Shifts in Growing Season of Tropical Deciduous Forests as Driven by El Niño and La Niña during 2001–2016

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
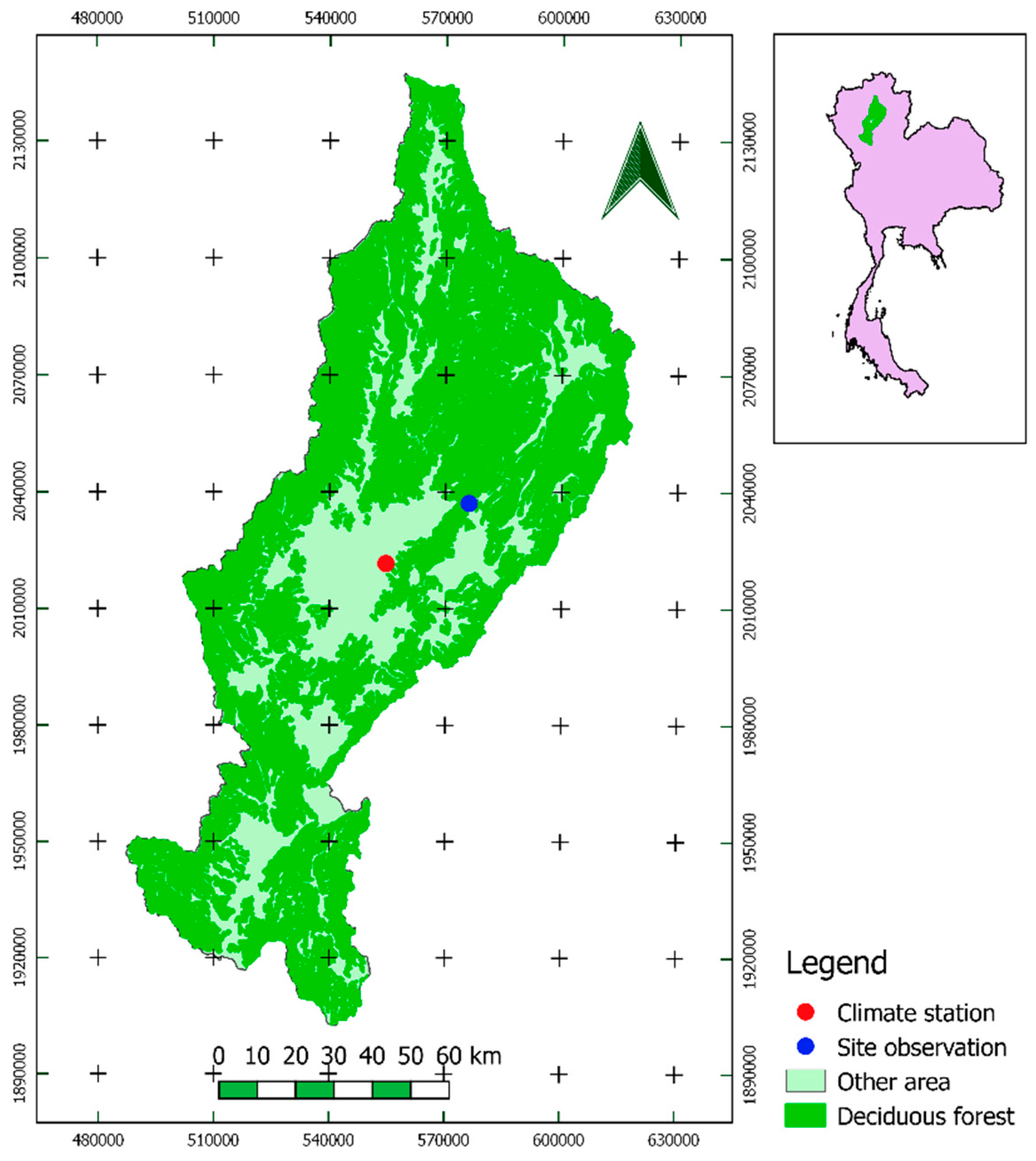
2.1. Study Area
2.2. Data Set
2.2.1. MODIS Fata
2.2.2. Landsat Imagery
2.2.3. Digital Elevation Models
