Water Balance of Mediterranean Quercus ilex L. and Pinus halepensis Mill. Forests in Semiarid Climates: A Review in A Climate Change Context


 , ,
, ,
Abstract
:1. Introduction
2. Plant Functional Characteristics and Strategy to Cope with Drought
3. Water Balance in Quercus ilex Forests
4. Water Balance in Pinus halepensis Forests
5. The Role of Mediterranean Forests in Aquifer Recharge for Semiarid Areas
6. Forest Management Effects on the Transpiration of Mixed Forests
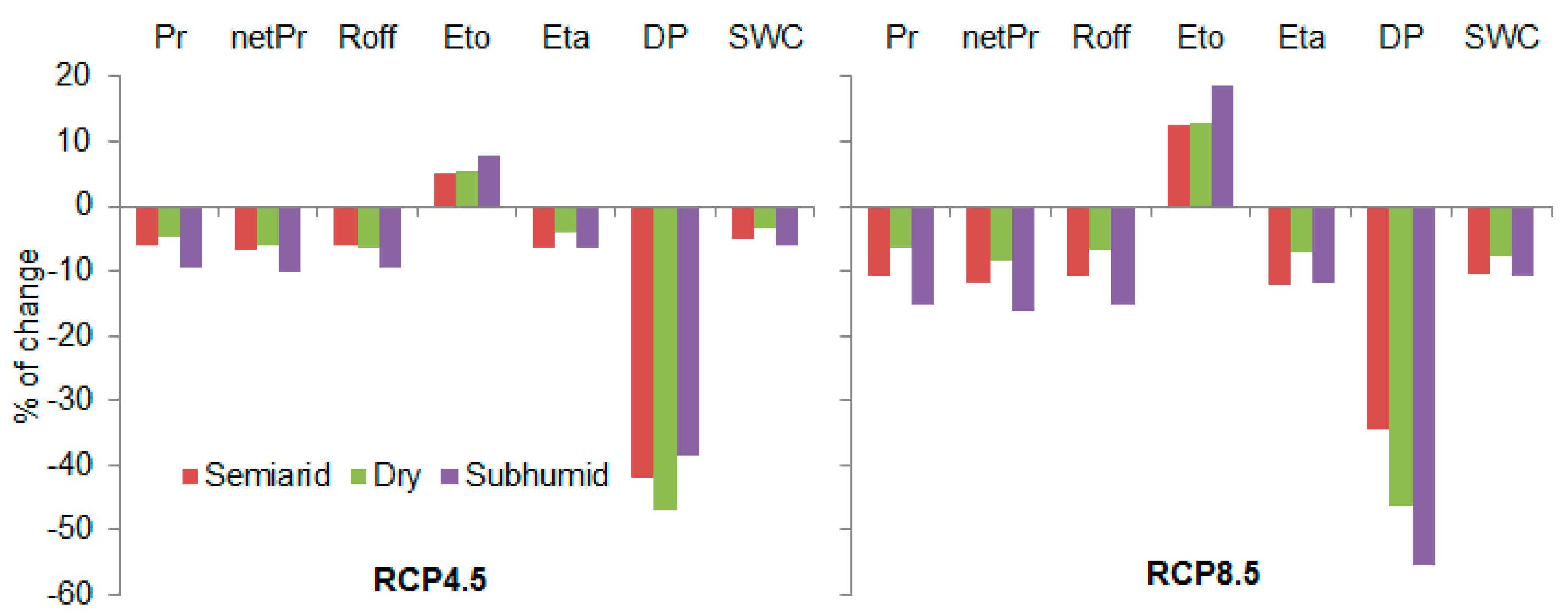
7. Future Scenarios in a Global Change Context
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellison, D.; Morris, C.E.; Locatellu, B.; Sheil, D.; Cohen, J.; Murdiyarso, D.; Gutierrez, V.; van Noordwijk, M.; Creed, I.F.; Pokorny, J.; et al. Trees, forests and water: Cool insigths for a hot world. Glob. Environ. Chang. 2017, 43, 51–61. [Google Scholar] [CrossRef]
- Rohatyn, S.; Rotenberg, E.; Ramati, E.; Tatarinov, F.; Tas, E.; Yakir, D. Differential impacts of land use and precipitation on ‘Ecosystem Water Yield’. Water Resour. Res. 2018. [Google Scholar] [CrossRef]
- Helman, D.; Lensky, I.M.; Osem, Y.; Rohatyn, S.; Rotenberg, E.; Yakir, D. A biophysical approach using water deficit factor for daily estimations of evapotranspiration and CO2 uptake in Mediterranean environments. Biogeosciences 2017, 14, 3909–3926. [Google Scholar] [CrossRef]
- Jackson, R.B.; Jobbagy, E.G.; Avissar, R.; Roy, S.B.; Barrett, D.J.; Cook, C.W.; Farley, K.A.; le Maitre, D.C.; McCarl, B.A.; Murray, B.C. Trading water for carbon with biological carbon sequestration. Science 2005, 310, 1944–1947. [Google Scholar] [CrossRef] [PubMed]
- Farley, K.A.; Jobbagy, E.G.; Jackson, R.B. Effects of afforestation on water yield: A global synthesis with implications for policy. Glob. Chang. Biol. 2005, 11, 1565–1576. [Google Scholar] [CrossRef]
- Muñoz-Villers, L.E.; Holwerda, F.; Gómez-Cárdenas, M.; Equihua, M.; Asbjornsen, H.; Bruijnzeel, L.A.; Marín-Castro, B.E.; Tobón, C. Water balances of old-growth and regenerating montane cloud forests in central Veracruz, Mexico. J. Hydrol. 2012, 462–463, 53–66. [Google Scholar] [CrossRef]
- Derak, M.; Cortina, J. Multi-criteria participative evaluation of Pinus halepensis plantations in a semiarid area of southeast Spain. Ecol. Indic. 2014, 43, 56–68. [Google Scholar] [CrossRef]
- Ilstedt, U.; Tobella, A.B.; Bazié, H.R.; Bayala, J.; Verbeeten, E.; Nyberg, G.; Sanou, J.; Benegas, L.; Murdiyarso, D.; Laudon, H.; et al. Intermediate tree cover can maximize groundwater recharge in the seasonally dry tropics. Sci. Rep. 2016, 6, 21930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, V.R.; Field, C.B.; Dokken, D.J.; Mastrandrea, M.D.; Mach, K.J.; Bilir, T.E.; Chatterjee, M.; Ebi, K.L.; Estrada, Y.O.; Genova, R.C.; et al. Climate Change 2014: Impacts, Adaptation, and Vulnerability—Part B: Regional Aspects—Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, New York, NY, USA, 2014; p. 688. [Google Scholar]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Hartmann, H.; Schuld, B.; Sanders, T.G.; Macinnis-Ng, C.; Boehmer, H.J.; Allen, C.D.; Bolte, A.; Crowther, T.W.; Hansen, M.C.; Medlyn, B.E.; et al. Monitoring global tree mortality patterns and trends. Report from the VW symposium ‘Crossing scales and disciplines to identify global trends of tree mortality as indicators of forest health’. New Phytol. 2018, 217, 984–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, L08707. [Google Scholar] [CrossRef]
- Herrera, C.M. Historical effects and sorting processes as explanations for contemporary ecological patterns; character syndromes in Mediterranean woody plants. Am. Nat. 1992, 140, 421–446. [Google Scholar] [CrossRef]
