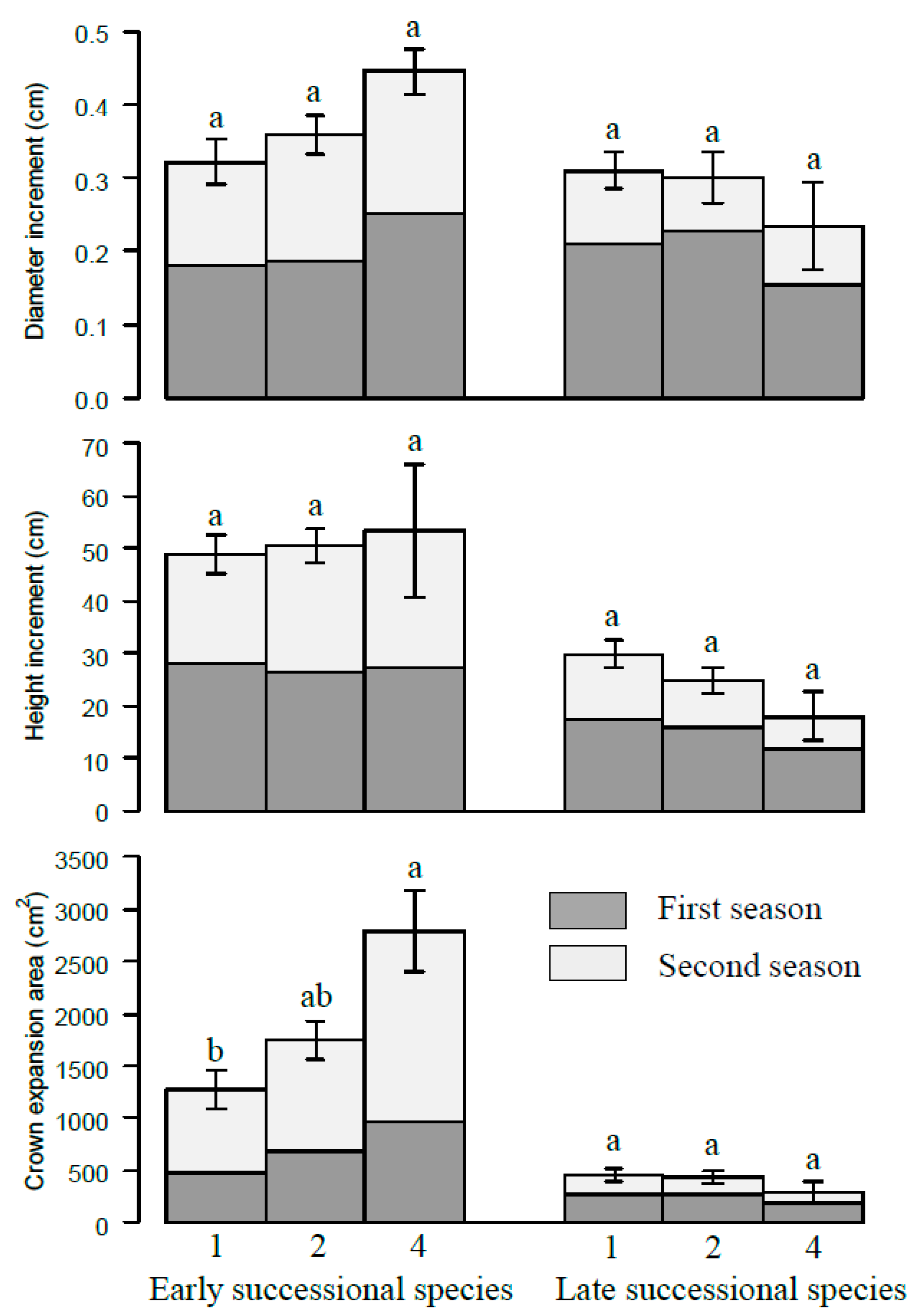
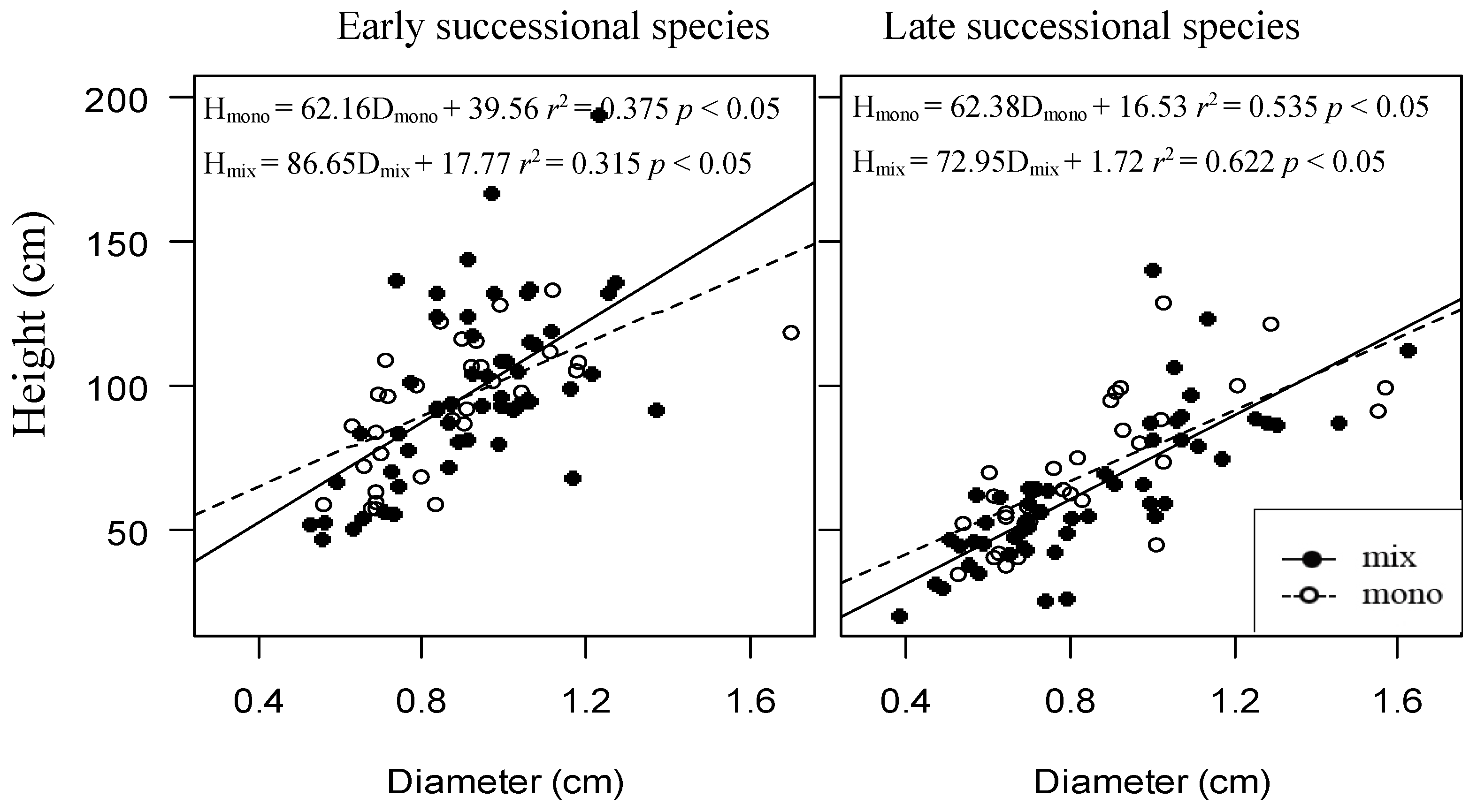
3. Results
4. Discussion
4.1. The Relationship between Tree Growth and Species Richness
4.2. Species-Specific Growth Variation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- Naeem, S.; Thompson, L.J.; Lawler, S.P.; Woodfin, R.M. Declining biodiversity can alter the performance of ecosystems. Nature 1994, 368, 734–737. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Hooper, D.U.; Vitousek, P.M. The effects of plant composition and diversity on ecosystem processes. Science 1997, 277, 1302–1305. [Google Scholar] [CrossRef]
- Hector, A.; Schmid, B.; Beierkuhnlein, C.; Caldeira, M.; Diemer, M.; Dimitrakopoulos, P.G.; Finn, J.A.; Freitas, H.; Giller, P.S.; Good, J.; et al. Plant diversity and productivity experiments in European grasslands. Science 1999, 286, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Kyoto Protocol to the United Nations Framework Convention on Climate Change. Available online: https://unfccc.int/resource/docs/convkp/kpeng.pdf (accessed on 20 April 2018).
