Simulating Climate Change Impacts on Hybrid-Poplar and Black Locust Short Rotation Coppices


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
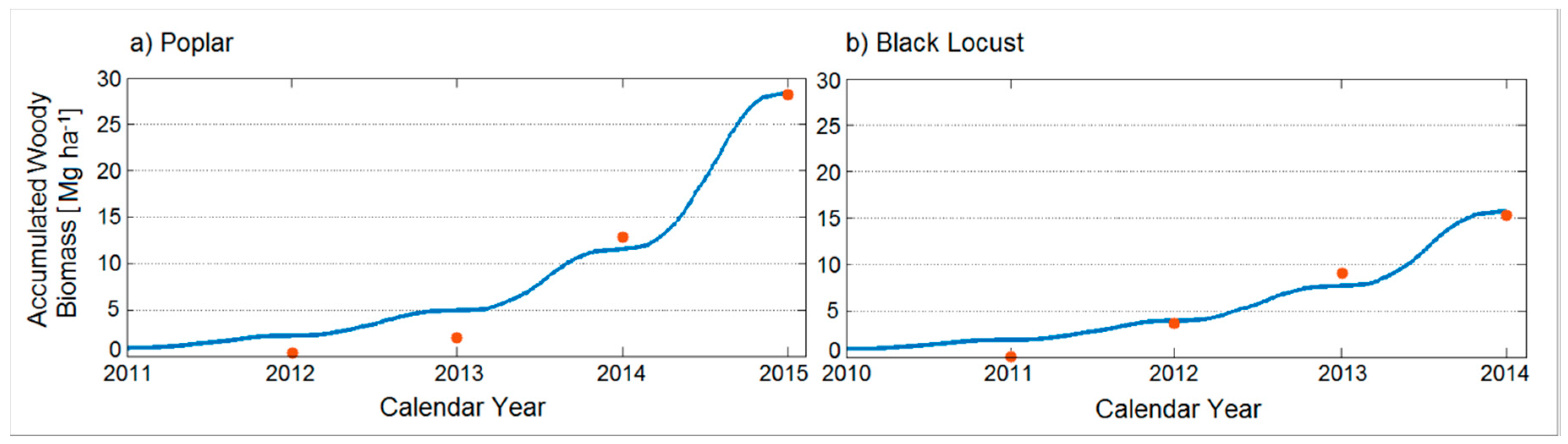
2.2. Yearly Measurements of Above-Ground Woody Biomass
2.3. Modelling the Above-Ground Woody Biomass
2.3.1. Description of the Yield-SAFE Model
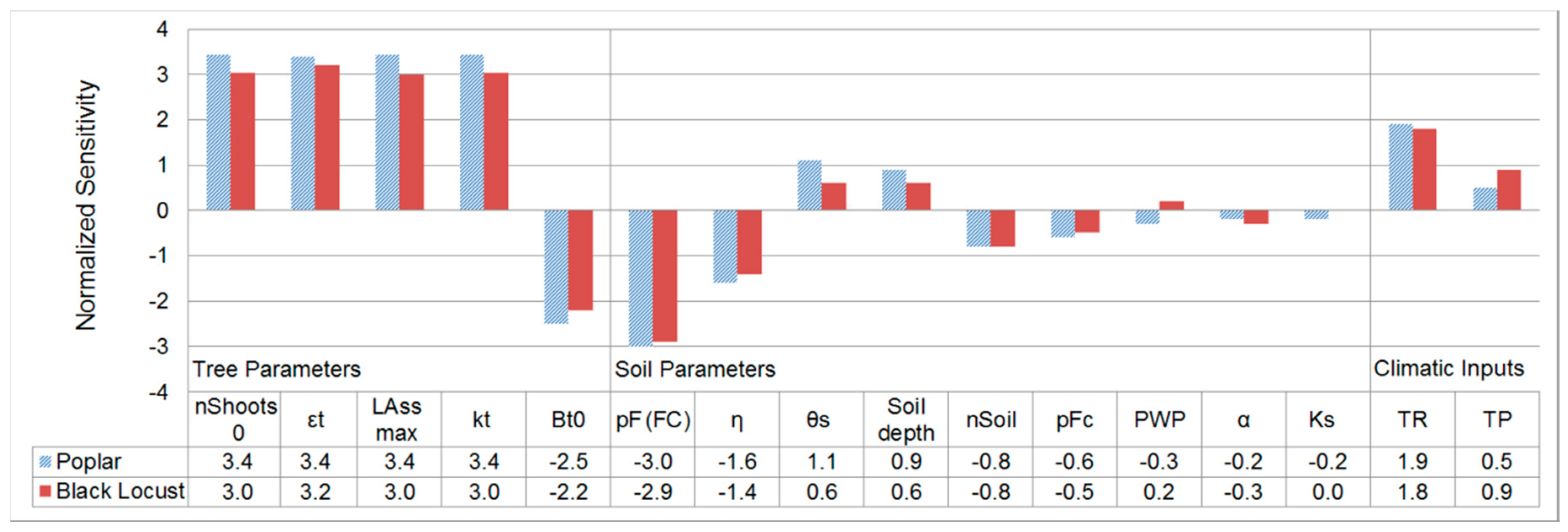
2.3.2. Sensitivity Analysis of the Yield-SAFE Model
2.3.3. Parameterization and Validation of the Yield-SAFE Model
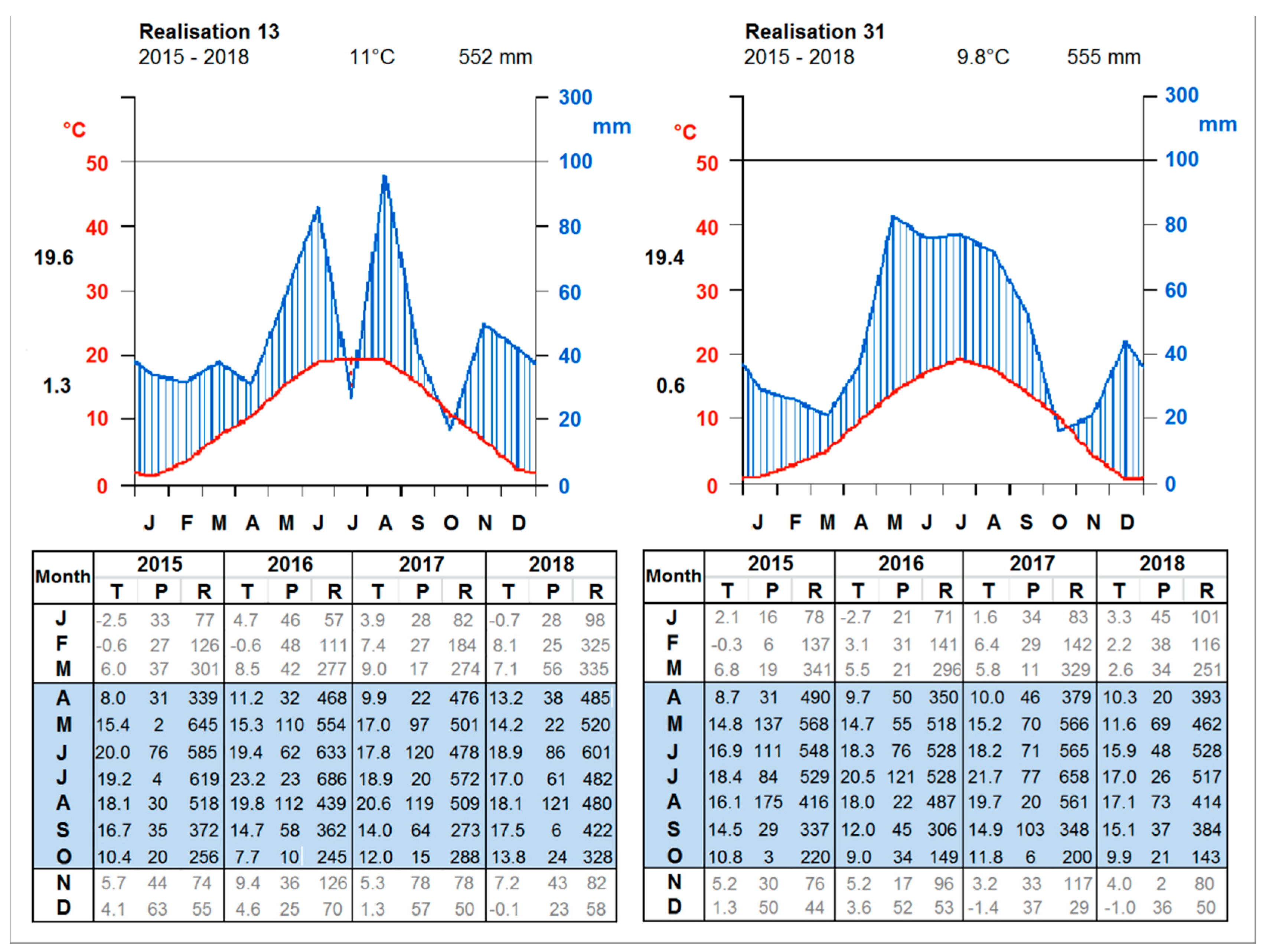
2.4. Prospective Climate Change
3. Results
3.1. Observed Woody Biomass Productivity of Poplar and Black Locust Trees
3.2. Sensitivity Analysis of the Yield-SAFE Model
3.3. Model Validation
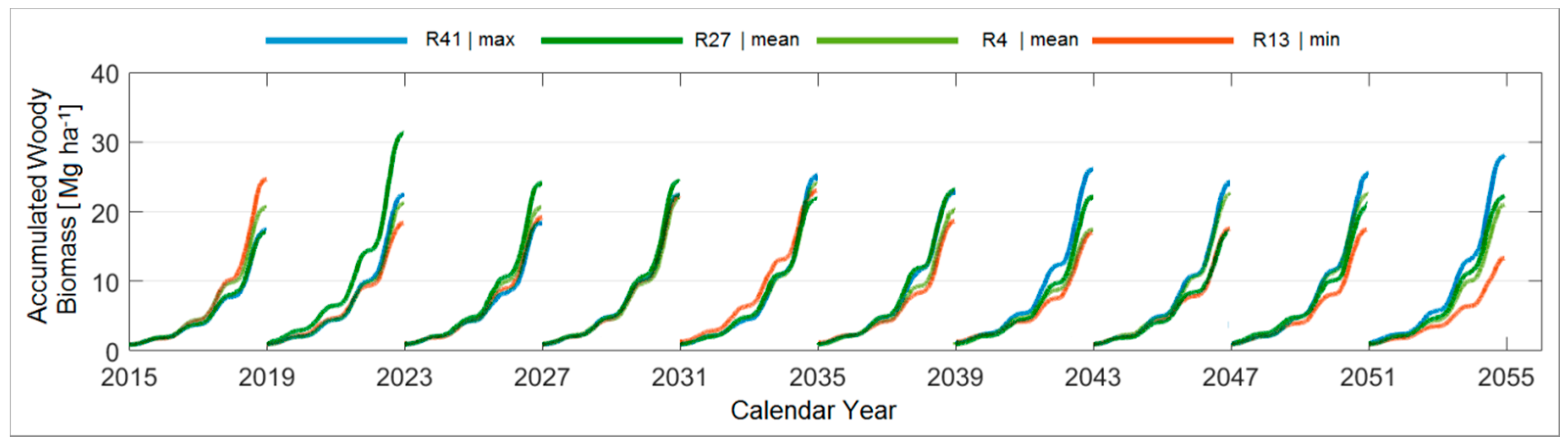
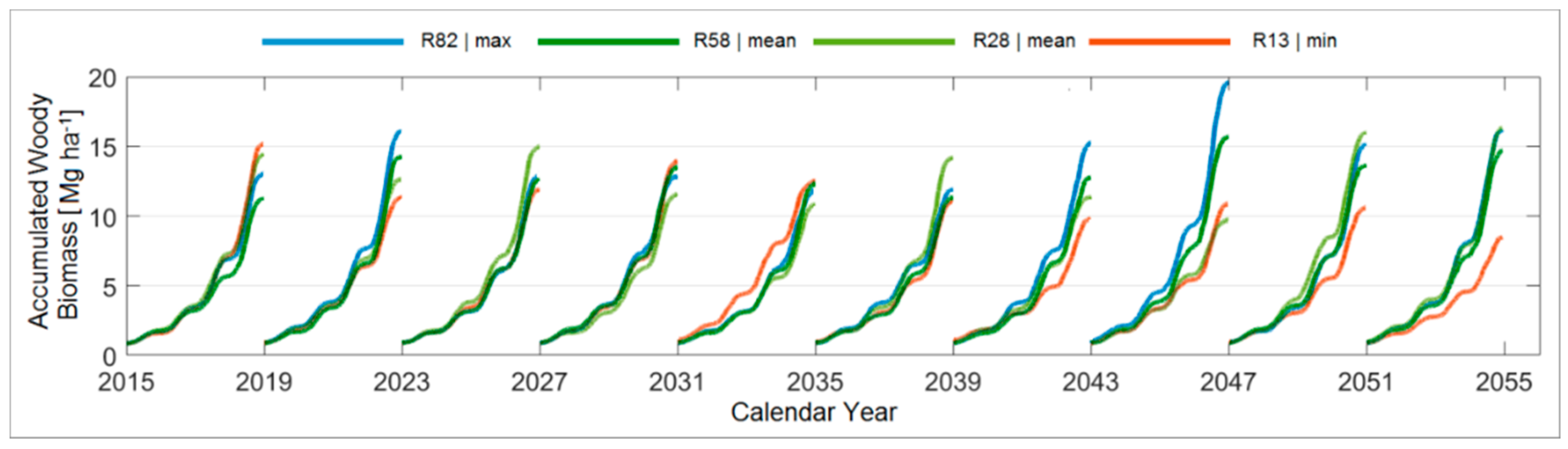
3.4. Modelled Woody Biomass under STAR 2K Weather Realisations
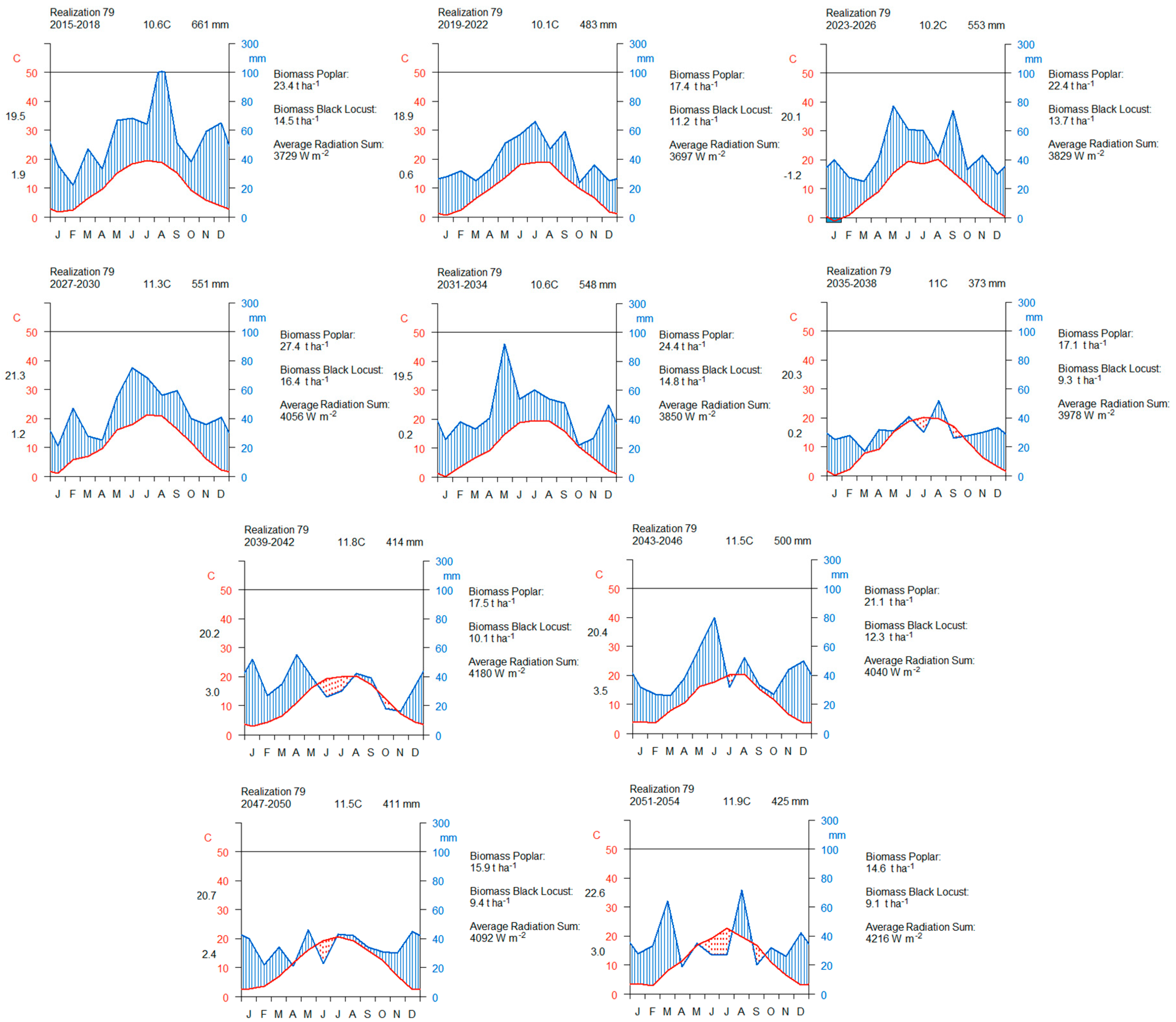
3.4.1. A Forty-Year Comparison with Respect to the Average Precipitation Sum
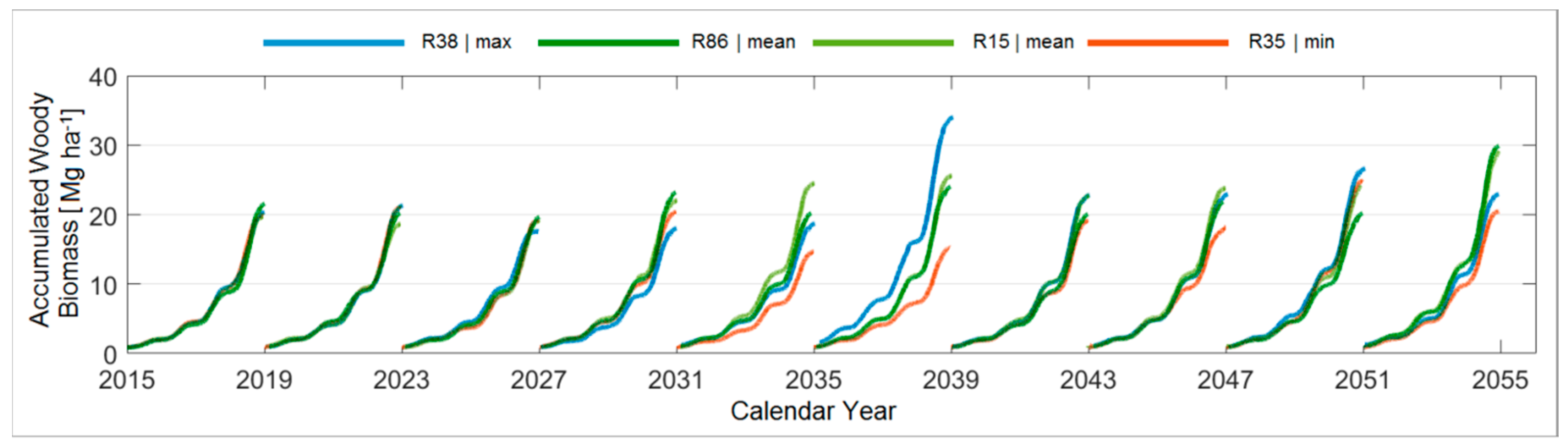
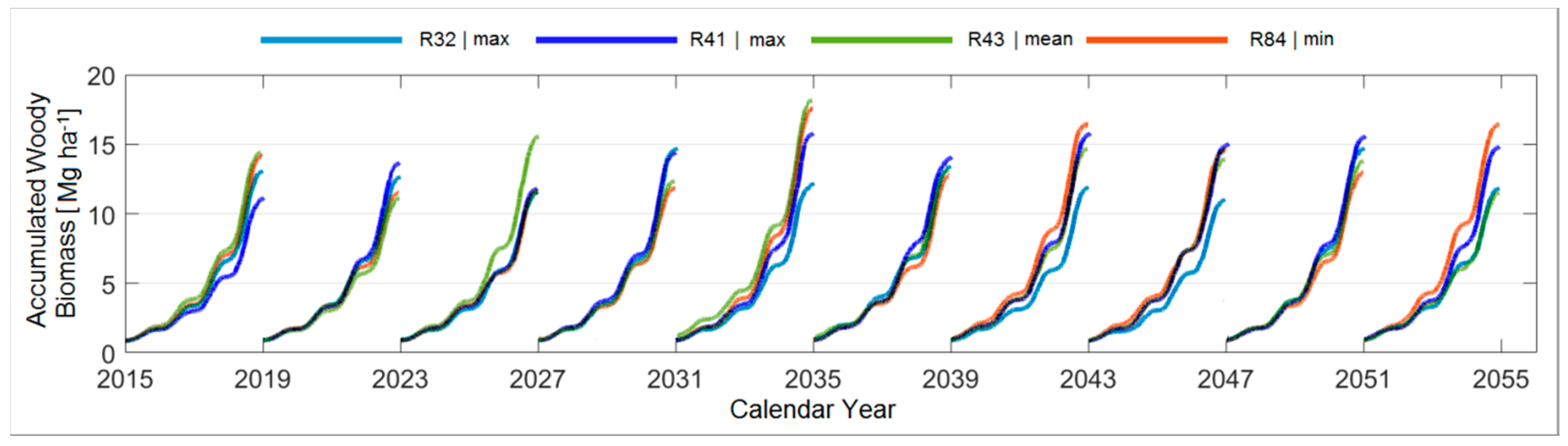
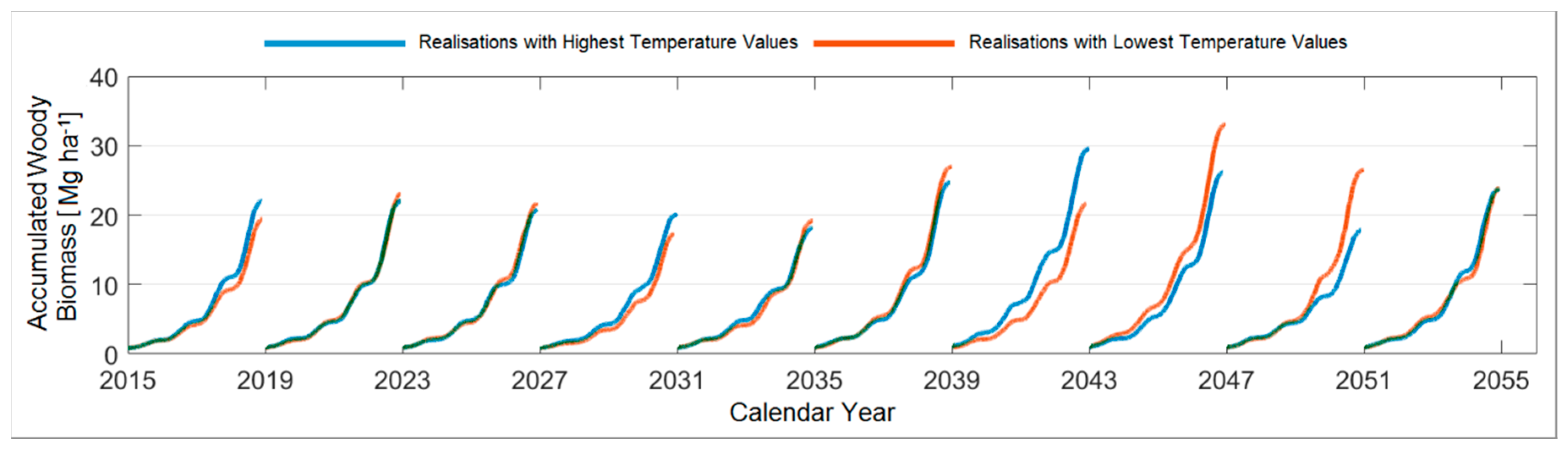
3.4.2. A Forty-Year Comparison with Respect to the Mean Temperature
3.4.3. Comparison between the Ten Year Growing Periods in Terms of Average Precipitation Sum
3.4.4. Comparison between the Ten Year Growing Periods in Terms of Mean Temperature
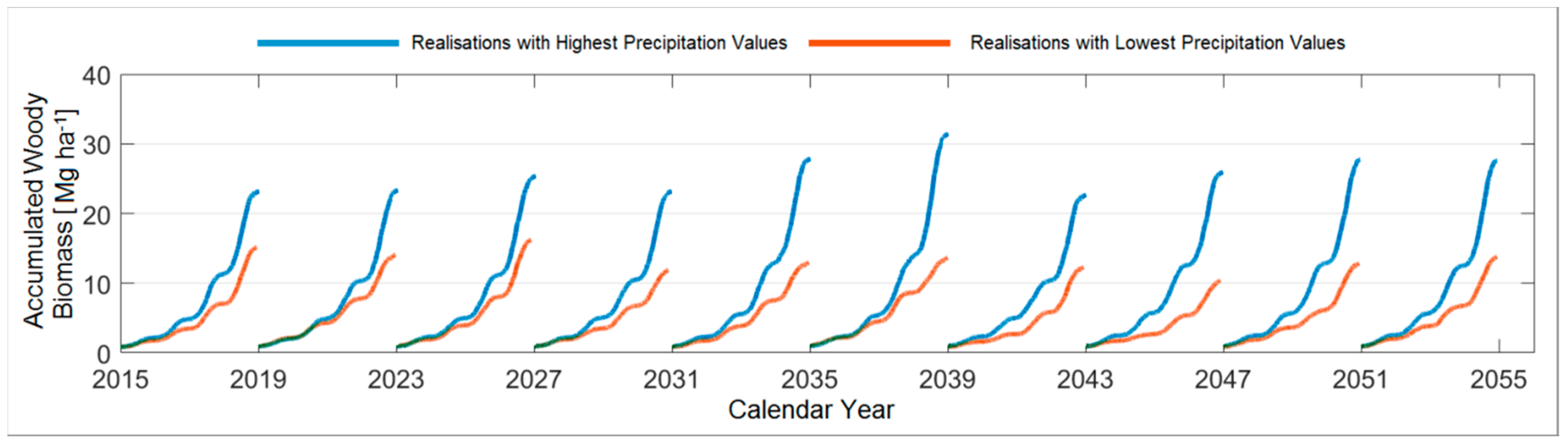
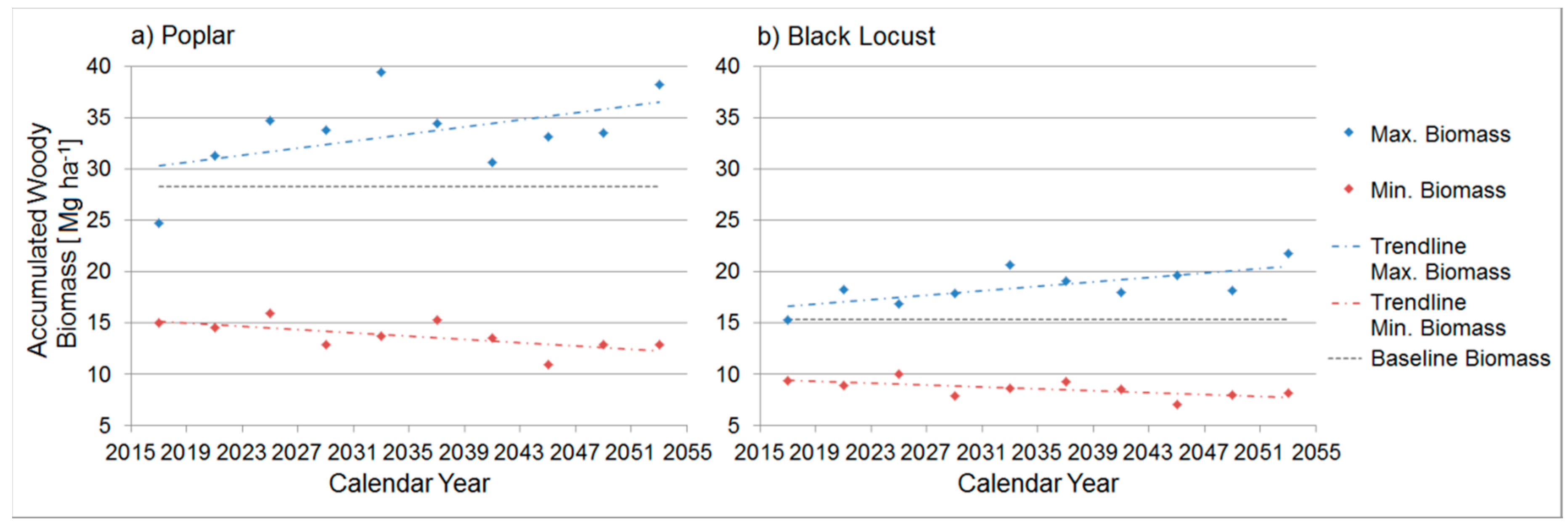
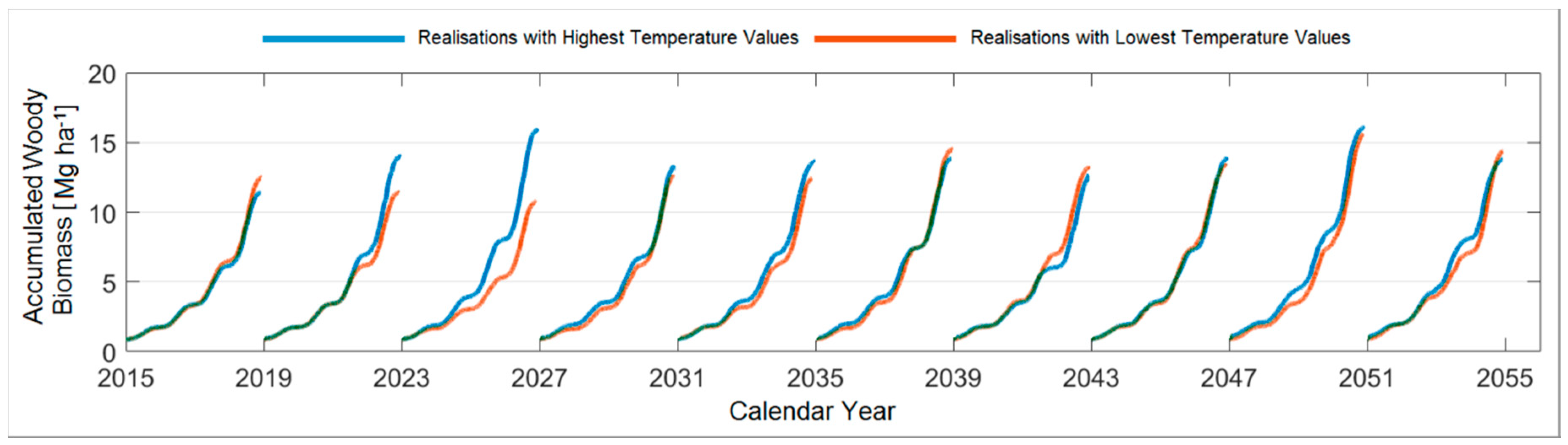
3.4.5. Comparison between the Ten Year Growing Periods in Terms of Accumulated Woody Biomass
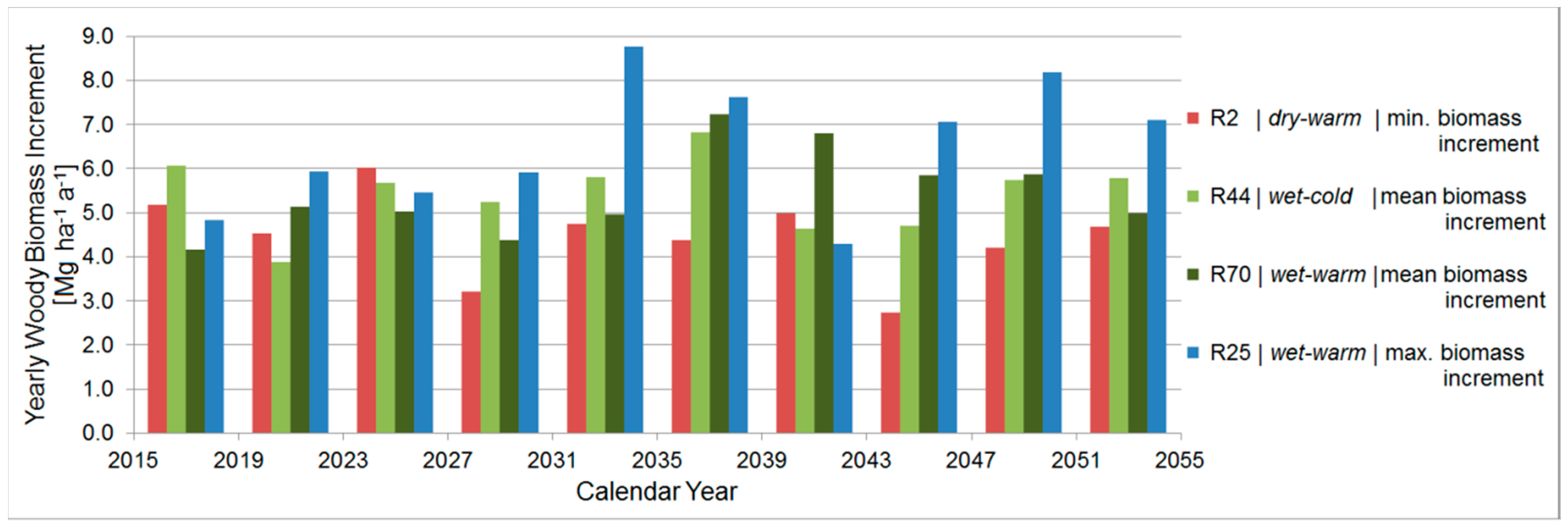
