Growth of Pinus cembroides Zucc. in Response to Hydroclimatic Variability in Four Sites Forming the Species Latitudinal and Longitudinal Distribution Limits


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
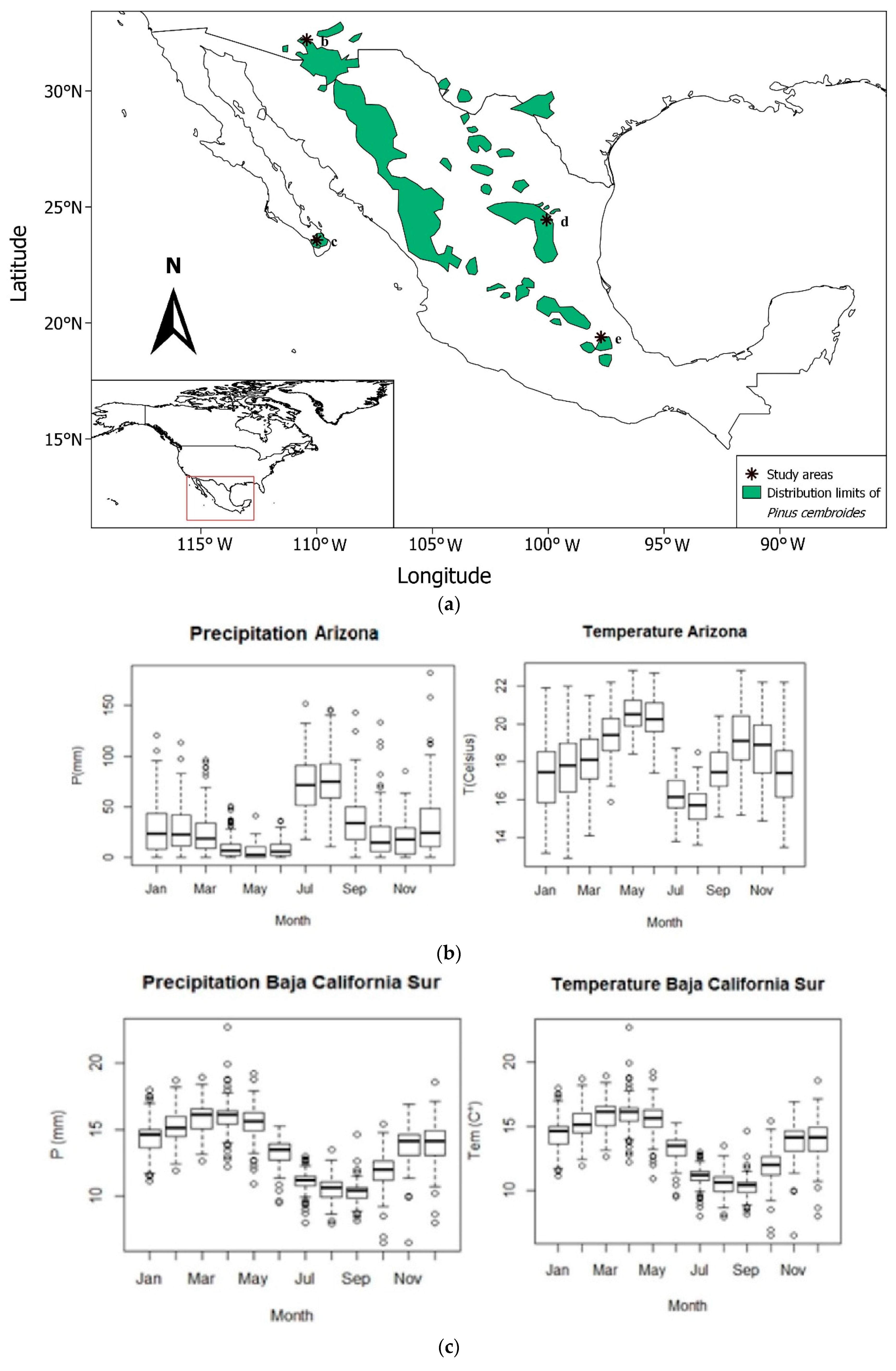
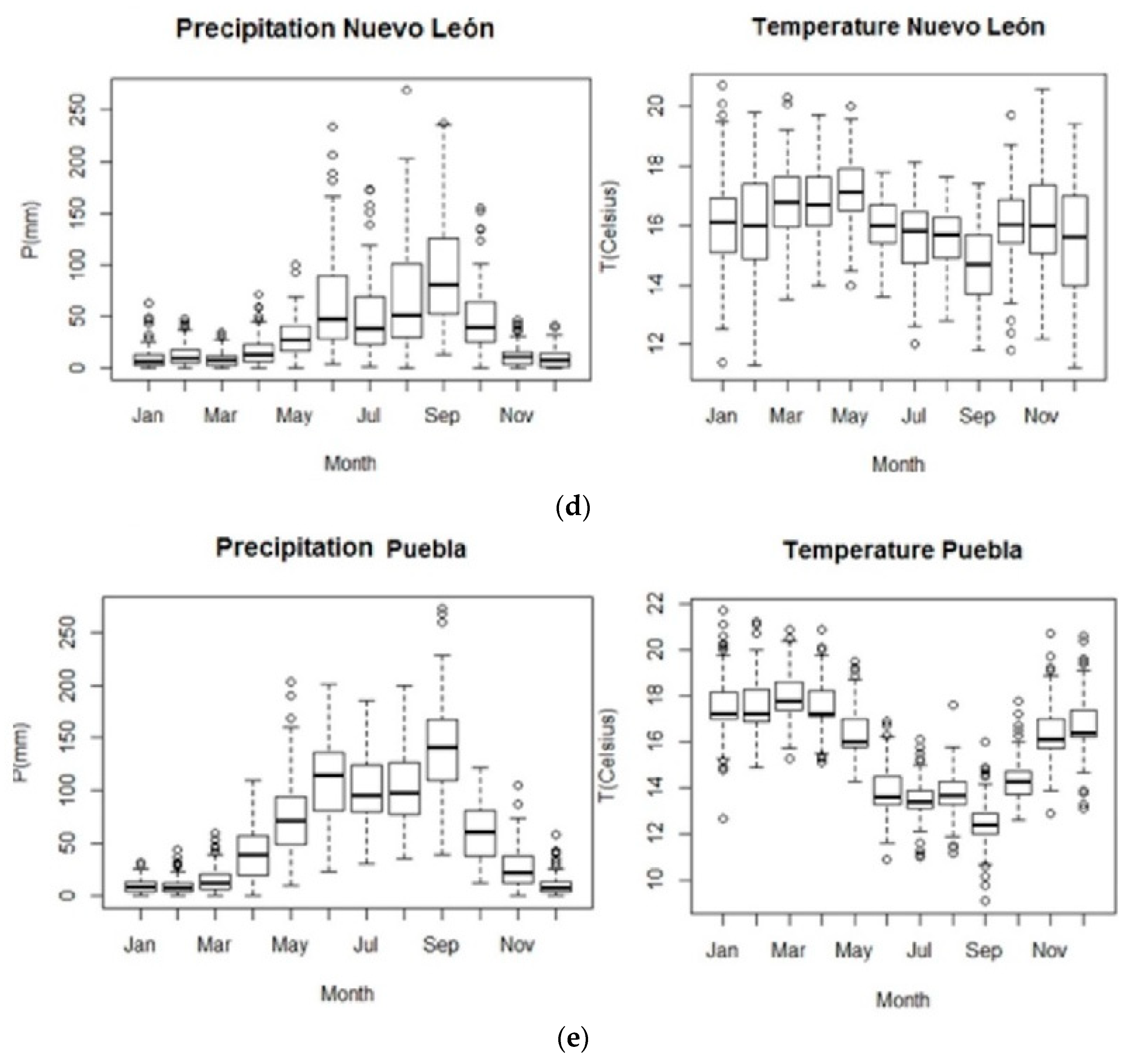
2.2. Study Sites and Dendrochronological Data
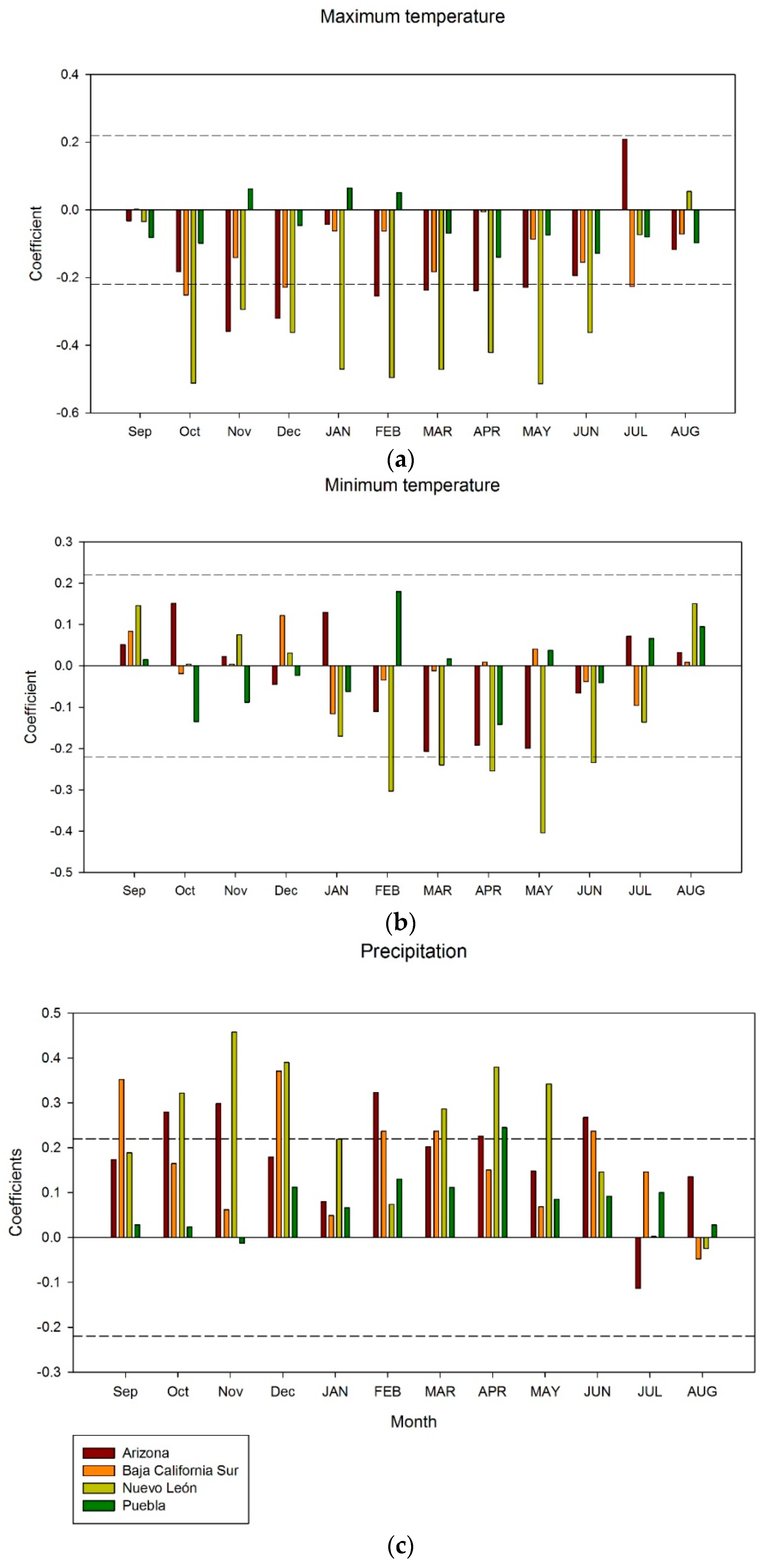
2.3. Relationships between Climate, Drought and Growth
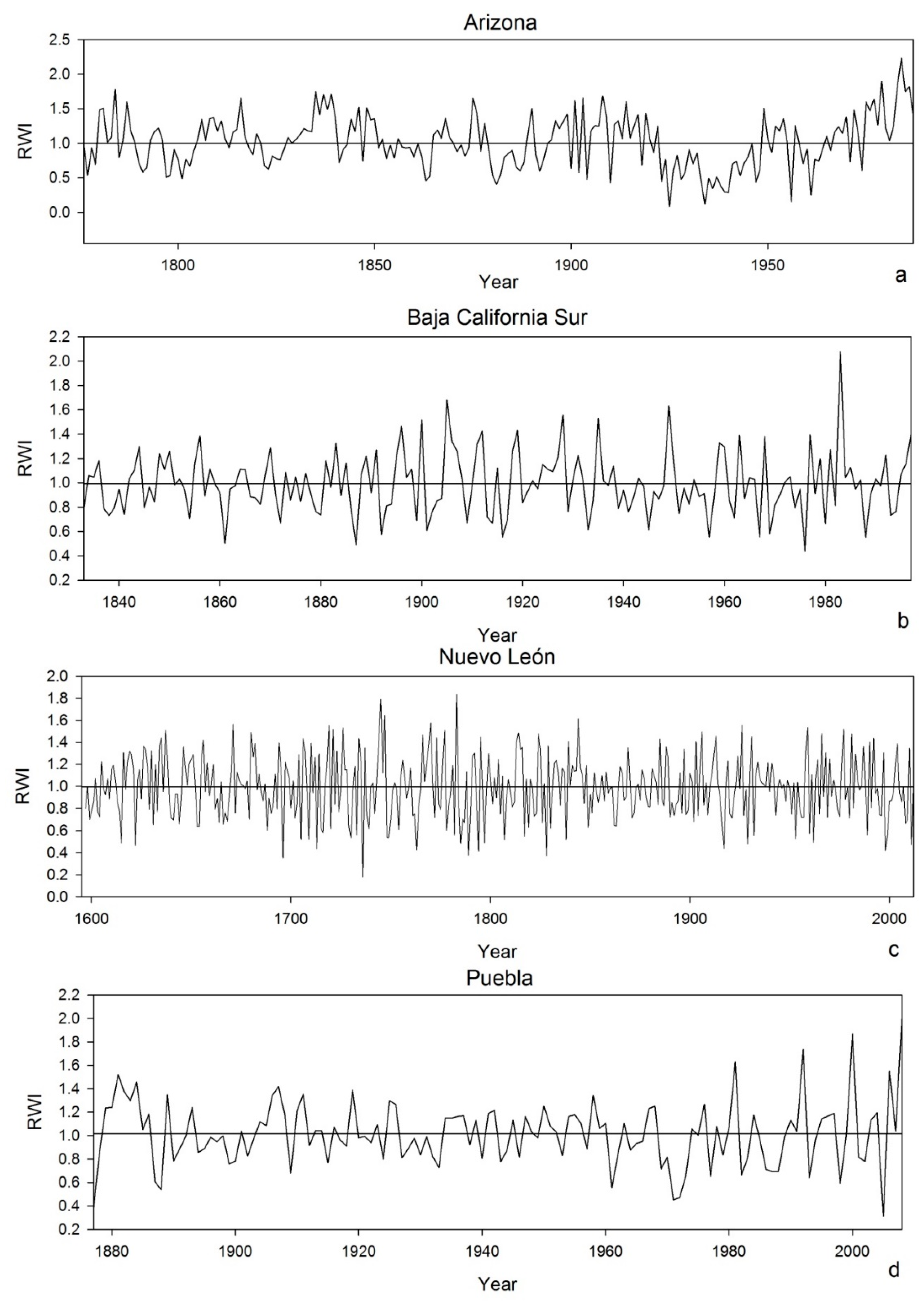
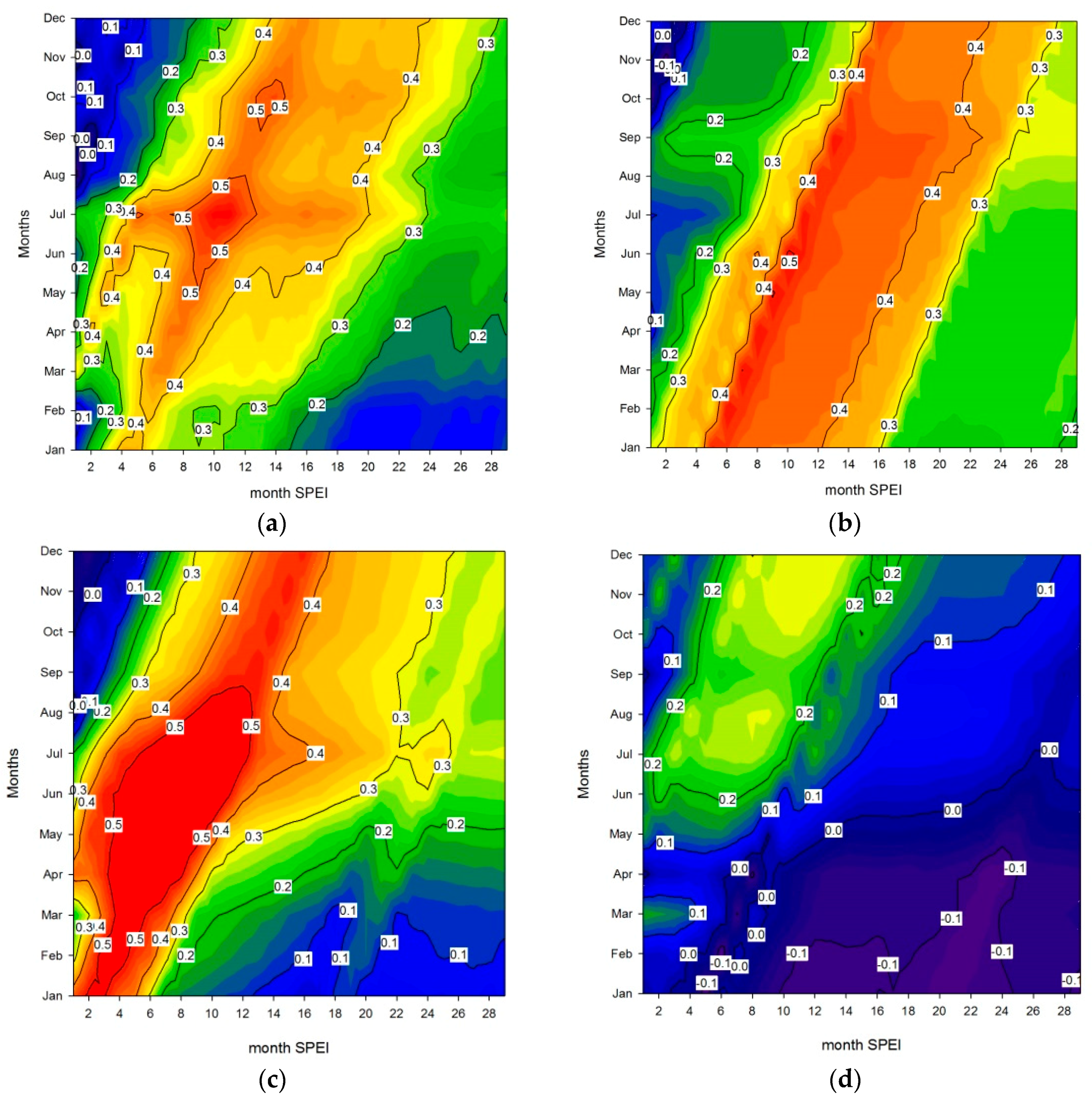
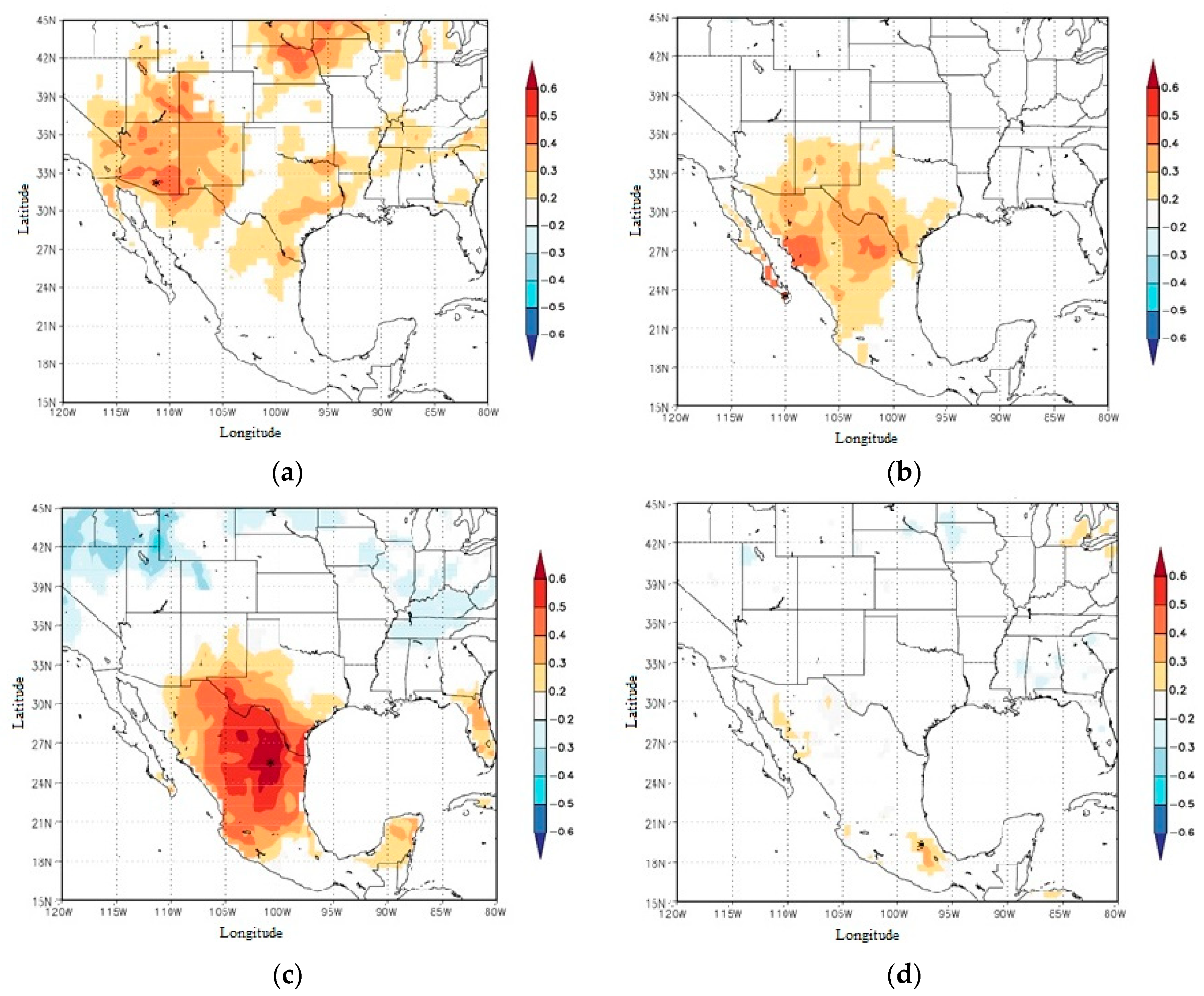
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schwab, N.; Kaczka, R.J.; Janecka, K.; Böhner, J.; Chaudhary, R.P.; Scholten, T.; Schickhoff, U. Climate Change-Induced Shift of Tree Growth Sensitivity at a Central Himalayan Treeline Ecotone. Forests 2018, 9, 267. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fulé, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Redmond, M.; Forcella, F.; Barger, N. Declines in pinyon pine cone production associated with regional warming. Ecosphere 2012, 3, 1–14. [Google Scholar] [CrossRef]
- Ortiz, E.; Villaseñor, J.L. Modelación de la distribución potencial del bosque húmedo de montaña en México. Agroproductividad 2017, 10, 24–28. [Google Scholar]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jump, A.; Hunt, J.M.; Penuelas, J. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob. Chang. Biol. 2006, 12, 2163–2174. [Google Scholar] [CrossRef]