- Li, Y.; Härdtle, W.; Bruelheide, H.; Nadrowski, K.; Scholten, T.; Wehrden, H.V.; Oheimb, G.V. Site and neighborhood effects on growth of tree saplings in subtropical plantations (China). For. Ecol. Manag. 2014, 327, 118–127. [Google Scholar] [CrossRef]
- Vilà, M.; Carrillo-Gavilán, A.; Vayreda, J.; Bugmann, H.; Fridman, J.; Grodzki, W.; Haase, J.; Kunstler, G.; Schelhaas, M.J.; Trasobares, A. Disentangling biodiversity and climatic determinants of wood production. PLoS ONE 2007, 8, e53530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilà, M.; Vayreda, J.; Comas, L.; Ibáñez, J.J.; Mata, T.; Obón, B. Species richness and wood production: A positive association in Mediterranean forests. Ecol. Lett. 2007, 10, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Kent, M.; Fang, X.F. Evergreen broad-leaved forest in Eastern China: Its ecology and conservation and the importance of resprouting in forest restoration. For. Ecol. Manag. 2007, 245, 76–87. [Google Scholar] [CrossRef]
- Lang, A.C.; Härdtle, W.; Baruffol, M.; Böhnke, M.; Bruelheide, H.; Schmid, B.; Von-Wehrden, H.; von Oheimb, G. Mechanisms promoting tree species co-existence: Experimental evidence with saplings of subtropical forest ecosystems of China. J. Veg. Sci. 2012, 23, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Von Oheimb, G.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.; Härdtle, W. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighborhood competition. For. Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Lang, A.C.; Härdtle, W.; Bruelheide, H.; Geißler, C.; Nadrowski, K.; Schuldt, A.; Yu, M.J.; Von-Oheimb, G. Tree morphology responds to neighborhood competition and slope in species-rich forests of subtropical China. For. Ecol. Manag. 2010, 260, 1708–1715. [Google Scholar] [CrossRef]
- Longuetaud, F.; Seifert, T.; Leban, J.M.; Pretzsch, H. Analysis of long-term dynamics of crowns of sessile oaks at the stand level by means of spatial statistics. For. Ecol. Manag. 2008, 255, 2007–2019. [Google Scholar] [CrossRef]
- Longuetaud, F.; Piboule, A.; Wernsdorfer, H.; Collet, C. Crown plasticity reduces inter-tree competition in a mixed broadleaved forest. Eur. J. For. Res. 2013, 132, 621–634. [Google Scholar] [CrossRef]
- He, Y.J.; Liang, X.Y.; Qin, L.; Li, Z.Y.; Shao, M.X.; Tan, L. Community characteristics and soil properties of coniferous plantation forest monocultures in the early stages after close-to-nature transformation management in southern subtropical China. Acta Ecol. Sin. 2013, 33, 2484–2495. [Google Scholar]
- Liu, C.; Xiang, W.H.; Lei, P.F.; Deng, X.W.; Tian, D.L.; Fang, X.; Peng, C.H. Standing fine root mass and production in four Chinese subtropical forests along a succession and species diversity gradient. Plant Soil 2014, 376, 445–459. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ratcliffe, S.; Holzwarth, F.; Nadrowski, K.; Levick, S.; Wirth, C. Tree neighbourhood matters—Tree species composition drives diversity–productivity patterns in a near-natural beech forest. For. Ecol. Manag. 2015, 335, 225–234. [Google Scholar] [CrossRef]
- Simard, S.W.; Zimonick, B.J. Neighborhood size effects on mortality, growth and crown morphology of paper birch. For. Ecol. Manag. 2005, 214, 251–265. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: Evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot size matters: A meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef]