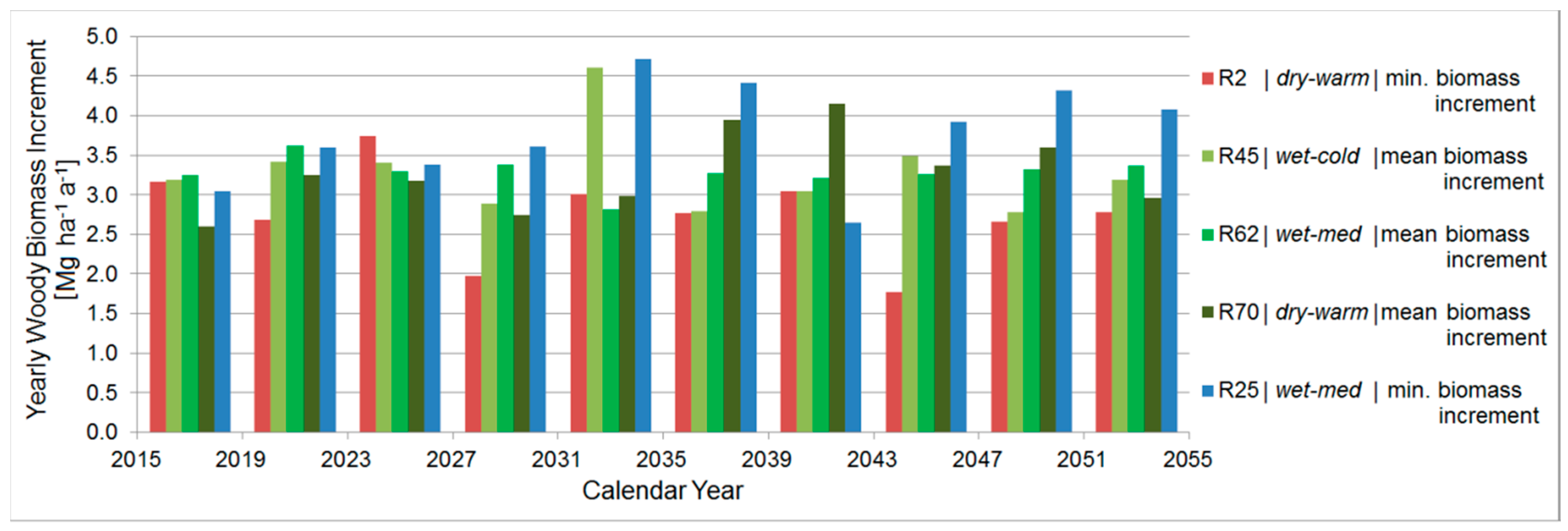
3.4.6. Comparison between the Ten Year Growing Periods in Terms of Woody Biomass Increment
4. Discussion
4.1. Parameterization and Validation of the Yield-SAFE Model
4.2. Evaluating the Woody Biomass Productivity under Prospective Climate Realisations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Unit | Tree Species | Value | Source |
|---|---|---|---|---|---|
| Tree Parameters | |||||
| Initial Conditions | |||||
| nShoots0 | Initial number of shoots per tree | tree−1 | Poplar | 0.3362 | Own data |
| Black Locust | 0.2520 | ||||
| Bt0 | Initial tree biomass | g tree−1 | Poplar | 100 | [39,41] |
| Black Locust | |||||
| LAt0 | Initial tree leaf area | m2 tree−1 | Poplar | 0 | [39,41] |
| Black Locust | |||||
| Parameters | |||||
| εt | Radiation use efficiency | g MJ−1 | Poplar | 0.2137 | Own data |
| Black Locust | 0.4820 | ||||
| kt | Light extinction coefficient | – | Poplar | 0.8 | [39,41] |
| Black Locust | |||||
| tt | The number of days after budburst at which the leaf area has reached 63.2% of its maximum leaf area LAssmax | d | Poplar | 10 | [39,41] |
| Black Locust | |||||
| LAssmax | Maximum leaf area for a single shoot | m2 | Poplar | 0.05 | [39,41] |
| Black Locust | 0.025 | [54] | |||
| nShootsmax | Maximum number of shoots per tree | tree−1 | Poplar | 10000 | [39,41] |
| Black Locust | |||||
| Kmain | Relative attrition rate of tree biomass | d−1 | Poplar | 10−4 | [39,41] |
| Black Locust | |||||
| γt | Transpiration coefficient of the trees | m3 kg−1 | Poplar | 0.35 | [41] |
| Black Locust | 0.42 | [54] | |||