- Caminero, L.; Génova, M.; Camarero, J.J.; Sánchez-Salguero, R. Growth responses to climate and drought at the southernmost European limit of Mediterranean Pinus pinaster forests. Dendrochronologia 2018, 48, 20–29. [Google Scholar] [CrossRef]
- González-Cásares, M.; Pompa-García, M.; Camarero, J.J. Differences in climate–growth relationship indicate diverse drought tolerances among five pine species coexisting in Northwestern Mexico. Trees 2016, 31, 531–544. [Google Scholar] [CrossRef]
- Gazol, A.; Ribas, M.; Gutiérrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought-induced growth decline and partial recovery. Agric. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- Esperón-Rodríguez, M.; Barradas, V. Potential vulnerability to climate change of four tree species from the central mountain region of Veracruz, Mexico. Clim. Res. 2014, 60, 163–174. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Y.; Zhao, S.; Jiao, L.; Wen, Y. Relationships between Tree Age and Climate Sensitivity of Radial Growth in Different Drought Conditions of Qilian Mountains, Northwestern China. Forests 2018, 9, 135. [Google Scholar] [CrossRef]
- Camarero, J.J.; Manzanedo, R.D.; Sánchez-Salguero, R.; Navarro-Cerrillo, R. Growth response to climate and drought change along an aridity gradient in the southernmost Pinus nigra relict forests. Ann. For. Sci. 2013, 70, 769–780. [Google Scholar] [CrossRef]
