What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments?

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
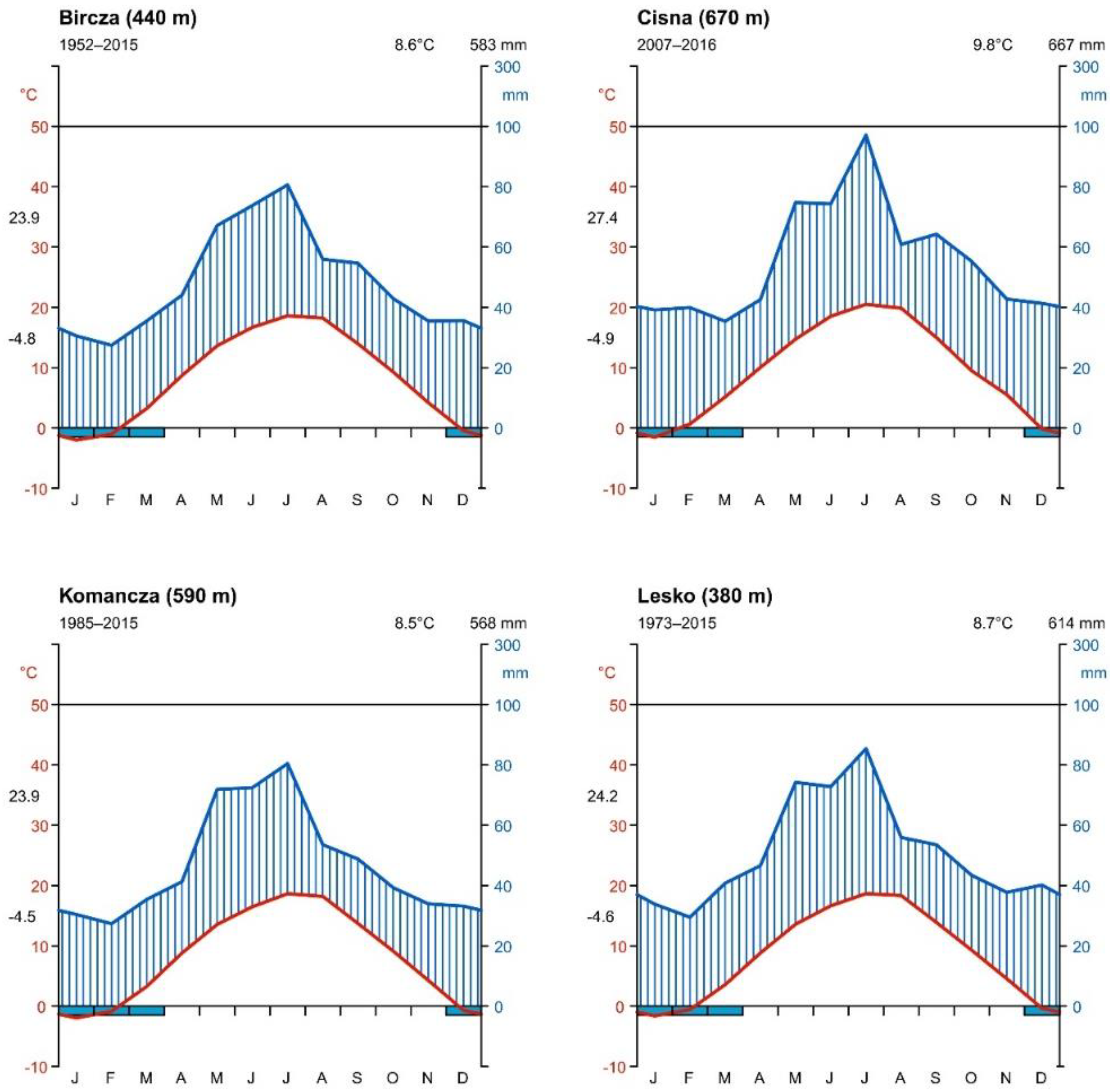
3.1. Bioclimatic Analyzes
3.2. Analysis of Variance
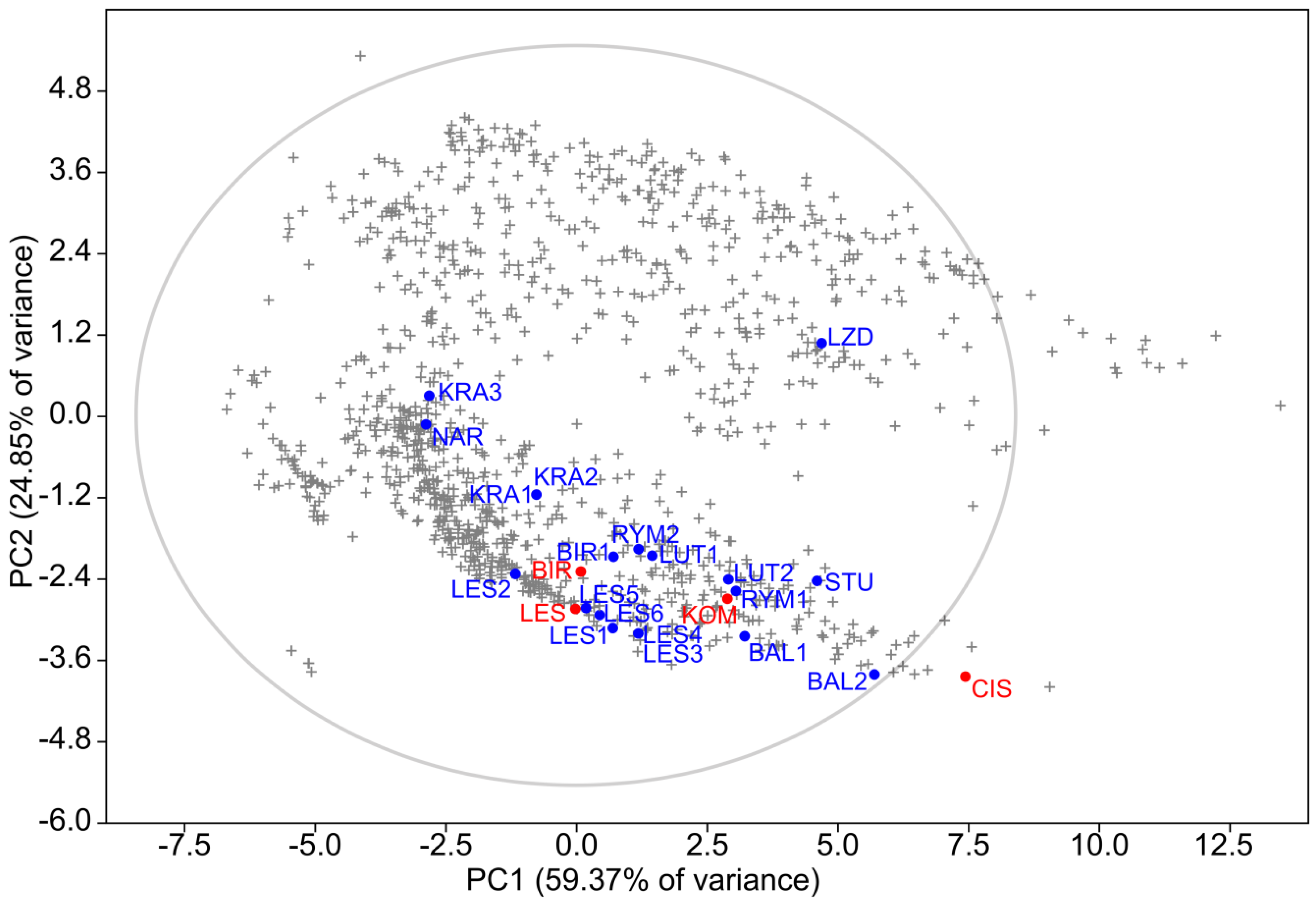
3.3. Similarity of Environments
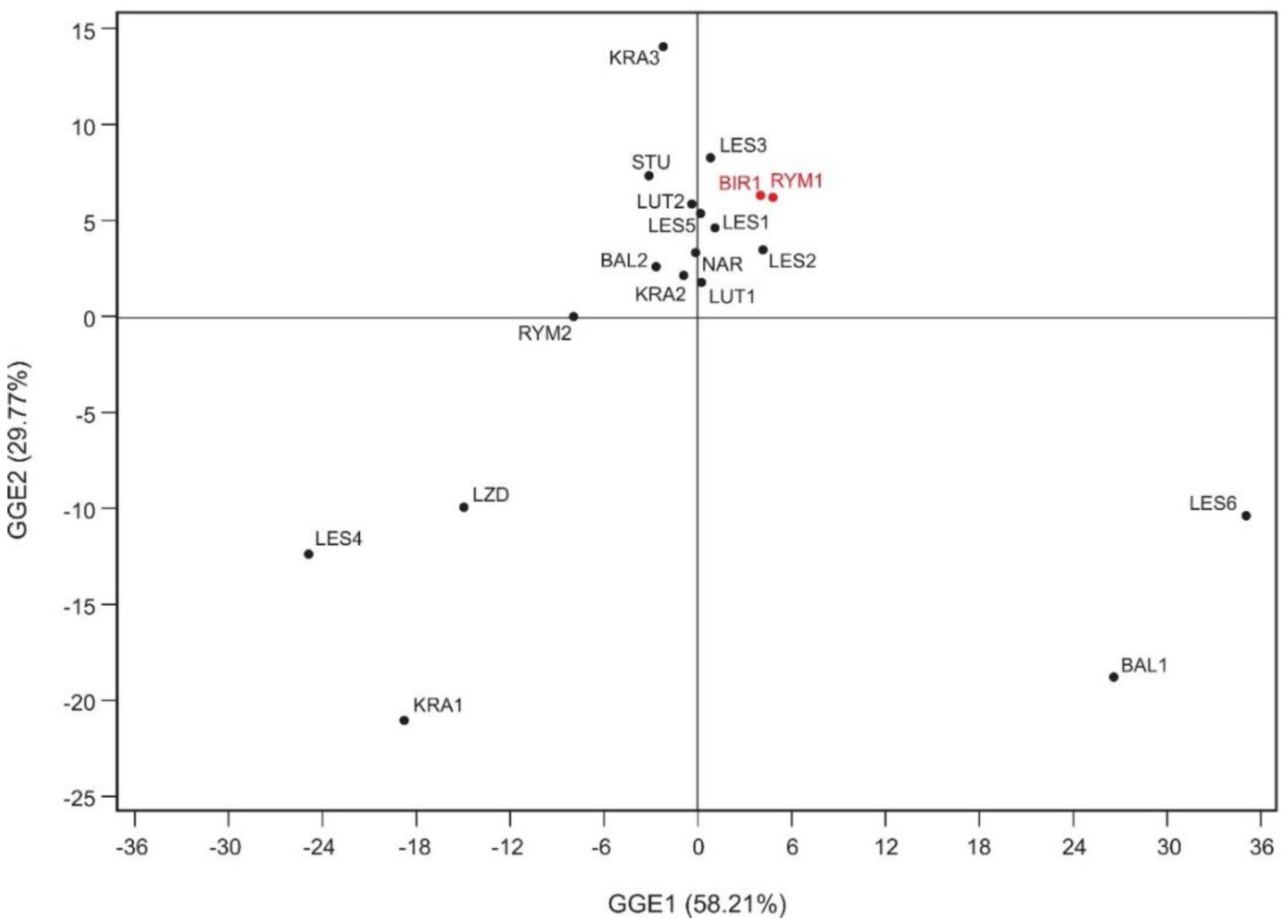
3.4. Similarity and Stability of Populations and Ideal Genotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AE | Average environment |
| AEC | Average environment coordination |
| BIR | Bircza experimental site |
| CIS | Cisna experimental site |
| E | Effect of environment |
| G | Effect of genotype |
| G × E | Genotype-environment interaction |
| GGE | Genotype and genotype-by-environment interaction biplot |
| KOM | Komańcza experimental site |
| LES | Lesko experimental site |
| MET | Multi-environment trial |
| PC1 | First principal component |
| PC2 | Second principal component |
| PCA | Principal components analysis |
| SVD | singular value decomposition |
References
- Soule, M. The Epistasis Cycle: A Theory of Marginal Populations. Annu. Rev. Ecol. Syst. 1973, 4, 165–187. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct effects of climate warming on populations of silver fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Skov, F. Limited filling of the potential range in European tree species: Limited range filling in European trees. Ecol. Lett. 2004, 7, 565–573. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.; Carraro, G.; Conedera, M. The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Koprowski, M. Reaction of Silver Fir (Abies alba) Growing Outside its Natural Range to Extreme Weather Events and a Long-Term Increase in March temperature. Tree-Ring Res. 2013, 69, 49–61. [Google Scholar] [CrossRef]
- Litkowiec, M.; Lewandowski, A.; Raczka, G. Spatial pattern of the mitochondrial and chloroplast genetic variation in Poland as a result of the migration of Abies alba Mill. from different glacial refugia. Forests 2016, 7, 284. [Google Scholar] [CrossRef]
- Eckert, C.G.; Samis, K.E.; Lougheed, S.C. Genetic variation across species’ geographical ranges: The central-marginal hypothesis and beyond. Mol. Ecol. 2008, 17, 1170–1188. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, A.; Filipiak, M.; Burczyk, J. Genetic variation of Abies alba Mill. In Polish part of Sudety Mts. Acta Soc. Bot. Pol. 2001, 70, 215–219. [Google Scholar] [CrossRef]
- Mejnartowicz, L. Genetic analysis of silver-fir populations in the North Carpathian and Sudeten Mountains (Poland). Acta Soc. Bot. Pol. 2004, 73, 285–292. [Google Scholar] [CrossRef]