| (pFcrit)t | Critical pF value for trees | log (cm) | Poplar | 4.0 | [39] |
| Black Locust | [54] | ||||
| (pFpwp)t | pF value at permanent wilting point | log (cm) | Poplar | 4.2 | [39] |
| Black Locust | |||||
| Tree Leaf Behaviour | |||||
| DOYbudburst, DOYleaffall | Day of year for budburst and leaf fall | DOY | Poplar | 105, 280 | [46] |
| Black Locust | 125, 310 | ||||
| Management Tree Density | |||||
| ρt | Planting density | trees ha−1 | Poplar | 8700 | [34] |
| Black Locust | |||||
| Soil Parameters | |||||
| Initial Conditions | |||||
| θ0 | Initial volumetric water content | m3 m−3 | Poplar | 0.552 | [39,41] |
| Black Locust | |||||
| Parameters | |||||
| δeva | Potential evaporation per unit energy | mm MJ−1 | Poplar | 0.15 | [41] |
| Black Locust | |||||
| D | Depth of the soil compartment | mm | Poplar | 1500 | [45] |
| Black Locust | |||||
| α | Van Genuchten parameter | – | Poplar | 0.0383 | [45] |
| Black Locust | |||||
| nsoil | Van Genuchten parameter | – | Poplar | 1.3774 | [45] |
| Black Locust | |||||
| δ | Parameter affecting the drainage rate below root zone | – | Poplar | 0.07 | [45] |
| Black Locust | |||||
| PWP | Permanent wilting point | log (cm) | Poplar | 4.2 | [39,41] |
| Black Locust | |||||
| (pFcrit)E | Critical pF value for evaporation | log (cm) | Poplar | 2.3 | [39,41] |
| Black Locust | |||||
| pFFC | Water tension at field capacity | log (cm) | Poplar | 2.3 | [39,41] |
| Black Locust | |||||
| Ks | Soil hydraulic conductivity at saturation | mm d−1 | Poplar | 60 | [45] |
| Black Locust | |||||
| θs | Saturated volumetric water content | m3 m−3 | Poplar | 0.403 | [45] |