- Constante, G.; Villanueva, D.; Cerano, P.; Cornejo, O.; Valencia, M. Dendrocronología de P. cembroides Zucc. y reconstrucción de precipitación estacional para el suroeste de Coahuila. Cienc. forestal en México 2009, 34, 17–40. [Google Scholar]
- Ortiz-Medrano, A.; Scantlebury, D.; Vázquez-Lobo, A.; Mastretta-Yanes, A.; Piñero, D. Morphological and niche divergence of pinyon pines. Ecol. Evol. 2016, 6, 2886–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompa-García, M.; González-Cásares, M.; Acosta-Hernández, A.; Camarero, J.; Rodríguez-Catón, M. Drought influence over radial growth of Mexican conifers inhabiting mesic and xeric sites. Forests 2017, 8, 175. [Google Scholar] [CrossRef]
- Irby, C.M.; Fulé, P.Z.; Yocom, L.L.; Villanueva, D. Dendrochronological reconstruction of long-term precipitation patterns in Basaseachi National Park, Chihuahua, Mexico. Madera y Bosques 2013, 19, 93–105. [Google Scholar]
- Villanueva, J.; Cerano, J.; Fulé, P.; Cortés, C.; Vázquez, L.; Yocom, L.; Ruiz, J. Cuatro siglos de variabilidad hidroclimática en el noroeste de Chihuahua, México, reconstruida con anillos de árboles. Investig. Geográficas 2015, 87, 141–153. [Google Scholar] [CrossRef]
- Ruiz-Talonia, L.; Sánchez-Vargas, N.; Bayuelo-Jiménez, J.; Lara-Cabrera, S.; Sáenz-Romero, C. Altitudinal genetic variation among native Pinus patula provenances: Performance in two locations, seed zone delineation and adaptation to climate change. Silvae Genetica 2014, 63, 139–149. [Google Scholar] [CrossRef]