2.2.4. Climate Variables
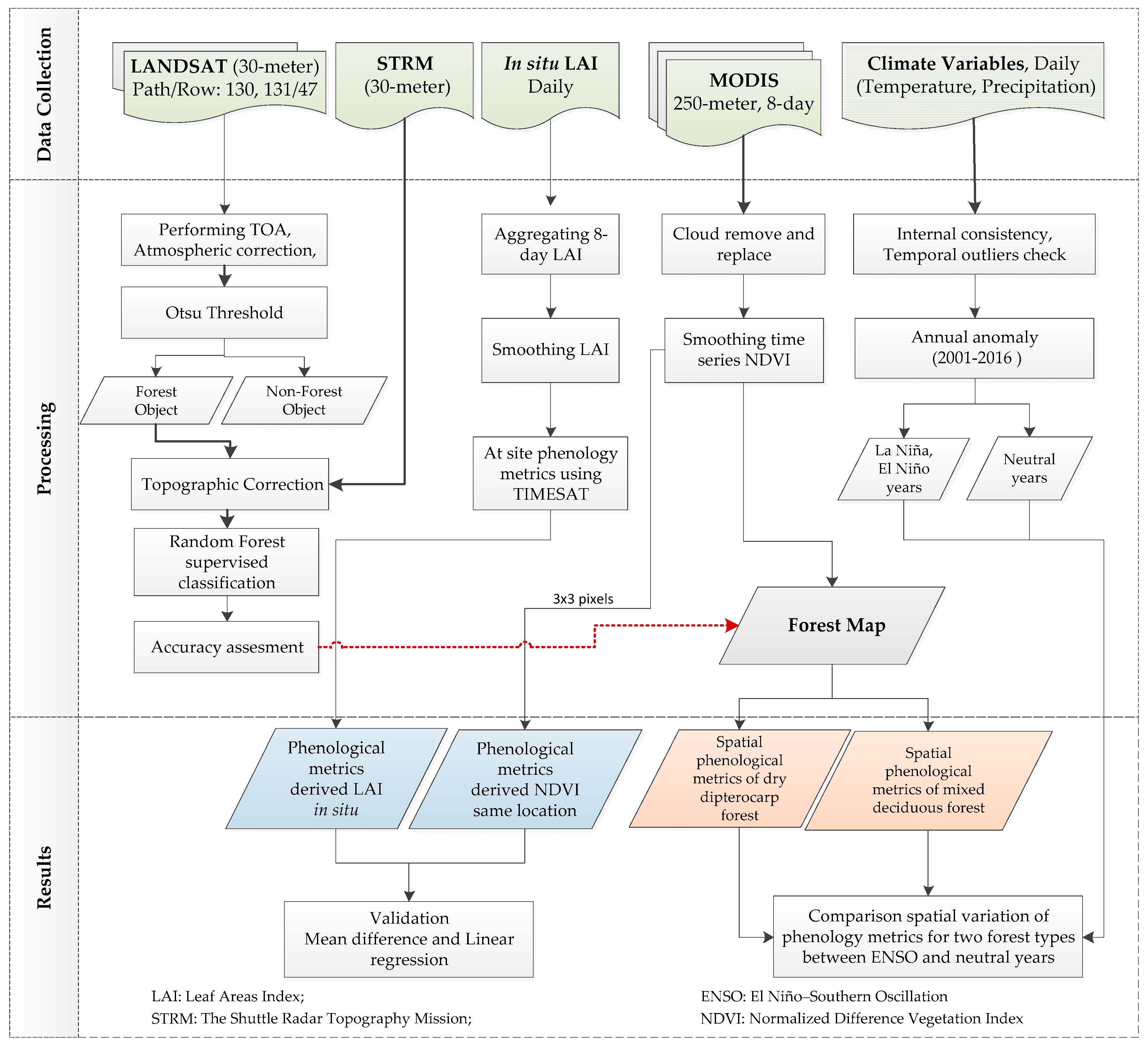
2.3. Methods
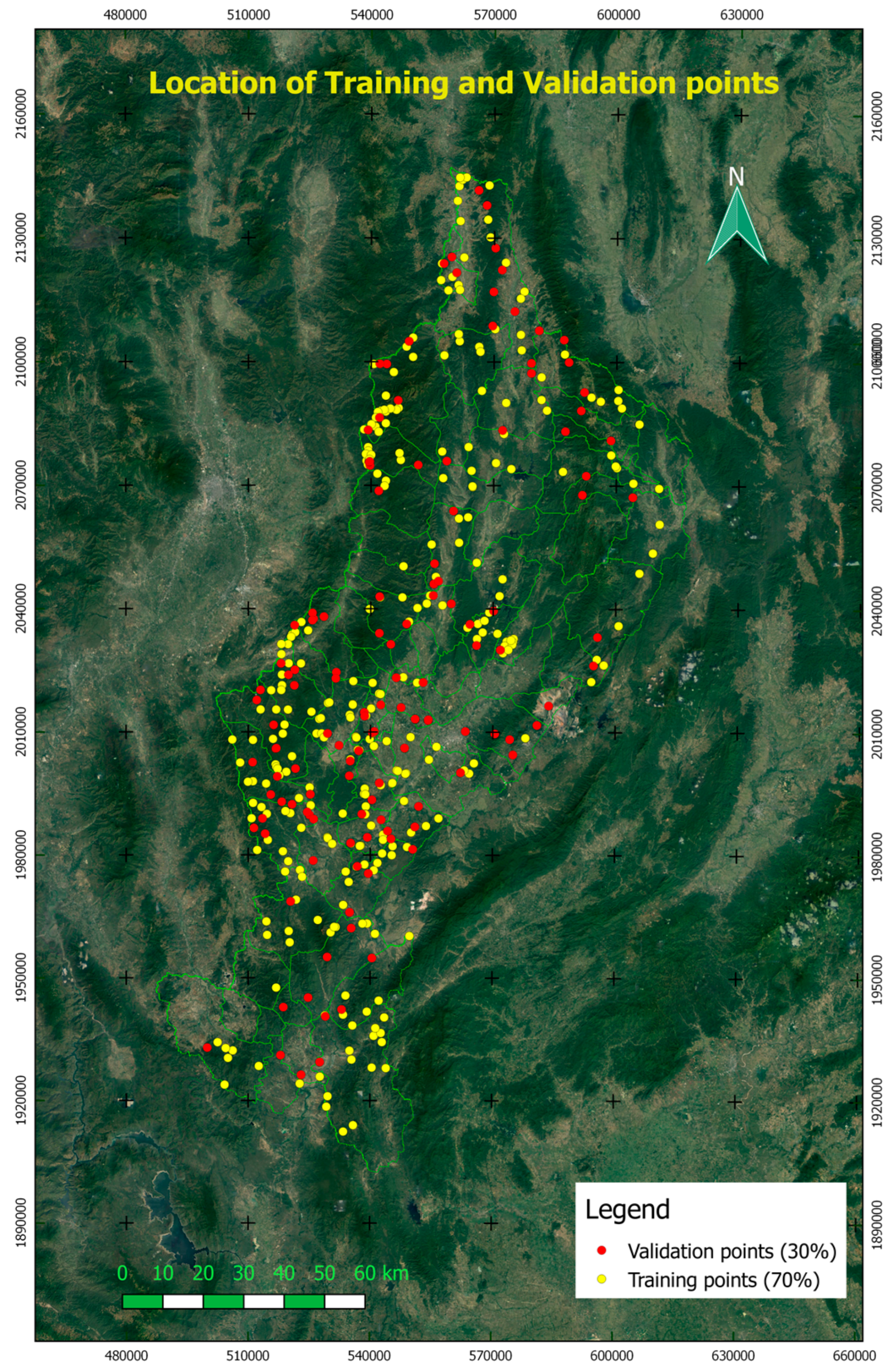
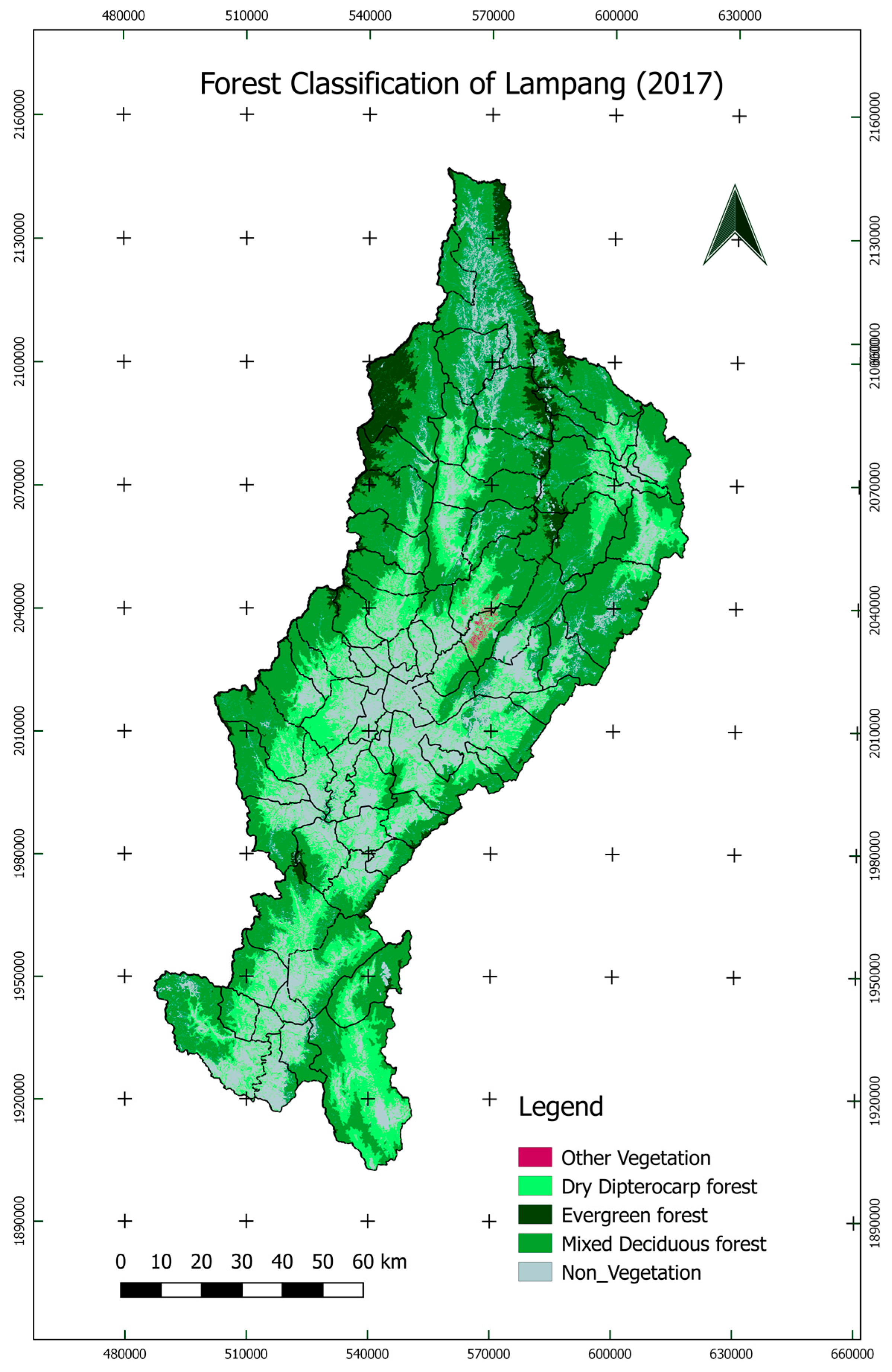
2.3.1. Forest Classification in Lampang
2.3.2. NDVI Time Series for Phenological Analysis
2.3.3. Determination of Forest Phenological Variables from Satellite
- Start of growing season (SOS): This is defined as the date of leaf unfolding (day of year, DOY) and this study considered SOS as a date when NDVI of the left edge has increases 20 percentage measured from the left minimum point.
- End of the season (EOS): This is defined as the dates of leaf discoloration (day of year, DOY) and leaf fall at the end of season. This study considered EOS as a date when NDVI of the right edge has decreases to 20 percentage of the right minimum level.
- Length of the season (LOS): This is the duration (number of days) from the start to the end of the season.