- Larsen, J. Waldbauliche Probleme und Genökologie der Weisstanne (Abies alba Mill.). Allg. Forst. Jagdztg. 1989, 160, 39–43. [Google Scholar]
- Shestakova, T.A.; Gutiérrez, E.; Kirdyanov, A.V.; Camarero, J.J.; Génova, M.; Knorre, A.A.; Linares, J.C.; de Dios, V.R.; Sánchez-Salguero, R.; Voltas, J. Forests synchronize their growth in contrasting Eurasian regions in response to climate warming. Proc. Natl. Acad. Sci. USA 2016, 113, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Matías, L.; Linares, J.C.; Sánchez-Miranda, Á.; Jump, A. Contrasting growth forecasts across the geographical range of Scots pine due to altitudinal and latitudinal differences in climatic sensitivity. Glob. Chang. Biol. 2017, 23, 4106–4116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant. Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgins, K.A.; Moore, J.L. Adapting to a warming world: Ecological restoration, climate change, and genomics. Am. J. Bot. 2016, 103, 590–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, A.; Sperisen, C.; Howe, G.T.; Brang, P.; Walthert, L.; St.Clair, J.B.; Heiri, C. Distinct genecological patterns in seedlings of Norway spruce and silver fir from a mountainous landscape. Ecology 2017, 98, 211–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, R.G.; Etterson, J.R. Rapid climate change and the rate of adaptation: Insight from experimental quantitative genetics. New Phytol. 2012, 195, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.D.; Cameron, A.; Green, R.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.Q.; Dong, Y.-F.; Herrando-Moraira, S.; Matsui, T.; Ohashi, H.; He, L.-Y.; Nakao, K.; Tanaka, N.; Tomita, M.; Li, X.-S.; et al. Potential effects of climate change on geographic distribution of the Tertiary relict tree species Davidia involucrata in China. Sci. Rep. 2017, 7, 43822. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-J.; Wan, J.-Z.; Zhang, Z.-X. Expansion potential of invasive tree plants in ecoregions under climate change scenarios: An assessment of 54 species at a global scale. Scand. J. For. Res. 2017, 32, 663–670. [Google Scholar] [CrossRef]
- Duan, R.-Y.; Kong, X.-Q.; Huang, M.-Y.; Fan, W.-Y.; Wang, Z.-G. The Predictive Performance and Stability of Six Species Distribution Models. PLoS ONE 2014, 9, e112764. [Google Scholar] [CrossRef] [PubMed]