- De León, M. Factores que Influyen en la Producción de Piñón de P. cembroides Zucc. Bachelor’s Thesis, Universidad Autónoma Agraria Antonio Narro, Coahuila, México, 2017. [Google Scholar]
- Gutiérrez-García, J.; Rodríguez-Trejo, D.; Villanueva-Morales, A.; García-Díaz, S.; Romo-Lozano, J. Calidad del agua en la producción de Pinus cembroides Zucc. en vivero. Agrociencia 2015, 49, 205–219. [Google Scholar]
- Farjon, A.; Pérez, J.; Styles, B. Field Guide to the Pines of Mexico and Central America; Royal Botanic Gardens: Kew, UK, 1997; 147p. [Google Scholar]
- International Tree Ring Data Bank. Tree Ring|National Centers for Environmental Information (NCEI) formerly known as National Climatic Data Center (NCDC). Available online: https://www.ncdc.noaa.gov/data-access/paleoclimatology-data/datasets/tree-ring (accessed on 17 April 2018).
- Harris, I.C.; Jones, P.D. CRU TS4. 00: Climatic Research Unit (CRU) Time-Series (TS) Version 4.00 of High Resolution Gridded Data of Month-by-Month Variation in Climate (Jan. 1901–Dec. 2015). Centre for Environmental Data Analysis: Oxford, UK, 2017. Available online: http://dx.doi.org/10.5285/edf8febfdaad48abb2cbaf7d7e846a86 (accessed on 17 April 2018).
- Bunn, A. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences. International Institute for Applied Systems Analysis, Kluwer Academic Publishers: Boston, MA, USA, 1990; 288p. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J. A multi-scalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Index: SPEI, The Standardised Precipitation-Evapotranspiration Index. Available online: http://spei.csic.es (accessed on 17 April 2018).