2.3.4. Determination of in situ Derived Forest Phenological Metrics
2.3.5. Validation of the Phenological Metrics
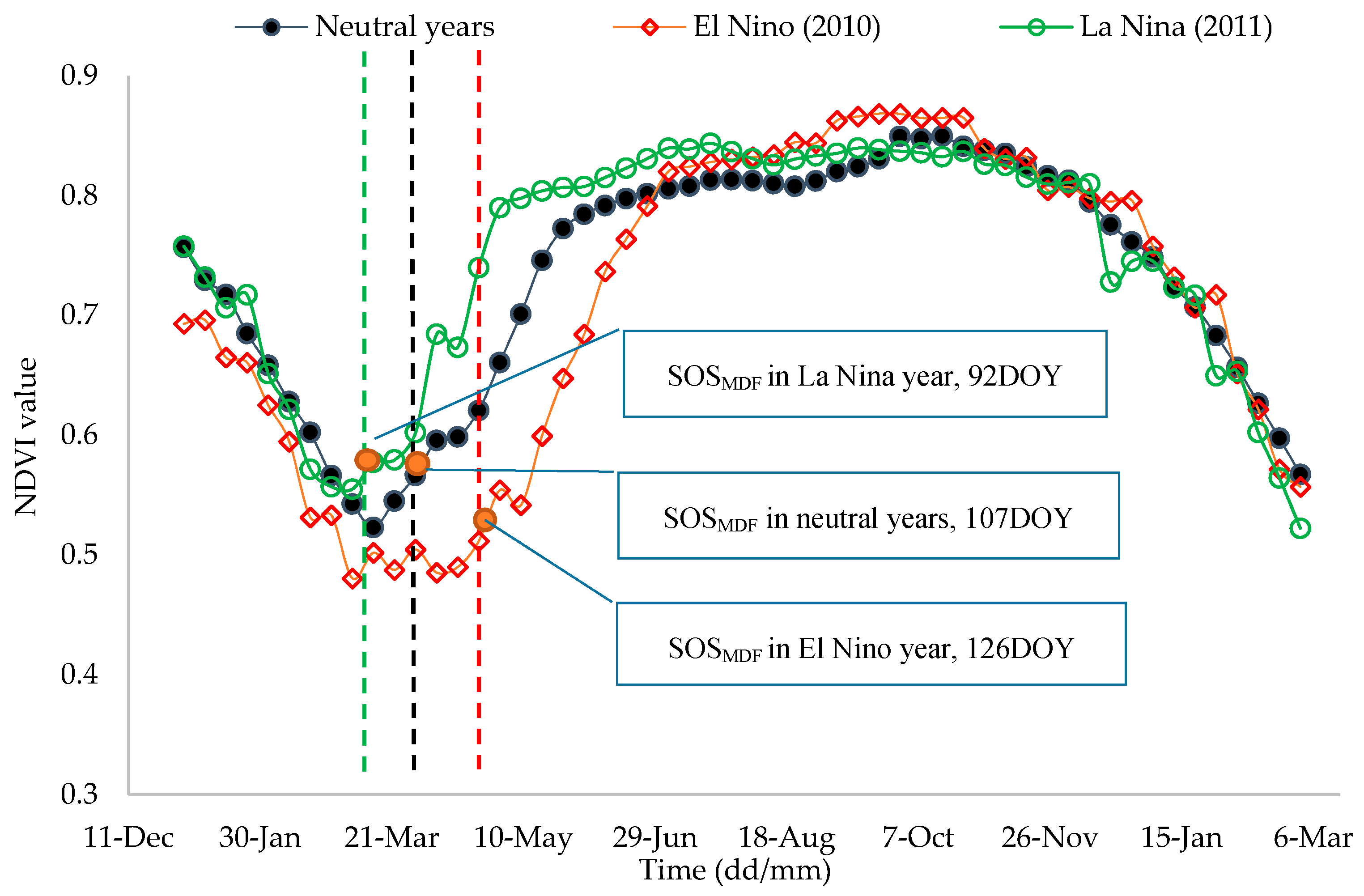
2.3.6. Assessing the ENSO-Related Patterns