- Poyatos, R.; Aguadé, D.; Gaiano, L.; Mencuccini, M.; Martínez-Vilalta, J. Drought-induced defoliation and long periods of near-zero gas Exchange play a key role in accentuating metabolic decline of Scots pine. New Phytol. 2013, 200, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Grossiord, C.; Forner, A.; Gessler, A.; Granier, A.; Pollastrini, M.; Valladares, F.; Bonal, D. Influence of species interactions on transpiration of Mediterranean tree species during a summer drought. Eur. J. For. Restaur. 2015, 134, 365–376. [Google Scholar] [CrossRef]
- De la Torre, J.R. Distribución y características de las masas forestales españolas. Fundación Dialnet 1990, 1, 11–30. [Google Scholar]
- Zavala, M.A. A model of stand dynamics for holm oak-Aleppo pine forests. In Ecology of Mediterranean Evergreen Oak Forests; Rodà, F., Retana, J., Gracia, C.A., Bellot, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 105–117. [Google Scholar]
- Hernández, E.I.; Vilagrosa, A.; Pausas, J.G.; Bellot, J. Morphological traits and water use strategies in seedlings of Mediterranean coexisting species. Plant Ecol. 2010, 207, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Peguero-Pina, J.J.; Sancho-Knapik, D.; Barrón, E.; Camarero, J.J.; Vilagrosa, A.; Gil-Pelegrín, E. Morphological and physiological divergences within Quercus ilex support the existence of different ecotypes depending on climatic dryness. Ann. Bot. 2014, 114, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Vilagrosa, A.; Hernández, E.I.; Luis, V.C.; Cochard, H.; Pausas, J.G. Physiological differences explain the co-existence of different regeneration strategies in Mediterranean ecosystems. New Phytol. 2014, 201, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Peìrez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef] [Green Version]
- Cayuela, C.; Llorens, P.; Sánchez-Costa, E.; Levia, D.; Latron, J. Effect of biotic and abiotic factors on inter- and intra-event variability in stemflow rates in oak and pine stands in a Mediterranean mountain area. J. Hydrol. 2018, 560, 396–406. [Google Scholar] [CrossRef]
- Bellot, J.; Chirino, E. Hydrobal: An eco-hydrological modelling approach for assessing water balances in different vegetation types in semi-arid areas. Ecol. Model. 2013, 266, 30–41. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Hernández, E.I.; Matos, A.; Vallejo, V.R. Effects of a deep container on morpho-functional characteristics and root colonization in Quercus suber L. seedlings for reforestation in Mediterranean climate. For. Ecol. Manag. 2008, 256, 779–785. [Google Scholar] [CrossRef]
- Van der Plas, F.; Manning, P.; Soliveres, S.; Allan, E.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Ampoorter, E.; Baeten, L.; et al. Biotic homogenization can decrease landscape-scale forest multifunctionality. Proc. Natl. Acad. Sci. USA 2016, 113, 3557–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pemán, J.; Voltas, J.; Gil Pelegrín, E. Morphological and functional variability in the root system of Quercus ilex L. subject to confinement: Consequences for afforestation. Ann. For. Sci. 2006, 63, 425–430. [Google Scholar] [CrossRef]
- Bellot, J.; Escarre, A. Stemflow and throughfall determination in a resprouted Mediterranean holm-oak forest. Ann. For. Sci. 1998, 55, 847–865. [Google Scholar] [CrossRef] [Green Version]
- Helman, D.; Osem, Y.; Yakir, D.; Lensky, I.M. Relationships between climate, topography, water use and productivity in two key Mediterranean forest types with different water-use strategies. Agric. For. Meteorol. 2017, 232, 319–330. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses; Academic Press: New York, NY, USA, 1980; Volume 2. [Google Scholar]