| Black Locust | |||||
| θr | Residual volumetric water content | m3 m−3 | Poplar | 0.025 | [45] |
| Average Values | Reference Period | 2015–2018 | 2019–2022 | 2023–2026 | 2027–2030 | 2031–2034 | 2035–2038 | 2039–2042 | 2043–2046 | 2047–2050 | 2051–2054 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Poplar | |||||||||||
| Pmax.[mm] T [°C] * | 366 | 389 16.0 * (R31) | 435 15.9 * (R7) | 413 17.3 * (R43) | 378 16.8 * (R41) | 442 16.8 * (R98) | 450 16.9 * (R96) | 376 17.6 * (R60) | 406 16.8 * (R10) | 413 17.3 * (R78) | 416 17.6 * (R39) |
| Pmin.[mm] T [°C] * | 225 16.5 * (R26) | 201 17.5 * (R56) | 211 16.9 * (R11) | 184 17.5 * (R2) | 212 16.8 * (R10) | 205 17.2 * (R79) | 203 17.4 * (R69) | 176 17.9 * (R2) | 197 17.1 * (R8) | 190 18.3 * (R79) | |
| Black Locust | |||||||||||
| Pmax.[mm] T [°C] * | 395 | 400 15.8 * (R79) | 440 15.4 * (R7) | 423 16.6 * (R43) | 408 16.3 * (R50) | 459 16.3 * (R98) | 445 16.4 * (R96) | 390 17.1 * (R60) | 413 17.0 * (R82) | 422 16.9 * (R78) | 429 17.1 * (R39) |
| Pmin.[mm] T [°C] * | 226 16.1 * (R19) | 207 17.0 * (R56) | 222 16.8 * (R11) | 188 17.1 * (R2) | 205 16.6 * (R10) | 209 17.1 * (R79) | 192 17.3 * (R79) | 196 17.2 * (R2) | 188 17.7 * (R46) | 200 16.9 * (R13) | |
| Poplar | |||||||||||
| Tmax. [°C] P [mm] * | 16.6 | 17.5 384 * (R16) | 17.6 269, 304 * (R33, R69) | 18.0 273 * (R55) | 17.9 274 * (R98) | 18.2 235 * (R62) | 18.3 247, 271 * (R1, R17) | 18.3 311 * (R97) | 18.3 234, 306 * (R36, R81) | 18.6 227 * (R37) | 18.5 327 * (R47) |