in Annual Phenological Metrics
3. Results and Discussion
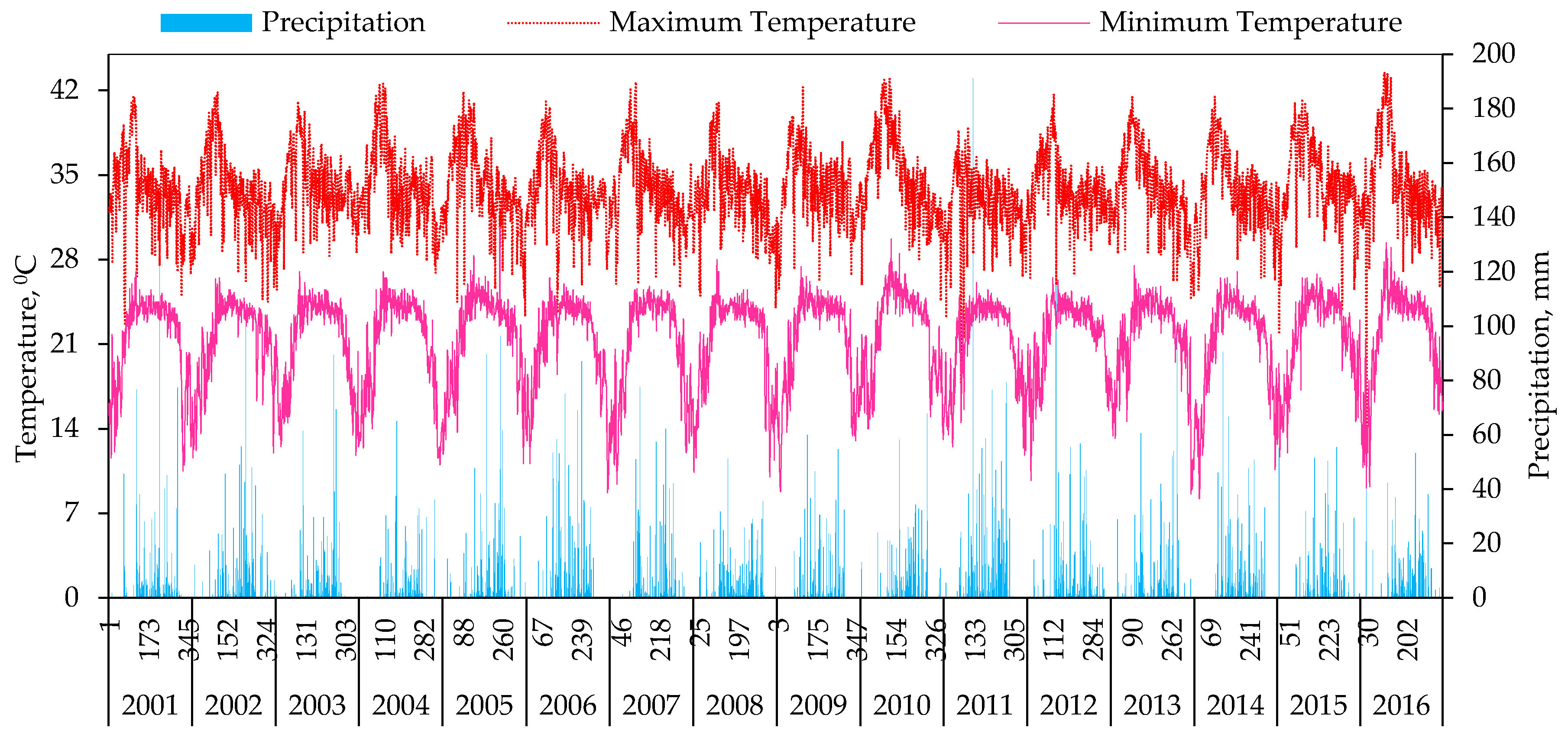
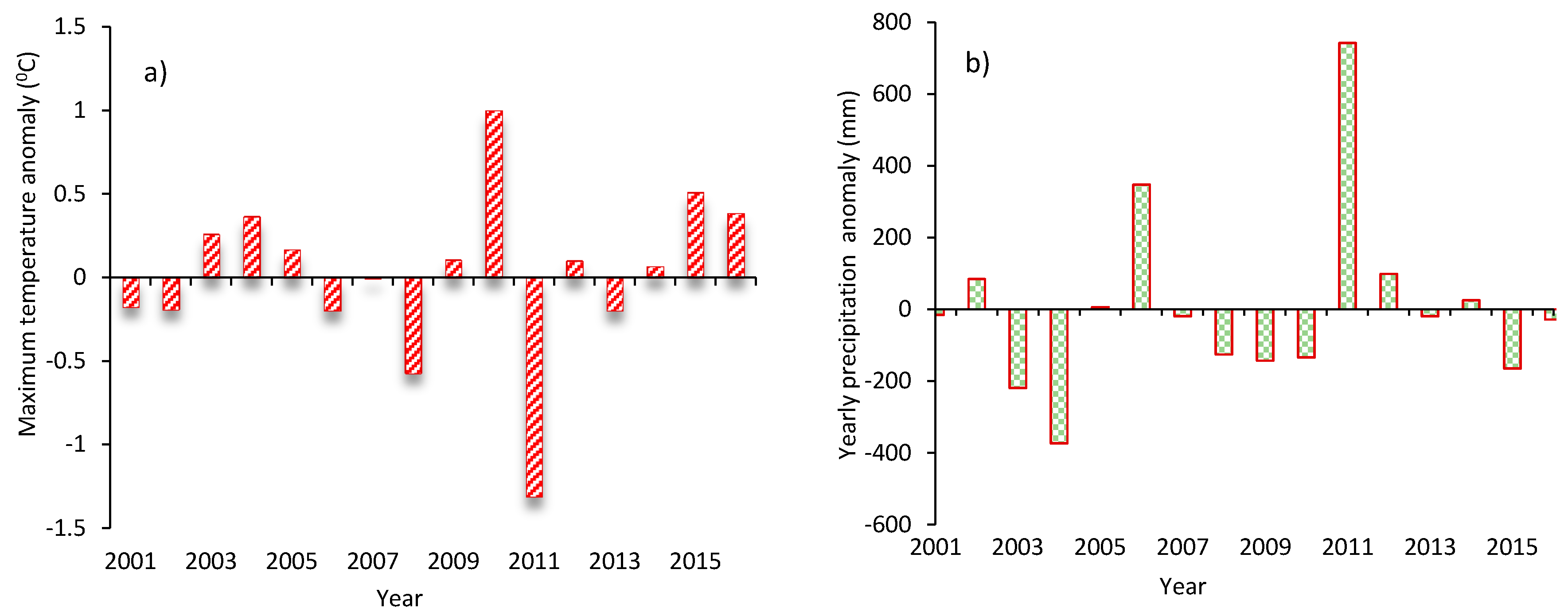
3.1. Variations in Temperature and Precipitation during 2001–2016