- Tardieu, F.; Simonneau, T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.; Cobb, N.; Kolb, T.; Plaut, P.; West, J.A.; Williams, D.G.; Yepez, E.A. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Grossiord, C.; Sevanto, S.; Borrego, I.; Chen, A.M.; Collins, A.D.; Dickman, L.T.; Hudson, P.J.; McBranch, N.; Michaletz, S.T.; Pockman, W.T.; et al. Tree water dynamics in a drying and warming world. Plant Cell Environ. 2017, 40, 1861–1873. [Google Scholar] [CrossRef] [PubMed]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Garcia-Forner, N. Water potential regulation, stomatal behaviour and hydraulic transport under drought: Deconstructing the iso/anisohydric concept. Plant Cell Environ. 2016, 40, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Carmona, A.V. Estrategias De Resistencia Al Déficit Hídrico En Pistacia lentiscus L. Y Quercus coccifera L. Implicaciones En La Repoblación Forestal. Ph.D. Thesis, Universidad de Alicante, Alicante, Spain, 2002. [Google Scholar]
- Martínez-Ferri, E.; Balaguer, L.; Valladares, F.; Chico, J.M.; Manrique, E. Energy dissipation in drought-avoiding and drought tolerant tree species at midday during the Mediterranean summer. Tree Physiol. 2000, 20, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Chirino, E.; Bellot, J.; Sánchez, J.R. Daily sap flow rate as an indicator of drought avoidance mechanisms in five Mediterranean perennial species in semi-arid southeastern Spain. Trees 2011, 25, 593–606. [Google Scholar] [CrossRef]
- Ruiz-Yanetti, S.; Chirino, E.; Bellot, J. Daily whole-seedling transpiration determined by minilysimeters, allows the estimation of the water requirements of seedlings used for dryland afforestation. J. Arid Environ. 2016, 124, 341–351. [Google Scholar] [CrossRef]
- Klein, T.; Shpringer, I.; Fikler, B.; Elbaz, G.; Cohen, S.; Yakir, D. Relationships between stomatal regulation, water-use, and water-use efficiency of two coexisting key Mediterranean tree species. For. Ecol. Manag. 2013, 302, 34–42. [Google Scholar] [CrossRef]
- Liniger, H.P.; Weingartner, R. Mountain Forests and Their Role in Providing Freshwater Resources; CABI Publishing, in association with IUFRO (International Union for Forestry Research Organizations): Wallingford, UK; New York, NY, USA, 2000. [Google Scholar]
- Limousin, J.M.; Rambal, S.; Ourcival, J.M.; Rocheteau, A.; Joffre, R.; Rodríguez-Cortina, R. Long-term transpiration change with rainfall decline in a Mediterranean Quercus ilex forest. Glob. Chang. Biol. 2009, 15, 2163–2175. [Google Scholar] [CrossRef]
- Joffre, R.; Rambal, S. How tree cover influences the water balance of Mediterranean rangelands. Ecology 1993, 74, 570–582. [Google Scholar] [CrossRef]
- Ceballos, A.; Schnabel, S. Hydrological behaviour of a small catchment in the dehesa landuse system (Extremadura, SW Spain). J. Hydrol. 1998, 210, 146–160. [Google Scholar] [CrossRef]
- Campos, I.; González-Piqueras, J.; Carrara, A.; Villodre, J.; Calera, A. Estimation of total available water in the soil layer by integrating actual evapotranspiration data in a remote sensing-driven soil water balance. J. Hydrol. 2016, 534, 427–439. [Google Scholar] [CrossRef]
- Del Campo, A.; González-Sanchis, M.; Garcia-Prats, A.; Ceacero, C.J.; Lull, C. The impact of adaptive forest management on water fluxes and growth dynamics in a water-limited low-biomass oak coppice. Agric. For. Meteorol. 2018. submitted for publication. [Google Scholar]
- González-Sanchis, M.; Del Campo, A.D.; Molina, A.J.; Fernandes, T.J.G. Modeling adaptive forest management of a semi-arid Mediterranean Aleppo pine plantation. Ecol. Model. 2015, 308, 34–44. [Google Scholar] [CrossRef]
- Paço, T.A.; David, T.S.; Henriques, M.O.; Pereira, J.S.; Valente, F.; Banza, J.; Pereira, F.L.; Pinto, C.; David, J.S. Evapotranspiration from a Mediterranean evergreen oak savannah: The role of trees and pasture. J. Hydrol. 2009, 369, 98–106. [Google Scholar] [CrossRef]