- CRAN-Package SPEI. Calculation of the Standardised precipitation-Evotranspiration Index. Available online: http://CRAN.R-project.org/package=SPEI (accessed on 10 February 2017).
- Bickford, I.; Fulé, P.; Kolb, T. Growth sensitivity to drought of co-occurring Pinus spp. along an elevation gradient in northern Mexico. West. N. Am. Nat. 2011, 71, 338–348. [Google Scholar] [CrossRef]
- De Soto, L.; Camarero, J.J.; Olano, J.M.; Rozas, V. Geographically structured and temporally unstable growth responses of Juniperus thurifera to recent climate variability in the Iberian Peninsula. Eur. J. For. Res. 2012, 131, 905–917. [Google Scholar] [CrossRef] [Green Version]
- Pompa-García, M.; Cerano-Paredes, J.; Fulé, P. Variation in radial growth of Pinus cooperi in response to climatic signals across an elevational gradient. Dendrochronologia 2013, 31, 198–204. [Google Scholar] [CrossRef]
- Castruita-Esparza, L.; Correa-Díaz, A.; Gómez-Guerrero, A.; Villanueva-Díaz, J.; Ramírez-Guzmán, M.; Velázquez-Martínez, A.; Ángeles-Pérez, G. Basal area increment series of dominant trees of Pseudotsuga menziesii (Mirb.) Franco show periodicity according to global climate patterns. Revista Chapingo. Serie Cienc. For. y del Ambiente 2016, 22, 379–397. [Google Scholar] [CrossRef]