3.2. Forest Classification
3.3. Variations in NDVI during 2001–2016
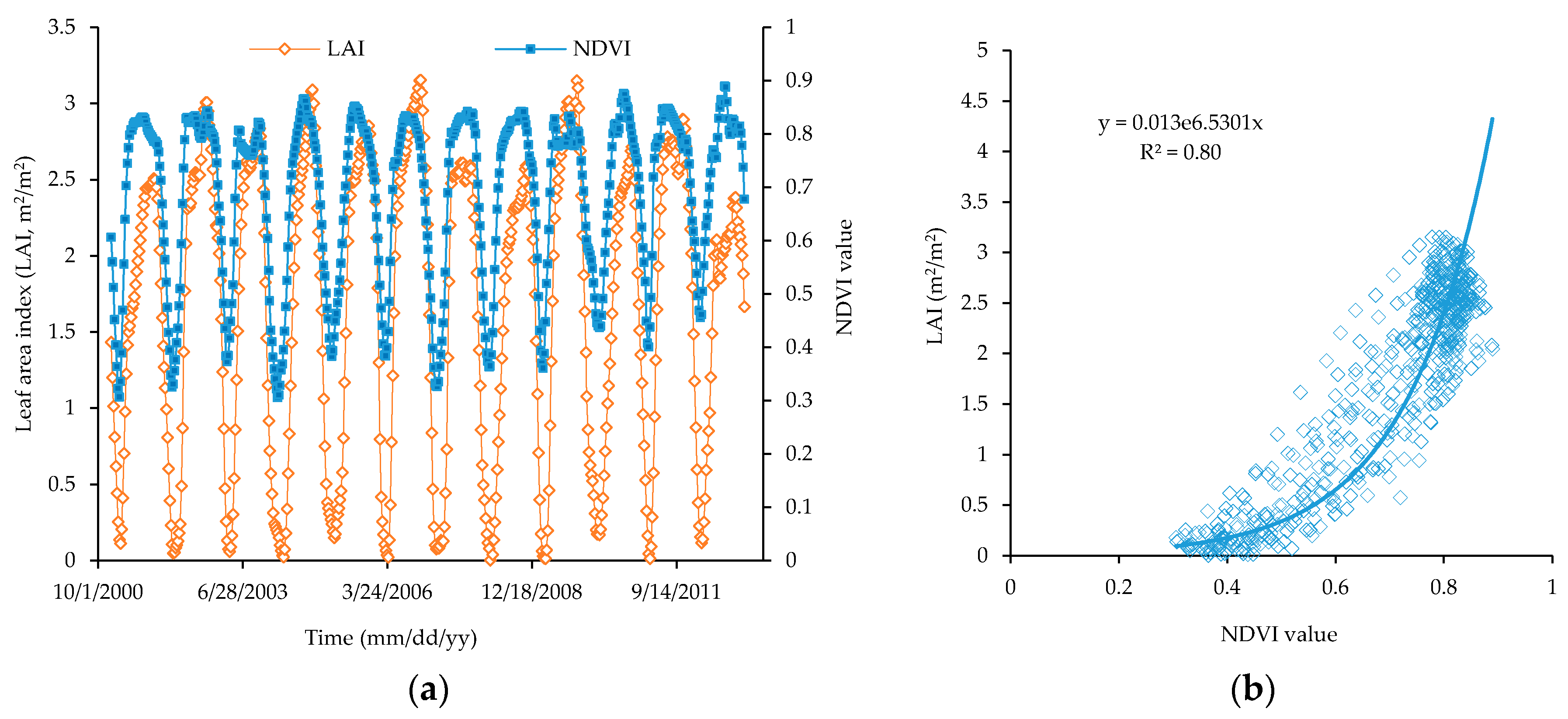
3.3.1. Relationship between Satellite-Based NDVI and Observed LAI of Teak Forest Plantation