- Infante, J.M.; Domingo, F.; Alés, R.F.; Joffre, R.; Rambal, S. Quercus ilex transpiration as affected by a prolonged drought period. Biol. Plant. 2003, 46, 49–55. [Google Scholar] [CrossRef]
- Schiller, G.; Cohen, Y. Water balance of Pinus halepensis Mill. afforestation in an arid region. For. Ecol. Manag. 1998, 105, 121–128. [Google Scholar] [CrossRef]
- Ungar, E.D.; Rotenberg, E.; Raz-Yaseef, N.; Cohen, S.; Yakir, D.; Schiller, G. Transpiration and annual water balance of Aleppo pine in a semiarid region: Implications for forest management. For. Ecol. Manag. 2013, 298, 39–51. [Google Scholar] [CrossRef]
- Raz-Yaseef, N.; Yakir, D.; Schiller, G.; Cohen, S. Dynamics of evapotranspiration partitioning in a semi-arid forest as affected by temporal rainfall patterns. Agric. For. Meteorol. 2012, 157, 77–85. [Google Scholar] [CrossRef]
- Barbeta, A.; Mejia-Chang, M.; Ogaya, R.; Voltas, J.; Dawson, T.E.; Penuelas, J. The combined effects of a long-term experimental drought and an extreme drought on the use of plant-water sources in a Mediterranean forest. Glob. Chang. Biol. 2015, 21, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, E.; Yakir, D. Distinct patterns of changes in surface energy budget associated with forestation in the semiarid region. Glob. Chang. Biol. 2011, 17, 1536–1548. [Google Scholar] [CrossRef]
- Tatarinov, F.; Rotenberg, E.; Maseyk, K.; Ogee, J.; Klein, T.; Yakir, D. Resilience to seasonal heat wave episodes in a Mediterranean pine forest. New Phytol. 2016, 210, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Helman, D.; Lensky, I.M.; Yakir, D.; Osem, Y. Forests growing under dry conditions have higher hydrological resilience to drought than do more humid forests. Glob. Chang. Biol. 2017, 23, 2801–2817. [Google Scholar] [CrossRef] [PubMed]
- Maseyk, K.S.; Lin, T.; Rotenberg, E.; Grünzweig, J.M.; Schwartz, A.; Yakir, D. Physiology-phenology interactions in a productive semi-arid pine forest. New Phytol. 2008, 178, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Yanetti, S. Respuesta A La Sequía De Especies Y Comunidades De Ambientes Contrastados: Comparación De Balances Hídricos. Ph.D. Thesis, University of Alicante, Alicante, Spain, 2017. [Google Scholar]
- Bellot, J.; Sanchez, J.R.; Chirino, E.; Hernandez, N.; Abdelli, F.; Martinez, J.M. Effect of different vegetation type cover on the soil water balance in semi-arid areas of south eastern Spain. Phys. Chem. Earth (B) 1999, 24, 353–357. [Google Scholar] [CrossRef]
- Bellot, J.; Bonet, A.; Sánchez, J.R.; Chirino, E. Likely effects of land use changes on the runoff aquifer recharge in a semiarid landscape using a hydrological model. Landsc. Urban Plan. 2001, 55, 41–53. [Google Scholar] [CrossRef]
- Chirino, E. Influencia De Las Precipitaciones Y De La Vegetación En El Balance Hídrico Superficial Y La Recarga De Acuíferos En Clima Semiárido. Ph.D. Thesis, Universidad de Alicante, Alicante, Spain, 2003. [Google Scholar]
- Llorens, P.; Latron, J.; Gallart, F. Dinámica Espacio-Temporal de la Humedad del Suelo en un Área de Montaña Mediterránea. Cuencas Experimentales de Vallcebre (Alto Llobregat). Available online: https://www.researchgate.net/profile/Pilar_Llorens/publication/268401867_DINAMICA_ESPACIO-TEMPORAL_DE_LA_HUMEDAD_DEL_SUELO_EN_UN_AREA_DE_MONTANA_MEDITERRANEA_CUENCAS_EXPERIMENTALES_DE_VALLCEBRE_ALTO_LLOBREGAT/links/55ed9e0d08ae21d099c77243/DINAMICA-ESPACIO-TEMPORAL-DE-LA-HUMEDAD-DEL-SUELO-EN-UN-AREA-DE-MONTANA-MEDITERRANEA-CUENCAS-EXPERIMENTALES-DE-VALLCEBRE-ALTO-LLOBREGAT.pdf (accessed on 1 June 2018).
- Nisbet, T. Water Use by Tree. Available online: https://www.forestry.gov.uk/pdf/FCIN065.pdf/$FILE/FCIN065.pdf (accessed on 1 June 2018).