- Matyas, C. Modeling climate change effects with provenance test data. Tree Physiol. 1994, 14, 797–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, J.P.; Schueler, S.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Grabner, M. Inter- and intra-specific variation in drought sensitivity in Abies sp. and its relation to wood density and growth traits. Agr. For. Meteorol. 2015, 214, 430–443. [Google Scholar] [CrossRef] [PubMed]
- George, J.P.; Grabner, M.; Karanitsch-Ackerl, S.; Mayer, K.; Weißenbacher, L.; Schueler, S. Genetic variation, phenotypic stability, and repeatability of drought response in European larch throughout 50 years in a common garden experiment. Tree Physiol. 2016, 37, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Pigliucci, M. Evolution of phenotypic plasticity: Where are we going now? Trends Ecol. Evol. 2005, 20, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Barzdajn, W. Adaptation of different silver fir (Abies alba Mill.) provenances to the conditions of the Sudetes. For. Res. Pap. 2009, 70, 49–58, (In Polish with English abstract). [Google Scholar] [CrossRef]
- Kempf, M.; Sabor, J. Evaluation of the variability of adaptive traits in 5−year−old silver fir progenies from provenances protected on conservation plots in the Carpathian Gene Bank. Sylwan 2009, 153, 651–661, (In Polish with English abstract). [Google Scholar]
- Skrzyszewska, K. Genetic and breeding value of Silver fir (Abies alba Mill.) represented in All-Poland Provenance Test Jd PL 86/90. Zesz. Nauk. Akad. Rol. Krakowie 1999, 61, 43–66, (In Polish with English abstract). [Google Scholar]
- Matías, L.; Gonzalez-Díaz, P.; Quero, J.L.; Camarero, J.J.; Lloret, F.; Jump, A.S. Role of geographical provenance in the response of silver fir seedlings to experimental warming and drought. Tree Physiol. 2016, 36, 1236–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, A.; Howe, G.T.; Sperisen, C.; Brang, P.; Schmatz, D.R.; Heiri, C. Risk of genetic maladaptation due to climate change in three major European tree species. Glob. Chang. Biol. 2017, 23, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sabor, J. Genetic value of Polish Carpathian fir. Zesz. Nauk. Akad. Rol. Krakowie 1999, 61, 29–41, (In Polish with English abstract). [Google Scholar]
- Klisz, M.; Jastrzębowski, S.; Ukalska, J.; Przybylski, P.; Matras, J.; Mionskowski, M. The vulnerability of silver fir populations to damage from late frosts. For. Res. Pap. 2016, 77, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Vessella, F.; Schirone, B. Predicting potential distribution of Quercus suber in Italy based on ecological niche models: Conservation insights and reforestation involvements. For. Ecol. Manag. 2013, 304, 150–161. [Google Scholar] [CrossRef]