| Tmin. [°C] P [mm] * | 15.2 368 * (R71) | 15.9 435, 317 * (R7, R79) | 16.1 294 * (R75) | 15.9 324 * (R1) | 16.1 362 * (R3) | 16.5 325, 373 * (R42, R95) | 16.5 325 * (R26) | 16.8 406, 330 * (R10, R94) | 16.4 329 * (R66) | 17.1 289 * (R91) | |
| Black Locust | |||||||||||
| Tmax. [°C] P [mm] * | 15.6 | 16.6 257, 257 * (R64, R74) | 17.1 336 * (R41) | 17.1 344, 342 * (R27, R83) | 17.4 278 * (R4) | 17.3 277 * (R96) | 17.4 246 * (R26, R65) | 17.6 248 * (R22) | 17.7 336 * (R4) | 17.8 291 * (R51) | 17.8 265 * (R73) |
| Tmin. [°C] P [mm] * | 14.6 391 * (R71) | 15.3 267 * (R84) | 15.6 280 * (R67) | 15.4 324 * (R1) | 15.2 377 * (R3) | 15.9 361 * (R81) | 16.0 341 * (R26) | 16.1 334 * (R31) | 16.0 350 * (R66) | 16.5 285 * (R91) | |
| Poplar | |||||||||||
| Rmax. [Wm−2] | 3119 | 2926 (R64) | 3002 (R53) | 3017 (R22) | 3016 (R4) | 3069 (R13) | 3133 (R17) | 3082 (R24) | 3165 (R100) | 3146 (R37) | 3158 (R79) |
| Rmin. [Wm−2] | 2497 (R71) | 2640 (R34) | 2709 (R60) | 2701 (R1) | 2593 (R3) | 2653 (R95) | 2741 (R26) | 2745 (R67) | 2789 (R3) | 2738 (R3) | |
| Black Locust | |||||||||||
| Rmax. [Wm−2] | 2966 | 2812 (R11) | 2932 (R51) | 2887 (R22) | 2913 (R4) | 2898 (R6) | 2952 (R46) | 2958 (R79) | 3045 (R100) | 2981 (R13) | 3003 (R79) |
| Rmin. [Wm−2] | 2379 (R71) | 2560 (R79) | 2538 (R67) | 2573 (R41) | 2419 (R3) | 2517 (R95) | 2621 (R8) | 2597 (R67) | 2632 (R3) | 2559 (R3) | |
Appendix B






References
- European Commission. Communication from The Commission to The European Parliament, The Council, The European Economic and Social Committee and The Committee of The Regions Energy Roadmap 2050. /* COM/2011/0885 final */. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=celex:52011DC0885 (accessed on 27 April 2018).