- Moutahir, H.; Bellot, P.; Monjo, R.; Bellot, J.; Garcia, M.; Touhami, I. Likely effects of climate change on groundwater availability in a Mediterranean region of Southeastern Spain. Hydrol. Process. 2017, 31, 161–176. [Google Scholar] [CrossRef]
- Carrera-Hernandez, J.J.; Gaskin, S.J. Spatio-temporal analysis of potential aquifer recharge: Application to the basin of Mexico. J. Hydrol. 2008, 353, 228–246. [Google Scholar] [CrossRef]
- Touhami, I.; Andreu, J.M.; Chirino, E.; Sánchez, J.R.; Moutahir, H.; Pulido-Bosch, A.; Martínez-Santos, P.; Bellot, J. Recharge estimation of a small karstic aquifer in a semiarid Mediterranean region (Southeastern Spain) using a hydrological model. Hydrol. Process. 2013, 27, 165–174. [Google Scholar] [CrossRef]
- Andreu, J.M.; Delgado, J.; García-Sánchez, E.; Pulido-Bosch, A.; Bellot, J.; Chirino, E.; Ortíz de Urbina, J.M. Caracterización del funcionamiento y la recarga del acuífero del Ventós-Castellar (Alicante). Revista de la Sociedad Geológica de España 2001, 14, 247–254. [Google Scholar]
- Osem, Y.; O’Hara, K. An ecohydrological approach to managing dryland forests: Integration of leaf area metrics into assessment and management. Forestry 2016, 89, 338–349. [Google Scholar] [CrossRef]
- Del Campo, A.D.; Fernandes, T.J.G.; Molina, A.J. Hydrology-oriented (adaptive) silviculture in a semiarid pine plantation: How much can be modified the water cycle through forest management? Eur. J. For. Res. 2014, 133, 879–894. [Google Scholar] [CrossRef] [Green Version]
- Molina, A.J.; del Campo, A.D. The effects of experimental thinning on throughfalland stemflow: A contribution towards hydrology-oriented silviculture in Aleppopine plantations. For. Ecol. Manag. 2012, 269, 206–213. [Google Scholar] [CrossRef]
- Bellot, J.; Maestre, F.T.; Chirino, E.; Hernández, N.; de Urbina, J.O. Afforestation with Pinus halepensis reduces native shrub performance in a Mediterranean semiarid area. Acta Oecol. 2004, 25, 7–15. [Google Scholar] [CrossRef]