- Kreyling, J.; Buhk, C.; Backhaus, S.; Hallinger, M.; Huber, G.; Huber, L.; Jentsch, A.; Konnert, M.; Thiel, D.; Wilmking, M.; et al. Local adaptations to frost in marginal and central populations of the dominant forest tree Fagus sylvatica L. as affected by temperature and extreme drought in common garden experiments. Ecol. Evol. 2014, 4, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Bachofen, C.; Wohlgemuth, T.; Ghazoul, J.; Moser, B. Cold temperature extremes during spring do not limit the range shift of Mediterranean pines into regions with intermittent frost. Funct. Ecol. 2016, 30, 856–865. [Google Scholar] [CrossRef]
- Wang, T.; Wang, Z.; Xia, F.; Su, Y. Local adaptation to temperature and precipitation in naturally fragmented populations of Cephalotaxus oliveri, an endangered conifer endemic to China. Sci. Rep. 2016, 6, 25031. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genet. Resour. Newslett. 2001, 127, 15–19. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Mejnartowicz, L. Genetic analysis of silver-fir populations in the Beskids. Acta Soc. Bot. Pol. 2003, 72, 115–119. [Google Scholar] [CrossRef]
- Kormuták, A.; Kádasi-Horáková, M.; Vooková, B.; Gömöry, D. Genetic structure of Silver fir Primeval Forest in Slovakia. For. J. 2008, 54, 37–42. [Google Scholar]
- Piepho, H.P. Analyzing genotype-environment data by mixed models with multiplicative effects. Biometrics 2016, 53, 761–766. [Google Scholar] [CrossRef]
- Balzarini, M. Applications of mixed models in plant breeding. In Quantitative Genetics, Genomics, and Plant Breeding; Kang, M.S., Ed.; CABI Publishing: New York, NY, USA, 2002; pp. 353–365. ISBN 0851996019. [Google Scholar]
- Ukalski, K.; Klisz, M. Application of GGE biplot graphs in multi-environment trials on selection of forest trees. Folia For. Pol. Ser. A 2016, 58, 228–239. [Google Scholar] [CrossRef]
- Klisz, M.; Jastrzębowski, S.; Ukalski, K.; Ukalska, J.; Przybylski, P. Adaptation of Norway spruce populations in Europe: A case study from northern Poland. N. Z. J. For. Sci. 2017, 47, 8. [Google Scholar] [CrossRef]
- Jastrzębowski, S.; Ukalski, K.; Klisz, M.; Ukalska, J.; Przybylski, P.; Matras, J.; Barzdajn, W.; Kowalkowski, W. Assessment of the height stability in progeny of Fagus sylvatica L. populations using the GGE biplot method. Dendrobiology 2018, 79, 34–46. [Google Scholar] [CrossRef]