- Strelher, A. Technologies of wood combustion. Ecol. Eng. 2000, 16, 25–40. [Google Scholar] [CrossRef]
- BMWi Bekanntmachung über die Förderung von Forschung und Entwicklung zur kosten- und energieeffizienten Nutzung von Biomasse im Strom- und Wärmemarkt „Energetische Biomassenutzung“. BAnz AT 21.07.2015 B1, 1–7. Available online: https://www.ptj.de/lw_resource/datapool/systemfiles/cbox/604/live/lw_bekdoc/fbk_energetische_biomassenutzung.pdf (accessed on 27 April 2018).
- Abbasi, T.; Abbasi, S.A. Biomass energy and the environmental impacts associated with its production and utilization. Renew. Sust. Energ. Rev. 2010, 14, 919–937. [Google Scholar] [CrossRef]
- Evans, A.; Strezov, V.; Evans, T.J. Sustainability considerations for electricity generation from biomass. Renew. Sust. Energ. Rev. 2010, 14, 1419–1427. [Google Scholar] [CrossRef]
- Ceulemans, R.; Deraedt, W. Production physiology and growth potential of poplars under short-rotation forestry culture. Forest Ecol. Manag. 1999, 121, 9–23. [Google Scholar] [CrossRef]
- Hauck, S.; Knocke, T.; Wittkopf, S. Economic evaluation of short rotation coppice systems for energy from biomass—A review. Renew. Sust. Energ. Rev. 2014, 29, 435–448. [Google Scholar] [CrossRef]
- Carl, C.; Biber, P.; Landgraf, D.; Buras, A.; Pretzsch, H. Allometric models to predict aboveground woody biomass of black locust (Robinia pseudoacacia L.) in short rotation coppice in previous mining and agricultural areas in Germany. Forests 2017, 8, 328. [Google Scholar]
- Ceulemans, R.; McDonald, A.J.S.; Pereira, J.S.A. A comparison among eucalypt, poplar and willow characteristics with particular reference to a coppice, growth-modelling approach. Biomass Bioenerg. 1996, 11, 215–231. [Google Scholar] [CrossRef]
- Walle, I.V.; Camp, N.V.; Van de Casteele, L.; Verheyen, K.; Lemeur, R. Short-rotation forestry of birch, maple, poplar and willow in Flanders (Belgium) II. Energy production and CO2 emission reduction potential. Biomass Bioenerg. 2007, 31, 276–283. [Google Scholar]
- Sims, R.E.H.; Maiava, T.G.; Bullock, B.T. Short rotation coppice tree species selection for woody biomass production in New Zealand. Biomass Bioenerg. 2001, 20, 329–335. [Google Scholar] [CrossRef]
- Aravanopoulos, F.A.; Kimb, K.H.; Zsuffa, L. Genetic diversity of superior Salix clones selected for intensive forestry plantations. Biomass Bioenerg. 1999, 16, 249–255. [Google Scholar] [CrossRef]
- Zewdie, M.; Olsson, M.; Verwijst, T. Above-ground biomass production and allometric relations of Eucalyptus globulus Labill coppice plantations along a chronosequence in the central highlands of Ethiopia. Biomass Bioenerg. 2009, 33, 421–428. [Google Scholar] [CrossRef]
- Sochacki, S.J.; Harper, R.J.; Smettem, K.R.J. Estimation of woody biomass production from a short-rotation bio-energy system in semi-arid Australia. Biomass Bioenerg. 2007, 31, 608–616. [Google Scholar] [CrossRef]
- Van Vooren, L.; Reubens, B.; Broekx, S.; Pardon, P.; Reheul, D.; van Winsen, F.; Verheyen, K.; Wauters, E.; Lauwers, L. Greening and producing: An economic assessment framework for integrating trees in cropping systems. Agric. Syst. 2016, 148, 44–57. [Google Scholar] [CrossRef]
- Gerstengarbe, F.W.; Badeck, F.; Hattermann, F.; Krysanova, V.; Lahmer, W.; Lasch, P.; Stock, M.; Suckow, F.; Wechsung, F.; Werner, P.C. Studie zur klimatischen Entwicklung im Land Brandenburg bis 2055 und deren Auswirkungen auf den Wasserhaushalt, die Forst- und Landwirtschaft sowie die Ableitung erster Perspektiven. PIK 2003, 83, 77. [Google Scholar]
- Gädeke, A.; Pohle, I.; Koch, H.; Grünewald, U. Trend analysis for integrated regional climate change impact assessments in the Lusatian river catchments (North-Eastern Germany). Reg. Environ. Chang. 2017, 17, 1751–1762. [Google Scholar] [CrossRef]
- Rauthe, M.; Steiner, H.; Riediger, U.; Mazurkiewicz, A.; Gratzki, A. A Central European precipitation climatology—Part I: Generation and validation of a high-resolution gridded daily data set (HYRAS). Meteorol. Zeitschrift 2013, 22, 235–256. [Google Scholar] [CrossRef]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Bøssing Christensen, O.; Bouwer, L.M.; Braun, A.; Colette, A.; Déqué, M.; Georgievski, G.; et al. EURO-CORDEX: New high-resolution climate change projections for European impact research. Reg. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Christensen, J.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.; Kwon, W.T.; Laprise, R.; et al. Regional climate projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K., Tignor, M., Miller, H., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Ruane, A.C.; Hudson, N.I.; Asseng, S.; Camarrano, D.; Ewert, F.; Martre, P.; Boote, K.J.; Thorburn, P.J.; Aggarwal, P.K.; Angulo, C.; et al. Multi-wheat-model ensemble responses to interannual climate variability. Environ. Modell. Softw. 2016, 81, 86–101. [Google Scholar] [CrossRef] [Green Version]
- Horemans, J.A.; van Gaelen, H.; Raes, D.; Zenone, T.; Ceulemans, R. Can the agricultural AquaCrop model simulate water use and yield of a poplar short-rotation coppice? GCB Bioenergy 2017, 9, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- de Vries, W.; Posch, M.; Simpson, D.; Reinds, G.J. Modelling long-term impacts of changes in climate, nitrogen deposition and ozone exposure on carbon sequestration of European forest ecosystems. Sci. Total Environ. 2017, 605–606, 1097–1116. [Google Scholar] [CrossRef] [PubMed]
- Lasch, P.; Kollas, C.; Rock, J.; Suckow, F. Potentials and impacts of short-rotation coppice plantation with aspen in Eastern Germany under conditions of climate change. Reg. Environ. Chang. 2010, 10, 83–94. [Google Scholar] [CrossRef]
- Lasch-Born, P.; Suckow, F.; Gutsch, M.; Reyer, C.; Hauf, I.; Murawski, A.; Pilz, T. Forests under climate change: Potential risks and opportunities. Meteorol. Zeitschrift 2015, 24, 157–172. [Google Scholar] [CrossRef]
- Reyer, C.; Lasch-Born, P.; Suckow, F.; Gutsch, M.; Murawski, A.; Pilz, T. Projections of regional changes in forest net primary productivity for different tree species in Europe driven by climate change and carbon dioxide. Ann. Forest Sci. 2014, 71, 211–225. [Google Scholar] [CrossRef]
- Wang, W.J.; He, H.S.; Thompson, F.R.; Fraser, J.S.; Dijak, W.D. Changes in forest biomass and tree species distribution under climate change in the northeastern United States. Landsc. Ecol. 2017, 32, 1399–1413. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Delmotte, F.M.; Villar, M.; Delay, D.; Boudouresque, E.; Petit, J.M.; Marron, N.; Bréchet, C.; Brignolas, F. Productivity, leaf traits and carbon isotope discrimination in 29 Populus deltoides × P. nigra clones. New Phytol. 2005, 167, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Calfapietra, C.; Gielen, B.; Karnosky, D.; Ceulemans, R.; Mugnozza, G.S. Response and potential of agroforestry crops under global change. Environ. Pollut. 2010, 158, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Medlyn, B.E.; Duursma, R.A.; Zeppel, M.J.B. Forest productivity under climate change: A checklist for evaluating model studies. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 332–355. [Google Scholar] [CrossRef]