- Rodríguez-Robles, U.; Arredondo, J.T.; Huber-Sannwald, E.; Vargas, R. Geoecohydrological mechanisms couple soil and leaf water dynamics and facilitate species coexistence in shallow soils of a tropical semiarid mixed forest. New Phytol. 2015, 207, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Castillo, J.; Comas, C.; Voltas, J.; Ferrio, J.P. Dynamics of competition over water in a mixed oak-pine Mediterranean forest: Spatio-temporal and physiological components. For. Ecol. Manag. 2016, 382, 214–224. [Google Scholar] [CrossRef]
- David, T.S.; Henriques, M.O.; Kurz-Besson, C.; Nunes, J.; Valente, F.; Vaz, M.; Pereira, J.S.; Siegwolf, R.; Chaves, M.M.; Gazarini, L.C.; et al. Water-use strategies in two co-occurring Mediterranean evergreen oaks: Surviving the summer drought. Tree Physiol. 2007, 27, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Voltas, J.; Lucabaugh, D.; Chambel, M.R.; Ferrio, J.P. Intraspecific variation in the use of water sources by the circum-Mediterranean conifer Pinus halepensis. New Phytol. 2015, 208, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, E.K.S. Interplay between nutrients, water, root growth and productivity in young plantations. For. Ecol. Manag. 1990, 30, 213–232. [Google Scholar] [CrossRef]
- Querejeta, J.I.; Roldán, A.; Albaladejo, J.; Castillo, V. Soil water availability improved by site preparation in a Pinus halepensis afforestation under semiarid climate. For. Ecol. Manag. 2001, 149, 115–128. [Google Scholar] [CrossRef]
- Klein, T.; Rotenberg, E.; Cohen-Hilaleh, E.; Raz-Yaseef, N.; Tatarinov, F.; Preisler, Y.; Ogée, G.; Cohen, S.; Yakir, D. Quantifying transpirable soil water and its relations to tree water use dynamics in a water-limited pine forest. Ecohydrology 2014, 7, 409–419. [Google Scholar] [CrossRef]
- Moreno, J.M. A Preliminary Assessment of the Impacts in Spain Due to the Effects of Climate Change; ECCE Project-Final Report; Spanish Ministry of Environment-University of Castilla de la Mancha: Madrid, Spain, 2005; p. 786. [Google Scholar]
- Gonzalez-Hidalgo, J.C.; Peña-Angulo, D.; Simolo, C.; Brunetti, M.; Cortesi, N. Variación Espacial de las Tendencias de los Promedios Estacionales de las Máximas y de las Mínimas en España (1951–2010); Asociación Española de Climatología: Almería, Spain, 2014; pp. 99–108. [Google Scholar]
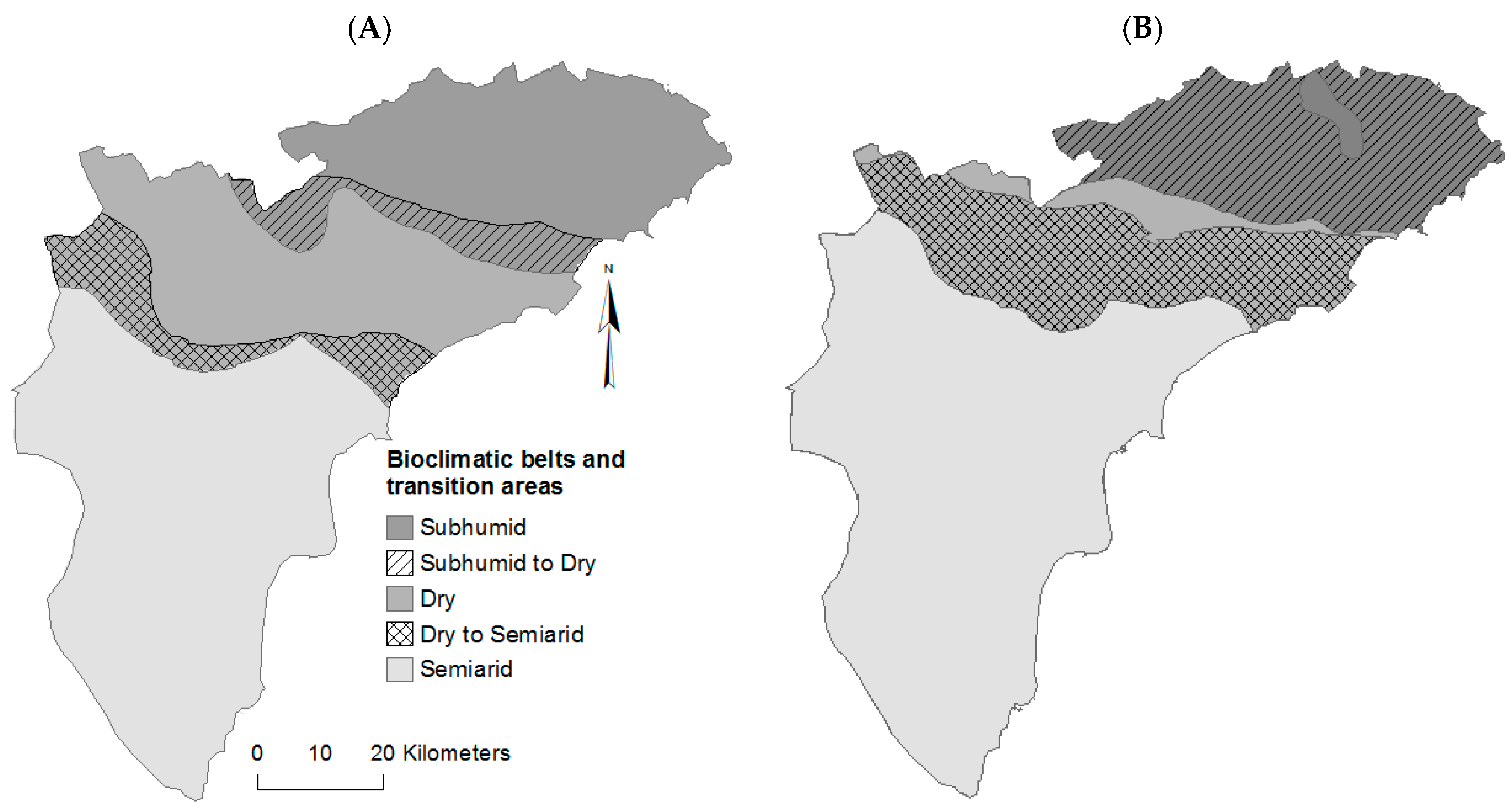