- Bertini, G.; Amoriello, T.; Fabbio, G.; Piovosi, M. Forest growth and climate change: Evidences from the ICP-Forests intensive monitoring in Italy. iForest 2011, 4, 262–267. [Google Scholar] [CrossRef]
- Gómez-Guerrero, A.; Silva, L.; Barrera-Reyes, M.; Kishchuk, B.; Velázquez-Martínez, A.; Martínez-Trinidad, T.; Plascencia-Escalante, F.; Horwath, W. Growth decline and divergent tree ring isotopic composition (δ13C and δ18O) contradict predictions of CO2 stimulation in high altitudinal forests. Glob. Chang. Biol. 2013, 19, 1748–1758. [Google Scholar]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Mérian, P.; Courdier, F.; Ladier, J.; Dreyfus, P. Instability of climate signal in tree-ring width in Mediterranean mountains: A multi-species analysis. Trees 2012, 26, 715–729. [Google Scholar] [CrossRef]
- Huang, J.; Tardif, J.; Bergeron, Y.; Denneler, B.; Berninger, F.; Girardin, M.P. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Glob. Chang. Biol. 2010, 16, 711–731. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 18, 471–480. [Google Scholar] [CrossRef] [PubMed]
- González-Elizondo, M.; Jurado, E.; Návar, J.; González-Elizondo, M.S.; Villanueva, J.; Aguirre, O.; Jiménez, J. Tree-rings and climate relationships for Douglas-fir chronologies from the Sierra Madre Occidental, Mexico: A 1681–2001 rain reconstruction. For. Ecol. Manag. 2005, 213, 39–53. [Google Scholar] [CrossRef]