- Potvin, C.; Gotelli, N.J. Biodiversity enhances individual performance but does not affect survivorship in tropical trees. Ecol. Lett. 2008, 11, 217–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattge, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Jacob, M.; Leuschner, C.; Thomas, F.M. Productivity of temperate broad-leaved forest stands differing in tree species diversity. Ann. For. Sci. 2010, 67, 503. [Google Scholar] [CrossRef]
- Nguyen, H.; Herbohn, J.; Firn, J.; Lamb, D. Biodiversity-productivity relationships in small-scale mixed-species plantations using native species in Leyte province, Philippines. For. Ecol. Manag. 2012, 274, 81–90. [Google Scholar] [CrossRef]
- Bruelheide, H.; Nadrowski, K.; Assmann, T.; Bauhus, J.; Both, S.; Buscot, F.; Chen, X.Y.; Ding, B.Y.; Durka, W.; Erfmeier, A.; et al. Designing forest biodiversity experiments: General considerations illustrated by a new large experiment in subtropical China. Methods Ecol. Evol. 2014, 5, 74–89. [Google Scholar] [CrossRef]
- Chen, Z.H.; Wang, B.S.; Zhang, H.D. Growth of the trees and saplings in the lower subtropical evergreen broad-leaved forest in Heishiding, Guangdon Province. Acta Phytoecol. Sin. 1999, 23, 441–450. [Google Scholar]
- Xing, P.; Zhang, Q.B.; Baker, P.J. Age and radial growth pattern of four tree species in a subtropical forest of China. Trees 2012, 26, 283–290. [Google Scholar] [CrossRef]
- Forrester, D.I.; Benneter, A.; Bouriaud, O.; Bauhus, J. Diversity and competition influence tree allometric relationships: Developing functions for mixed-species forests. J. Ecol. 2017, 105, 761–774. [Google Scholar] [CrossRef]
- Forrester, D.I.; Pretzsch, H. Tamm Review: On the strength of evidence when comparing ecosystem functions of mixtures with monocultures. For. Ecol. Manag. 2015, 356, 41–53. [Google Scholar] [CrossRef]
- Bauhus, J.; Khanna, P.K.; Menden, N. Aboveground and belowground interactions in mixed plantations of Eucalyptus globulus and Acacia mearnsii. Can. J. For. Res. 2000, 30, 1886–1894. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Richness | Diameter (cm) | Height (cm) | Crown Projection Area (cm2) | n |
|---|---|---|---|---|---|
| P. massoniana Lamb. | 1 | 0.88 ± 0.07 | 92.62 ± 5.28 | 2381.79 ± 359.88 | 15 |
| 2 | 0.87 ± 0.04 | 84.68 ± 3.98 | 2332.70 ± 281.20 | 24 | |
| 4 | 1.01 ± 0.09 | 100.35 ± 11.00 | 3906.15 ± 405.29 | 4 | |
| L. formosana Hance. | 1 | 0.83 ± 0.04 | 92.51 ± 6.76 | 859.08 ± 106.40 | 15 |
| 2 | 0.89 ± 0.04 | 105.84 ± 6.32 | 1879.22 ± 283.89 | 24 | |
| 4 | 1.21 ± 0.03 | 116.35 ± 30.22 | 2747.92 ± 451.11 | 4 | |
| S. superba Champ. | 1 | 0.67 ± 0.02 | 52.97 ± 2.72 | 602.94 ± 58.81 | 16 |
| 2 | 0.64 ± 0.02 | 50.54 ± 2.10 | 604.17 ± 57.49 | 22 | |
| 4 | 0.49 ± 0.06 | 27.10 ± 2.32 | 326.33 ± 26.35 | 4 | |
| E. decipiens Hemsl | 1 | 1.09 ± 0.07 | 90.37 ± 4.95 | 1101.05 ± 136.35 | 16 |
| 2 | 1.04 ± 0.05 | 75.58 ± 4.94 | 969.66 ± 183.31 | 22 | |
| 4 | 0.76 ± 0.09 | 62.88 ± 17.61 | 418.12 ± 254.01 | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Lei, P.; Xiang, W.; Ouyang, S.; Hui, X. Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment. Forests 2018, 9, 380. https://doi.org/10.3390/f9070380
Yang J, Lei P, Xiang W, Ouyang S, Hui X. Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment. Forests. 2018; 9(7):380. https://doi.org/10.3390/f9070380
Chicago/Turabian StyleYang, Jingjun, Pifeng Lei, Wenhua Xiang, Shuai Ouyang, and Xinrong Hui. 2018. "Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment" Forests 9, no. 7: 380. https://doi.org/10.3390/f9070380
APA StyleYang, J., Lei, P., Xiang, W., Ouyang, S., & Hui, X. (2018). Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment. Forests, 9(7), 380. https://doi.org/10.3390/f9070380