- Moutahir, H. Likely Effects of Climate Change on Water Resources and Vegetation Growth Period in the Province of Alicante, Southeastern Spain. Ph.D. Thesis, University of Alicante, Alicante, Spain, 2016. [Google Scholar]
- Rivas-Martínez, S. Pisos bioclimáticos de España. Lazaroa 1983, 5, 33–43. [Google Scholar]
- Rubel, F.; Kottek, M. Observed and projected climate shifs 1901–2100 depicted by world maps of Köppen-Geiger climate classification. Meteorol. Z. 2010, 19, 135–141. [Google Scholar] [CrossRef]
- Tweed, S.; Leblanc, M.; Cartwright, I.; Favreau, G.; Leduc, C. Arid zone groundwater recharge and salinisation processes; an example from the Lake Eyre Basin, Australia. J. Hydrol. 2011, 408, 257–275. [Google Scholar] [CrossRef]
- Manrique-Alba, À.; Ruiz-Yanetti, S.; Moutahir, H.; Novak, K.; De Luis, M.; Bellot, J. Soil moisture and its role in growth-climate relationships across an aridity gradient in semiarid Pinus halepensis forests. Sci. Total Environ. 2017, 574, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Moutahir, H.; Casady, G.; Manrique-Alba, A.; Ruiz-Yanetti, S.; Maturano, A.; Zeramdini, A.; Bellot Abad, J. Can we better understand land surface phenology changes using hydrological variables instead of climate variables? In Proceedings of the XIV MEDECOS & XIII AEET Meeting, Seville, Spain, 31 January–4 February 2017. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. halepensis | Q. ilex | ||
|---|---|---|---|
| Plant functional traits | |||
| Life form | Tree | Tree | |
| Leaf habits | Evergreen | Evergreen | |
| Life histories | Seeder | Resprouter | |
| Plant height | 15–20 m | 8–25 m | |
| Functional strategy to cope drought | Isohydric species Drought avoidance | Anisohydric species Drought-tolerant | |
| Cel-water relationships | |||
| Ψtlp | −MPa | 2.2 | 3.9 |
| Пo | −MPa | 1.7 | 2.8 |
| Emax | −MPa | 13 | 13 |
| WSDtlp | % | 17 | 30 |
| Gsmax | mmol m−2 s−1 | 270 | 250 |
| Stomata closure at midday | Strong | Moderate | |
| Photochemical efficiency PSII | Low photoinhibition | High photoinhibition | |
| SLW | mg cm−2 | 18.5 | 21.0 |
| Lethal leaf temperature | °C | 50 | 45 |
| Xylem conducting system | |||
| Dmax | μm | 13.6 | 16.7 |
| PLC50 | −4.7 | −5.5 | |
| LSC | m2 MPa−1 s−1 10−8 | 9.54 | 5.96 |
| Ks | m2 MPa−1 s−1 10−4 | 1.52 | 1.04 |
| Root system | |||