- Pompa-García, M.; Camarero, J.J. Reconstructing evaporation from pine tree rings in northern Mexico. Tree-Ring Res. 2015, 71, 95–105. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; Vicente-Serrano, S. Climatic impacts and drought control of radial growth and seasonal wood formation in Pinus halepensis. Trees 2012, 26, 1875–1886. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorín-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yocom, L.; Fulé, P.Z.; Brown, P.M.; Cerano, J.; Villanueva-Díaz, J.; Falk, D.A.; Cornejo-Oviedo, E. El Niño-Southern Oscillation effect on a fire regime in northeastern Mexico has changed over time. Ecology 2010, 91, 1660–1671. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study site | Latitude (N) | Longitude (W) | Elevation (m a. s. l.) | ITRDB Code | Contributors |
|---|---|---|---|---|---|
| Arizona | 32.21 | 110.43 | 1645 | az506 | J.A. Parks |
| California Sur | 23.58 | 109.99 | 2300 | mexi105 | J. Villanueva |
| Nuevo León | 24.73 | 100.12 | 1265 | mexi093 | J. Villanueva |
| Puebla | 19.39 | 97.72 | 2582 | mexi102 | J. Villanueva |
| Study Site | Timespan | No. Series | Mean ± SD Tree-Ring Width (mm) | AC | MS | Series Intercorrelation | Rbt | SNR | EPS |
|---|---|---|---|---|---|---|---|---|---|
| Arizona | 1766–1987 | *NA | *NA | *NA | *NA | *NA | *NA | *NA | *NA |
| Baja California Sur | 1833–1997 | 17 | 2.03 ± 0.88 | 0.38 | 0.36 | 0.68 | 0.45 | 7.39 | 0.88 |