- van der Werf, W.; Keesman, K.; Burgess, P.; Graves, A.; Pilbeam, D.; Incoll, L.D.; Metselaar, K.; Mayus, M.; Stappers, R.; van Keulen, H.; et al. Yield-SAFE: A parameter-sparse, process-based dynamic model for predicting resource capture, growth, and production in agroforestry systems. Ecol. Eng. 2007, 29, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Werner, P.C.; Gerstengarbe, F.W. Proposal for the development of climate scenarios. Clim. Res. 1997, 8, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Orlowsky, B.; Gerstengarbe, F.W.; Werner, P.C. A resampling scheme for regional climate simulations and its performance compared to a dynamical RCM. Theor. Appl. Climatol. 2008, 92, 209–223. [Google Scholar] [CrossRef]
- Kanzler, M.; Böhm, C. Nachhaltige Erzeugung von Energieholz in Agroforstsystemen (AgroForstEnergie II)—Abschlussbericht Teilvorhaben 2: Bodenschutz und Bodenfruchtbarkeit, Wasserhaushalt und Mikroklima; Brandenburgische Technische Universität Cottbus-Senftenberg: Cottbus, Germany, 2016. [Google Scholar]
- Bielefeldt, J.; Bolte, A.; Busch, G.; Dohrenbusch, A.; Kroiher, F.; Lamersdorf, N.; Schulz, U.; Stoll, B. Energieholzproduktion in der Landwirtschaft. Chancen und Risiken aus Sicht der Natur-und Umweltschutzes. NABU Bundesverb. 2008, 1, 17–19. [Google Scholar]
- Verwijst, T.; Telenius, B. Biomass estimation procedures in short rotation forestry. Forest Ecol. Manag. 1999, 121, 137–146. [Google Scholar] [CrossRef]
- Zianis, D.; Mencuccini, M. On simplifying allometric analyses of forest biomass. Forest Ecol. Manag. 2003, 187, 311–332. [Google Scholar] [CrossRef]
- Böhm, C.; Quinkenstein, A.; Freese, D. Yield prediction of young black locust (Robinia pseudoacacia L.) plantations for woody biomass production using allometric relations. Ann. For. Res. 2011, 54, 215–227. [Google Scholar]
- Keesman, K.J.; Graves, A.; van der Werf, W.; Burgess, P.; Palma, J.; Dupraz, C.; van Keulen, H. A system identification approach for developing and parameterising an agroforestry system model under constrained availability of data. Environ. Modell. Softw. 2011, 26, 1540–1553. [Google Scholar] [CrossRef]
- Stappers, R.; Keesman, K.J.; van der Werf, W. The SAFE-RESULT Equations: An Agro-Forestry Model; Technical Report for Wageningen University: Wageningen, The Netherlands, 2003. [Google Scholar]
- Graves, A.R.; Burgess, P.J.; Palma, J.; Keesman, K.J.; van der Werf, W.; Dupraz, C.; van Keulen, H.; Herzog, F.; Mayus, M. Implementation and calibration of the parameter-sparse Yield-SAFE model to predict production and land equivalent ratio in mixed tree and crop systems under two contrasting production situations in Europe. Ecol. Model. 2010, 221, 1744–1756. [Google Scholar] [CrossRef] [Green Version]
- Burgess, P.; Graves, A.; Palma, J.; Herzog, F.; Keesman, K.; van der Werf, W. EU SAFE Project Deliverable 6.4: Parametrization of the Yield-SAFE Model and Its Use to Determine Yields at the Landscape Test Sites; Cranfield University-Institute of Water and Environment: Silsoe, UK, 2005. [Google Scholar]
- Palma, J.H.N.; Graves, A.R.; Bunce, R.G.H.; Burgess, P.J.; de Filippi, R.; Keesman, K.J.; van Keulen, H.; Liagre, F.; Mayus, M.; Reisner, Y.; et al. Modeling environmental benefits of silvoarable agroforestry in Europe. Agr. Ecosyst. Environ. 2007, 119, 320–334. [Google Scholar] [CrossRef] [Green Version]
- Palma, J.H.N.; Paulo, J.A.; Tome, M. Carbon sequestration of modern Quercus suber L. silvoarable agroforestry systems in Portugal: A YieldSAFE-based estimation. Agroforest. Syst. 2004, 88, 791–801. [Google Scholar] [CrossRef]
- Wösten, J.H.M.; Lilly, A.; Nemes, A.; Le Bas, C. Development and use of a database of hydraulic properties of European soils. Geoderma 1999, 90, 169–185. [Google Scholar] [CrossRef]
- Küppers, M.; Schmitt, D.; Liner, S.; Böhm, C.; Kanzler, M.; Veste, M. Photosynthetic characteristics and simulation of annual leaf carbon gains of hybrid poplar (Populus nigra L. × P. maximowiczii Henry) and black locust (Robinia pseudoacacia L.) in a temperate agroforestry system. Agroforest. Syst. 2017, 1–20. [Google Scholar] [CrossRef]
- Page, J.K. The estimation of monthly mean values of daily total short-wave radiation of vertical and inclined surface from sunshine records for latitudes 40° N-40° S. In Proceedings of UN Conference on New Sources of Energy; Ruiz Elizondo, J., Ed.; United Nations: New York, NY, USA, 1964. [Google Scholar]
- Gädeke, A.; Hölzel, H.; Koch, H.; Pohle, I.; Grünewald, U. Analysis of uncertainties in the hydrological response of a model-based climate change impact assessment in a subcatchment of the Spree River, Germany. Hydrol. Process. 2014, 28, 3978–3998. [Google Scholar] [CrossRef]
- Pohle, I.; Koch, H.; Conradt, T.; Gädeke, A.; Grünewald, U. Potential impacts of climate change and regional anthropogenic activities in Central European mesoscale catchments. Hydrolog. Sci. J. 2015, 60, 912–928. [Google Scholar] [CrossRef]
- Weemstra, M.; Eilmann, B.; Sass-Klaassen, U.G.W.; Sterck, F.J. Summer droughts limit tree growth across 10 temperate species on a productive forest site. Forest Ecol. Manag. 2013, 306, 142–149. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought matters—Declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in north-eastern Germany. Forest Ecol. and Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Veste, M.; Schillem, S.; Böhm, C. Baumarten für die Agrarholzproduktion. In Agrarholz—Schnellwachsende Bäume in der Landwirtschaft; Veste, M., Böhm, C., Eds.; Springer: Heidelberg, Germany, 2018. [Google Scholar]
- Veste, M.; Kriebitzsch, W.U. Einfluss von Trockenstress auf Photosynthese, Transpiration und Wachstum junger Robinien (Robinia pseudoacacia L.). Forstarchiv 2013, 84, 35–42. [Google Scholar]
- Mantovani, D.; Veste, M.; Böhm, C.; Vignudelli, M.; Freese, D. Spatial and temporal variation of growth performance and plant water status of black locust (Robinia pseudoacacia L.) in agroforestry systems in Lower Lusatia (Germany). iForest 2015, 8, 743–747. [Google Scholar] [CrossRef]
- Mantovani, D.; Veste, M.; Boldt-Burisch, K.; Fritsch, S.; Koning, L.; Freese, D. Carbon allocation, nodulation, and biological nitrogen fixation of black locust (Robinia pseudoacacia L.) under soil water limitation. Ann. For. Res. 2015, 58, 259–274. [Google Scholar] [CrossRef]
- Veste, M.; Halke, C. Ökophysiologische Plastizität der Photosynthese von Robinien (Robinia pseudoacacia L.) und Hybrid-Pappeln (Populus nigra L. × P. maximowiczii Henry) bei Hitzestress und Sommertrockenheit in der Niederlausitz. In Proceedings 5. Agroforstforum; Böhm, C., Ed.; Brandenburgische Technische Universität Cottbus-Senftenberg IKMZ—Universitätsbibliothek: Senftenberg, Germany, 2017. [Google Scholar]
- Mantovani, D.; Veste, M.; Freese, D. Black locust (Robinia pseudoacacia L.) ecophysiological and morphological adaptations to drought and their consequence on biomass production and water use efficiency. N. Z. J. For. Sci. 2014, 44, 29. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Müller, A.; Küchler, W. Possible impacts of climate change on natural vegetation in Saxony (Germany). Int. J. Biometeorol. 2005, 50, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavska, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, D.A. Standortbasierte Ertragsmodellierung von Pappel-und Weidenklonen in Kurzumtriebsplantagen. Available online: http://nbn-resolving.de/urn:nbn:de:bsz:14-qucosa-144670 (accessed on 5 December 2016).