- Ukalska, J.; Kociuba, W. Phenotypical diversity of winter triticale genotypes collected in the Polish gene bank between 1982 and 2008 with regard to major quantitative traits. Field Crops Res. 2013, 149, 203–212. [Google Scholar] [CrossRef]
- Hocking, R.R.; Speed, F.M. A Full Rank Analysis of Some Linear Model Problems. J. Am. Stat. Assoc. 1975, 70, 706–712. [Google Scholar] [CrossRef]
- Gabriel, R.K. The biplot graphic display of matrices with application to principal component analysis. Biometrika 1971, 58, 453–467. [Google Scholar] [CrossRef]
- Gabriel, R.K. Analysis of meteorological data by means of canonical decompositions and biplots. J. Appl. Meteorol. 1972, 11, 1071–1077. [Google Scholar] [CrossRef]
- Gabriel, R.K. Least squares approximation of matrices by additive and multiplicative models. J. R. Stat. Soc. Ser. B 1978, 40, 186–196. [Google Scholar]
- Bradu, D.; Gabriel, R.K. The biplot as a diagnostic tool for models of two-way tables. Technometrics 1978, 20, 47–68. [Google Scholar] [CrossRef]
- Dodge, Y. Analysis of Experiments with Missing Data; Willey: New York, NY, USA, 1985; pp. 1–449. ISBN 0471887366. [Google Scholar]
- Dray, S.; Josse, J. Principal component analysis with missing values: A comparative survey of methods. Plant. Ecol. 2015, 216, 657–667. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A. Interpretation of Genotype. Crop Sci. 2001, 41, 19–25. [Google Scholar] [CrossRef]
- Yan, W. Singular-value partitioning in biplot analysis of multienvironment trial data. Agron. J. 2002, 94, 990–996. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT 13.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D. SAS System for Mixed Models; SAS Institute Inc.: Cary, NC, USA, 1996; pp. 1–663. ISBN 1555447791. [Google Scholar]
- Khattree, R.; Naik, D.N. Multivariate Data Reduction and Discrimination with SAS Software; SAS Institute Inc.: Cary, NC, USA, 2000; pp. 1–574. ISBN 1580256961. [Google Scholar]
- Latreille, A.C.; Pichot, C. Local-scale diversity and adaptation along elevational gradients assessed by reciprocal transplant experiments: Lack of local adaptation in silver fir populations. Ann. For. Sci. 2017, 74, 77. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and silviculture of silver fir (Abies alba Mill.): A review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Anic, I.; Vukelic, J.; Mikac, S.; Baksic, D.; Ugarkovic, D. Effects of Global Climate Change on the Ecological Niche of Silver Fir (Abies alba Mill.) in Croatia. Sumar. List 2009, 133, 135–144. [Google Scholar]