| Nuevo León | 1597–2012 | 57 | 1.47 ± 0.82 | 0.56 | 0.43 | 0.69 | 0.32 | 36.2 | 0.97 |
| Puebla | 1873–2008 | 51 | 1.80 ± 1.10 | 0.78 | 0.68 | 0.68 | 0.45 | 27.6 | 0.96 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera-Soto, G.; González-Cásares, M.; Pompa-García, M.; Camarero, J.J.; Solís-Moreno, R. Growth of Pinus cembroides Zucc. in Response to Hydroclimatic Variability in Four Sites Forming the Species Latitudinal and Longitudinal Distribution Limits. Forests 2018, 9, 440. https://doi.org/10.3390/f9070440
Herrera-Soto G, González-Cásares M, Pompa-García M, Camarero JJ, Solís-Moreno R. Growth of Pinus cembroides Zucc. in Response to Hydroclimatic Variability in Four Sites Forming the Species Latitudinal and Longitudinal Distribution Limits. Forests. 2018; 9(7):440. https://doi.org/10.3390/f9070440
Chicago/Turabian StyleHerrera-Soto, Gerardo, Marcos González-Cásares, Marín Pompa-García, J. Julio Camarero, and Raúl Solís-Moreno. 2018. "Growth of Pinus cembroides Zucc. in Response to Hydroclimatic Variability in Four Sites Forming the Species Latitudinal and Longitudinal Distribution Limits" Forests 9, no. 7: 440. https://doi.org/10.3390/f9070440
APA StyleHerrera-Soto, G., González-Cásares, M., Pompa-García, M., Camarero, J. J., & Solís-Moreno, R. (2018). Growth of Pinus cembroides Zucc. in Response to Hydroclimatic Variability in Four Sites Forming the Species Latitudinal and Longitudinal Distribution Limits. Forests, 9(7), 440. https://doi.org/10.3390/f9070440