- Leuzinger, S.; Zotz, G.; Asshoff, R.; Körner, C. Responses of deciduous forest trees to severe drought in Central Europe. Tree Physiol. 2015, 25, 641–650. [Google Scholar] [CrossRef]
- Gessler, A.; Keitel, C.; Kreuzwieser, J.; Matyssek, R.; Seiler, W.; Rennenberg, H. Potential risks for European beech (Fagus sylvatica L.) in a changing climate. Trees 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Kriebitzsch, W.U.; Veste, M. Bedeutung trockener Sommer für die Photosynthese und Transpiration von verschiedenen Herkünften der Rotbuche (Fagus sylvatica L.). Landbauforschung 2012, 62, 193–209. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Bolte, A.; Madsen, P.; Derkyi, M.A.A.; Stanturf, J.A. Forest adaptation and restoration under global change—Concept and status of an IUFRO Task Force. Flora Mediterr. 2017, 27, 6. [Google Scholar]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C.; Basler, D. Phenology under global warming. Science 2010, 327, 1461–1462. [Google Scholar]
- Vitasse, Y.; Basler, D. What role for photoperiod in the bud burst phenology of European beech. Eur. J. Forest Res. 2013, 132, 1–8. [Google Scholar] [CrossRef]
- Malyshev, A.V.; Henry, H.A.; Bolte, A.; Arfin Khan, M.A.; Kreyling, J. Temporal photoperiodic sensitivity and forcing requirements for budburst in temperate tree seedlings. Agr. Forest. Meteorol. 2018, 248, 82–90. [Google Scholar] [CrossRef]
- Lasch, P.; Lindner, M.; Erhard, M.; Suckow, F.; Wenzel, A. Regional impact assessment on forest structure and functions under climate change–the Brandenburg case study. Forest Ecol. Manag. 2002, 162, 73–86. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Rosenzweig, C.; Jones, J.W.; Hatfield, J.L.; Duane, A.C.; Boote, K.J.; Thorburn, P.J.; Rötter, R.P.; Cammarano, D.; et al. Uncertainty in simulating wheat yields under climate change. Nat. Clim. Chang. 2013, 3, 827–832. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schehaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Krysanova, V.; Österle, H.; Hattermann, F.F. Simulation of spatiotemporal dynamics of water fluxes in Germany under climate change. Hydrol. Processes 2010, 24, 3289–3306. [Google Scholar] [CrossRef]









| Species | Biomass after 1 Year (Mg ha−1) | Biomass after 2 Years (Mg ha−1) | Biomass after 3 Years (Mg ha−1) | Biomass after 4 Years (Mg ha−1) |
|---|---|---|---|---|
| Poplar | 0.4 ± 0.1 (n = 333) | 2.0 ± 0.5 (n = 150) | 12.9 ± 0.1 (n = 242) | 28.2 ± 2.7 (n = 50) |
| Black Locust | 0.1 ± 0.1 (n = 360) | 3.6 ± 0.5 (n = 306) | 9.1 ± 1.4 (n = 152) | 15.3 ± 3.4 (n = 219) |
| Average Values for the Vegetation Period | Poplar | Black Locust | ||||||
|---|---|---|---|---|---|---|---|---|
| Base Period (1974–2014) | Min. (2015–2054) | Mean (2015–2054) | Max. (2015–2054) | Base Period (1974–2014) | Min. (2015–2054) | Mean (2015–2054) | Max. (2015–2054) | |
| P [mm] T [°C] * | 324 | 254 17.3 * (R13) | 296 17.1, 17.4 * (R4, R27) | 327 17.5 * (R41) | 336 | 271 16.7 * (R13) | 303 17.1, 17.3 * (R28, R58) | 335 17.2 * (R82) |
| T [°C] P [mm] * | 16.0 | 17.0 283 * (R35) | 17.2 299, 299 * (R15, R86) | 17.5 283 * (R38) | 15.3 | 16.2 315 * (R84) | 16.6 298 * (R43) | 16.9 298, 332 * (R32, R41) |
| Accumulated Woody Biomass [Mg ha−1] | 2015–2018 | 2019–2022 | 2023–2026 | 2027–2030 | 2031–2034 | 2035–2038 | 2039–2042 | 2043–2046 | 2047–2050 | 2051–2054 |
|---|---|---|---|---|---|---|---|---|---|---|
| Poplar | ||||||||||
| Max. | 24.7 | 31.3 | 34.7 | 33.8 | 39.4 | 34.5 | 30.6 | 33.1 | 33.5 | 38.8 |
| Realisation | (R13) | (R27) | (R6) | (R49) | (R100) | (R37) | (R75) | (R82) | (R75) | (R62) |
| Min. | 15.0 | 14.6 | 15.9 | 12.9 | 13.7 | 15.3 | 13.5 | 10.9 | 12.9 | 12.9 |
| Realisation | (R26) * | (R56) * | (R18) | (R2) * | (R10) * | (R35) | (R69) * | (R2) * | (R16) | (R90) |
| Black Locust | ||||||||||
| Max. | 15.3 | 18.3 | 16.8 | 17.9 | 20.6 | 19.1 | 18.0 | 19.61 | 18.2 | 21.8 |
| Realisation | (R51) | (R27) | (R6) | (R74) | (R100) | (R96) * | (R75) | (R82) * | (R75) | (R63) |
| Min. | 9.4 | 8.9 | 10.0 | 7.9 | 8.6 | 9.3 | 8.5 | 7.1 | 8.0 | 8.2 |
| Realisation | (R19) * | (R56) * | (R18) | (R2) * | (R10) * | (R79) * | (R87) | (R2) * | (R16) | (R24) |
| Average Woody Biomass Increment [Mg ha−1 a−1] | Projected Period (2015–2054) | R2 | R44 | R45 | R70 | R62 | R25 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Poplar | Black Locust | Poplar | Black Locust | Poplar | Black Locust | Poplar | Black Locust | Black Locust | Poplar | Black Locust | |
| - | - | 4.5 | 2.8 | 5.4 | 3.3 | 5.4 | 3.3 | 3.3 | 6.5 | 3.8 | |
| P [mm a−1] | 296 | 303 | 270 | 279 | 313 | 316 | 304 | 301 | 310 | 306 | 313 |
| T [°C a−1] | 17.2 | 16.6 | 17.3 | 16.9 | 17.1 | 16.5 | 17.3 | 16.7 | 16.6 | 17.3 | 16.6 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seserman, D.-M.; Pohle, I.; Veste, M.; Freese, D. Simulating Climate Change Impacts on Hybrid-Poplar and Black Locust Short Rotation Coppices. Forests 2018, 9, 419. https://doi.org/10.3390/f9070419
Seserman D-M, Pohle I, Veste M, Freese D. Simulating Climate Change Impacts on Hybrid-Poplar and Black Locust Short Rotation Coppices. Forests. 2018; 9(7):419. https://doi.org/10.3390/f9070419
Chicago/Turabian StyleSeserman, Diana-Maria, Ina Pohle, Maik Veste, and Dirk Freese. 2018. "Simulating Climate Change Impacts on Hybrid-Poplar and Black Locust Short Rotation Coppices" Forests 9, no. 7: 419. https://doi.org/10.3390/f9070419
APA StyleSeserman, D. -M., Pohle, I., Veste, M., & Freese, D. (2018). Simulating Climate Change Impacts on Hybrid-Poplar and Black Locust Short Rotation Coppices. Forests, 9(7), 419. https://doi.org/10.3390/f9070419