- Wang, T.; Campbell, E.M.; O’Neill, G.A.; Aitken, S.N. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. For. Ecol. Manag. 2012, 279, 128–140. [Google Scholar] [CrossRef]
- Lewandowski, A.; Litkowiec, M.; Grygier, A. Verification of the origin of Norway spruce (Picea abies) stands in the Gołdap Forest District. Sylwan 2012, 156, 494–501, (In Polish with English abstract). [Google Scholar]
- Lewandowski, A.; Szydlarski, M.; Litkowiec, M. Origin of Norway spruce (Picea abies (L.) Karst.) in the Kartuzy Forest District. Sylwan 2014, 158, 509–515, (In Polish with English abstract). [Google Scholar]
- Bolibok, L.; Zajączkowski, J.; Dobrowolska, D.; Mionskowski, M. Potential climatic range for Abies alba Mill. in Poland. Sylwan 2016, 160, 519–528, (In Polish with English abstract). [Google Scholar]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Sabor, J.; Skrzyszewska, K.; Banach, J. All-Poland provenance test Jd PL 86/90 comparative plot with Silver fir in the Baligród Forest District. Zesz. Nauk. Akad. Rol. Krakowie 1999, 61, 263–280, (In Polish with English abstract). [Google Scholar]
- Willi, Y.; Van Buskirk, J.; Hoffmann, A.A. Limits to the Adaptive Potential of Small Populations. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 433–458. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population ID | Forest Base Material ID | Forest District | Production Cycle (Year/Year) | Elevation (m a.s.l.) | Latitude | Longitude |
|---|---|---|---|---|---|---|
| BAL1 | 11625 | Baligród | 2/1 | 631 | 49.3226 | 22.2699 |
| BAL2 | 11626 | Baligród | 1.5/1.5 | 624 | 49.3226 | 22.1912 |
| BIR1 | 27146 | Bircza | 1.5/1.5 | 506 | 49.5667 | 22.5713 |
| KRA1 | 11270 | Krasiczyn | 2/1 | 399 | 49.6918 | 22.5931 |
| KRA2 | 11271 | Krasiczyn | 1.5/1.5 | 374 | 49.6919 | 22.5961 |
| KRA3 | 11273 | Krasiczyn | 1.5/1.5 | 365 | 49.7168 | 22.6460 |
| LES1 | 30320 | Lesko | 1.5/1.5 | 456 | 49.4862 | 22.1964 |
| LES2 | 30326 | Lesko | 1.5/1.5 | 363 | 49.5056 | 22.2871 |
| LES3 | 30327 | Lesko | 1.5/1.5 | 491 | 49.5150 | 22.4125 |
| LES4 | 30328 | Lesko | 2/1 | 450 | 49.4548 | 22.3039 |
| LES5 | 30329 | Lesko | 1.5/1.5 | 499 | 49.4590 | 22.2859 |
| LES6 | 30330 | Lesko | 2/1 | 396 | 49.4489 | 22.3366 |
| LUT1 | 30205 | Lutowiska | 1.5/1.5 | 516 | 49.3151 | 22.5550 |
| LUT2 | 30212 | Lutowiska | 1.5/1.5 | 599 | 49.3129 | 22.6590 |
| LZD | MP/2/45114/06 | LZD Krynica | 2/1 | 670 | 49.3750 | 20.9870 |
| NAR | 10995 | Narol | 1.5/1.5 | 314 | 50.3837 | 23.3674 |