| Rooting depth | Medium/Deep | Deep | |
| R/S | Low | High | |
| khroot | kg s−1 m2 MPa−1 10−4 | 0.42 | 2.49 |
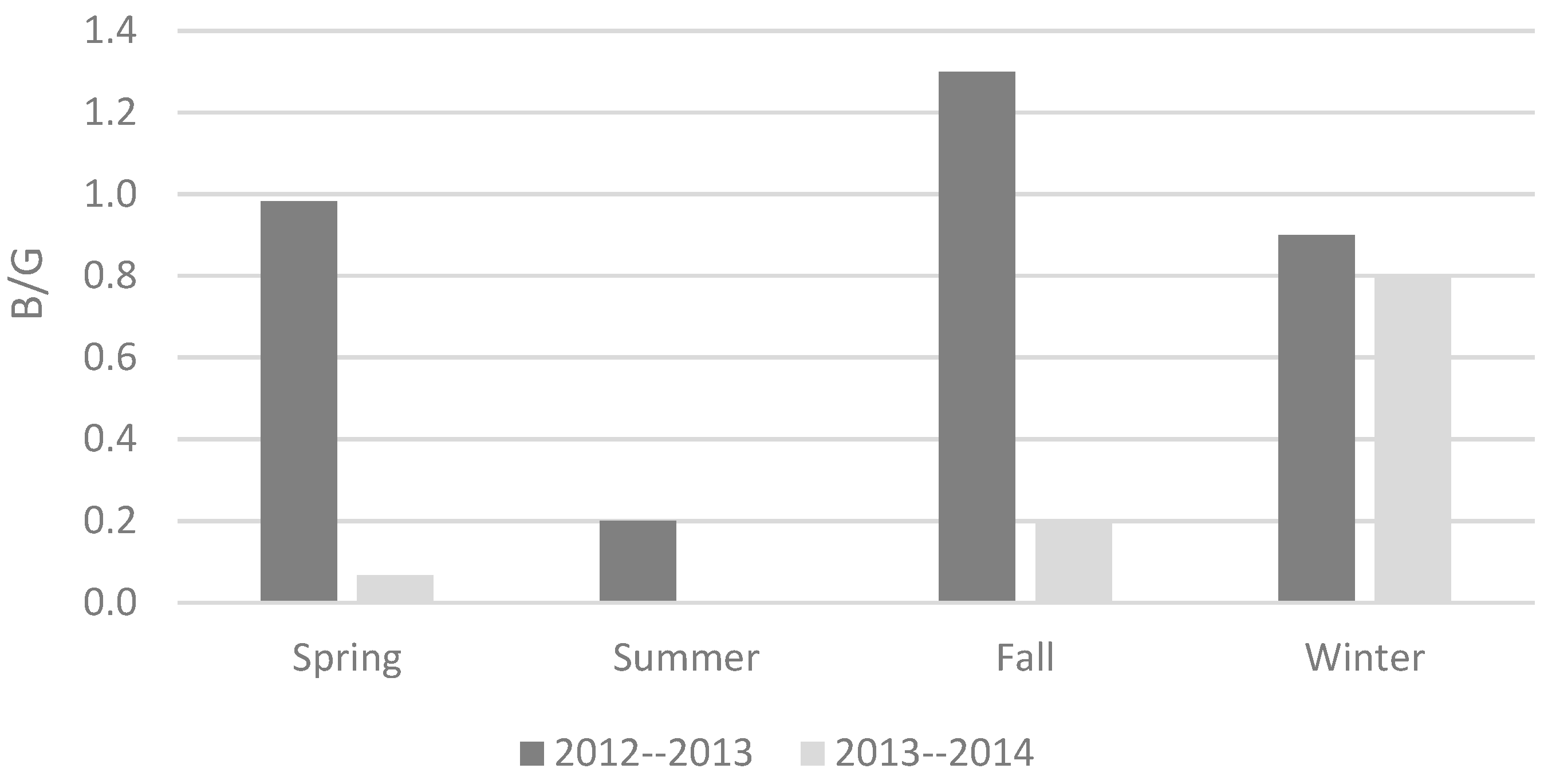
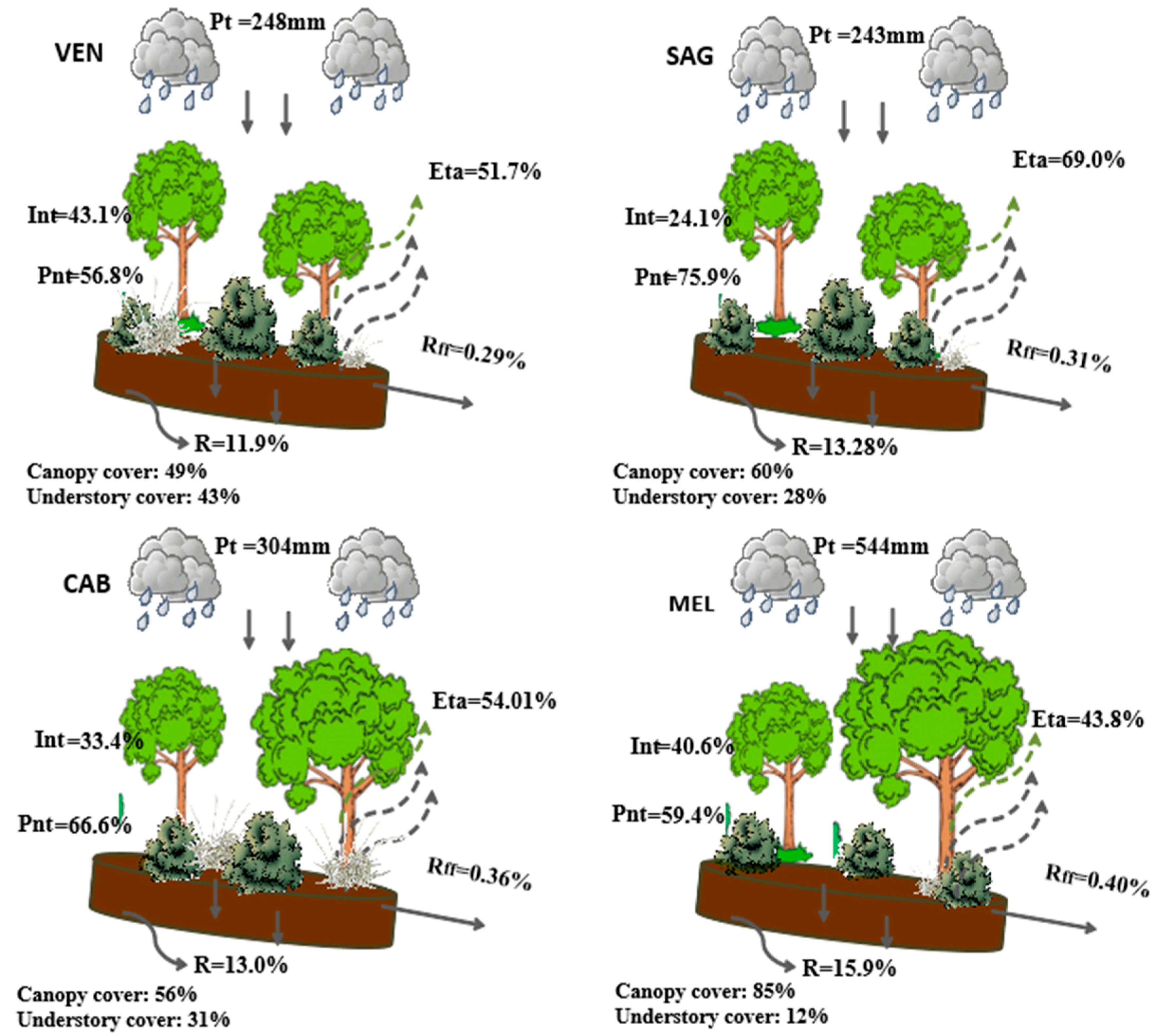
| Years | Species | Site Name | P (mm) | ET + Int (%) | Roff (%) | R (%) | B/G |
|---|---|---|---|---|---|---|---|
| 2012–2013 | P. halepensis | MEL | 911 | 72 | 0.5 | 27.6 | 0.39 |
| CAB | 417 | 82.1 | 0.4 | 17.6 | 0.22 | ||
| VEN | 425 | 79.9 | 0.3 | 19.8 | 0.25 | ||
| SAG | 420 | 79.4 | 0.3 | 20.3 | 0.26 | ||
| Q. ilex | 534 | 51 | 0 | 49 | 0.9 | ||
| 2013–2014 | P. halepensis | MEL | 318 | 91.1 | 0.4 | 8.5 | 0.1 |
| CAB | 146 | 95.1 | 0.4 | 4.6 | 0.05 | ||
| VEN | 114 | 97.8 | 0.2 | 2 | 0.02 | ||
| SAG | 142 | 91.4 | 0.3 | 8.3 | 0.09 | ||
| Q. ilex | 271 | 81 | 0 | 19 | 0.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicente, E.; Vilagrosa, A.; Ruiz-Yanetti, S.; Manrique-Alba, À.; González-Sanchís, M.; Moutahir, H.; Chirino, E.; Del Campo, A.; Bellot, J. Water Balance of Mediterranean Quercus ilex L. and Pinus halepensis Mill. Forests in Semiarid Climates: A Review in A Climate Change Context. Forests 2018, 9, 426. https://doi.org/10.3390/f9070426
Vicente E, Vilagrosa A, Ruiz-Yanetti S, Manrique-Alba À, González-Sanchís M, Moutahir H, Chirino E, Del Campo A, Bellot J. Water Balance of Mediterranean Quercus ilex L. and Pinus halepensis Mill. Forests in Semiarid Climates: A Review in A Climate Change Context. Forests. 2018; 9(7):426. https://doi.org/10.3390/f9070426
Chicago/Turabian StyleVicente, Eduardo, Alberto Vilagrosa, Samantha Ruiz-Yanetti, Àngela Manrique-Alba, María González-Sanchís, Hassane Moutahir, Esteban Chirino, Antonio Del Campo, and Juan Bellot. 2018. "Water Balance of Mediterranean Quercus ilex L. and Pinus halepensis Mill. Forests in Semiarid Climates: A Review in A Climate Change Context" Forests 9, no. 7: 426. https://doi.org/10.3390/f9070426
APA StyleVicente, E., Vilagrosa, A., Ruiz-Yanetti, S., Manrique-Alba, À., González-Sanchís, M., Moutahir, H., Chirino, E., Del Campo, A., & Bellot, J. (2018). Water Balance of Mediterranean Quercus ilex L. and Pinus halepensis Mill. Forests in Semiarid Climates: A Review in A Climate Change Context. Forests, 9(7), 426. https://doi.org/10.3390/f9070426