| RYM1 | 36145 | Rymanów | 1.5/1.5 | 580 | 49.3810 | 21.9499 |
| RYM2 | 36146 | Rymanów | 1.5/1.5 | 536 | 49.4380 | 21.8773 |
| STU | 10691 | Stuposiany | 1.5/1.5 | 688 | 49.1613 | 22.6977 |
| Population | Mean Height (cm) (CV (%)) | Mean Survival (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BIR | KOM | CIS | LES | Across Sites | BIR | KOM | CIS | LES | Across Sites | |
| BAL2 | 55 (12) | 52 (14) | 58 (13) | 50 (6) | 54 (12) | 93.3 | 90.5 | 98.5 | 94.8 | 94.3 |
| BIR1 | 57 (13) | 57 (5) | 59 (10) | 55 (10) | 57 (9) | 93.5 | 95.8 | 96.3 | 93.0 | 94.7 |
| KRA1 | 38 (36) | 30 (46) | 55 (14) | 44 (4) | 42 (32) | 77.7 | 90.9 | 95.2 | 94.7 | 89.6 |
| KRA2 | 52 (26) | 55 (11) | 61 (9) | 51 (2) | 55 (15) | 87.8 | 90.0 | 96.3 | 88.3 | 90.6 |
| LES1 | 53 (29) | 63 (7) | 49 (14) | 48 (17) | 53 (20) | 92.8 | 93.8 | 93.5 | 94.0 | 93.5 |
| LES2 | 57 (11) | 56 (11) | 53 (15) | 57 (10) | 55 (11) | 94.8 | 88.5 | 94.5 | 91.0 | 92.2 |
| LES3 | 61 (10) | 54 (16) | 61 (9) | 55 (14) | 57 (13) | 96.3 | 84.5 | 96.0 | 94.3 | 92.8 |
| LES5 | 55 (28) | 56 (4) | 60 (4) | 51 (13) | 56 (15) | 93.3 | 90.3 | 96.0 | 83.6 | 90.8 |
| LUT1 | 55 (18) | 53 (6) | 56 (15) | 53 (16) | 54 (13) | 94.8 | 91.0 | 98.0 | 94.3 | 94.5 |
| LUT2 | 57 (19) | 54 (14) | 64 (2) | 52 (7) | 57 (13) | 94.8 | 89.8 | 97.3 | 95.3 | 94.3 |
| LZD | 42 (10) | 46 (16) | 48 (5) | 38 (38) | 44 (18) | 88.8 | 88.8 | 98.3 | 92.3 | 92.1 |
| NAR | 55 (13) | 55 (6) | 54 (5) | 51 (8) | 54 (8) | 91.3 | 92.0 | 95.3 | 95.0 | 93.4 |
| RYM1 | 57 (8) | 58 (13) | 58 (11) | 56 (7) | 57 (9) | 94.5 | 91.5 | 98.0 | 93.5 | 94.4 |
| RYM2 | 52 (14) | 49 (9) | 57 (4) | 46 (14) | 51 (12) | 92.0 | 87.0 | 95.8 | 94.8 | 92.4 |
| STU | 56 (6) | 58 (10) | 60 (14) | 47 (26) | 55 (16) | 94.8 | 94.8 | 96.5 | 89.0 | 93.8 |
| BAL1 | 30 (63) | 63 (16) | 76 (16) | 59 (38) | 55.1 | 91.8 | 93.3 | 80.1 | ||
| KRA3 | 66 (11) | 56 (12) | 50 (16) | 58 (17) | 96.3 | 89.8 | 92.5 | 92.9 | ||
| LES4 | 49 (16) | 28 (48) | 39 (17) | 39 (33) | 92.3 | 91.8 | 95.0 | 93.0 | ||
| LES6 | 45 (29) | 61 (35) | 89 (11) | 65 (36) | 87.1 | 80.3 | 95.5 | 87.6 | ||
| Multi-population mean | 53 (22) | 53 (23) | 57 (12) | 53 (25) | 5 4 (21) | 90.1 | 90.2 | 96.4 | 92.9 | 91.9 |
| Bioclimatic Variables | Abbreviation | PC 1 | PC 2 | PC 3 |
|---|---|---|---|---|
| Altitude | Alt | 0.99 | 0.04 | −0.05 |
| Annual Mean Temperature | bio1 | −0.88 | 0.42 | 0.10 |
| Mean Monthly Temperature Range | bio2 | 0.07 | 0.81 | 0.49 |
| Isothermality (BIO2/BIO7) (× 100) | bio3 | 0.45 | 0.85 | −0.11 |
| Temperature Seasonality (STD × 100) | bio4 | −0.67 | −0.51 | 0.52 |
| Max Temperature of Warmest Month | bio5 | −0.87 | 0.33 | 0.34 |
| Min Temperature of Coldest Month | bio6 | −0.56 | 0.63 | −0.47 |
| Temperature Annual Range (5–6) | bio7 | −0.56 | −0.14 | 0.81 |
| Mean Temperature of Wettest Quarter | bio8 | −0.92 | 0.25 | 0.25 |
| Mean Temperature of Driest Quarter | bio9 | −0.61 | 0.76 | −0.17 |
| Mean Temperature of Warmest Quarter | bio10 | −0.92 | 0.25 | 0.25 |
| Mean Temperature of Coldest Quarter | bio11 | −0.51 | 0.80 | −0.24 |
| Annual Precipitation | bio12 | 0.91 | 0.34 | 0.19 |
| Precipitation of Wettest Month | bio13 | 0.85 | 0.46 | 0.20 |
| Precipitation of Driest Month | bio14 | 0.94 | 0.05 | 0.14 |
| Precipitation Seasonality (CV) | bio15 | −0.32 | 0.71 | 0.04 |
| Precipitation of Wettest Quarter | bio16 | 0.85 | 0.47 | 0.21 |
| Precipitation of Driest Quarter | bio17 | 0.94 | 0.10 | 0.20 |
| Precipitation of Warmest Quarter | bio18 | 0.85 | 0.47 | 0.21 |
| Precipitation of Coldest Quarter | bio19 | 0.93 | 0.04 | 0.21 |
| Eigenvalue | 11.87 | 4.97 | 2.02 | |
| Variance explained | 59.37 | 24.85 | 10.08 |
| Source of Variation | Df | Mean Square | F | p | % (G + E + GE) |
|---|---|---|---|---|---|
| Genotype (G) | 18 | 518.08 | 2.11 | 0.019 | 41.27 |
| Site (E) | 3 | 318.56 | 0.69 | 0.564 | 4.23 |
| Block (Site) | 12 | 274.96 | 4.79 | <0.001 | |
| Genotype × Site (GE) | 50 | 246.31 | 4.29 | <0.001 | 54.50 |
| Experimental error | 201 | 57.38 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klisz, M.; Ukalski, K.; Ukalska, J.; Jastrzębowski, S.; Puchałka, R.; Przybylski, P.; Mionskowski, M.; Matras, J. What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments? Forests 2018, 9, 441. https://doi.org/10.3390/f9070441
Klisz M, Ukalski K, Ukalska J, Jastrzębowski S, Puchałka R, Przybylski P, Mionskowski M, Matras J. What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments? Forests. 2018; 9(7):441. https://doi.org/10.3390/f9070441
Chicago/Turabian StyleKlisz, Marcin, Krzysztof Ukalski, Joanna Ukalska, Szymon Jastrzębowski, Radosław Puchałka, Paweł Przybylski, Marcin Mionskowski, and Jan Matras. 2018. "What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments?" Forests 9, no. 7: 441. https://doi.org/10.3390/f9070441
APA StyleKlisz, M., Ukalski, K., Ukalska, J., Jastrzębowski, S., Puchałka, R., Przybylski, P., Mionskowski, M., & Matras, J. (2018). What Can We Learn from an Early Test on the Adaptation of Silver Fir Populations to Marginal Environments? Forests, 9(7), 441. https://doi.org/10.3390/f9070441