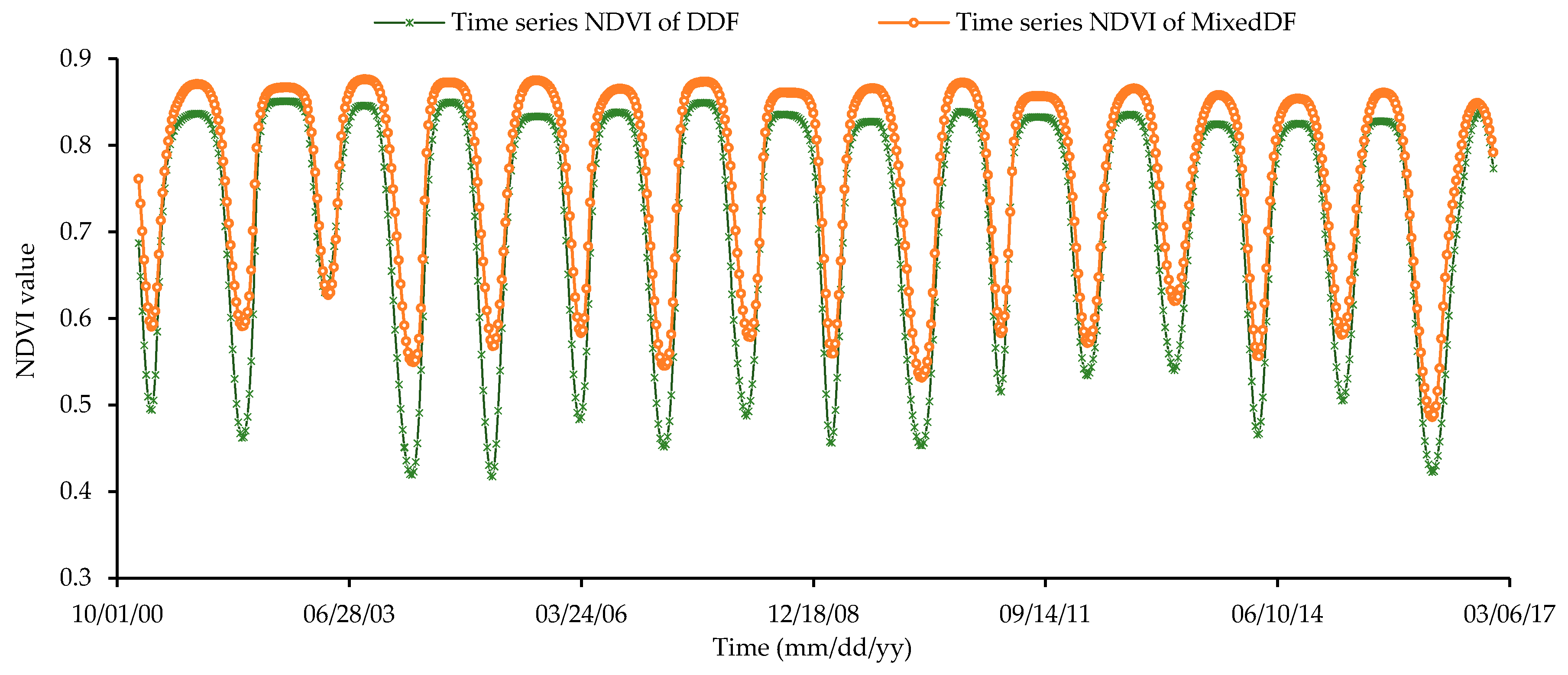
3.3.2. Temporal Variations of NDVI in Lampang Province during 2001–2016
3.4. Variations of Phenological Metrics and the Effects of ENSO
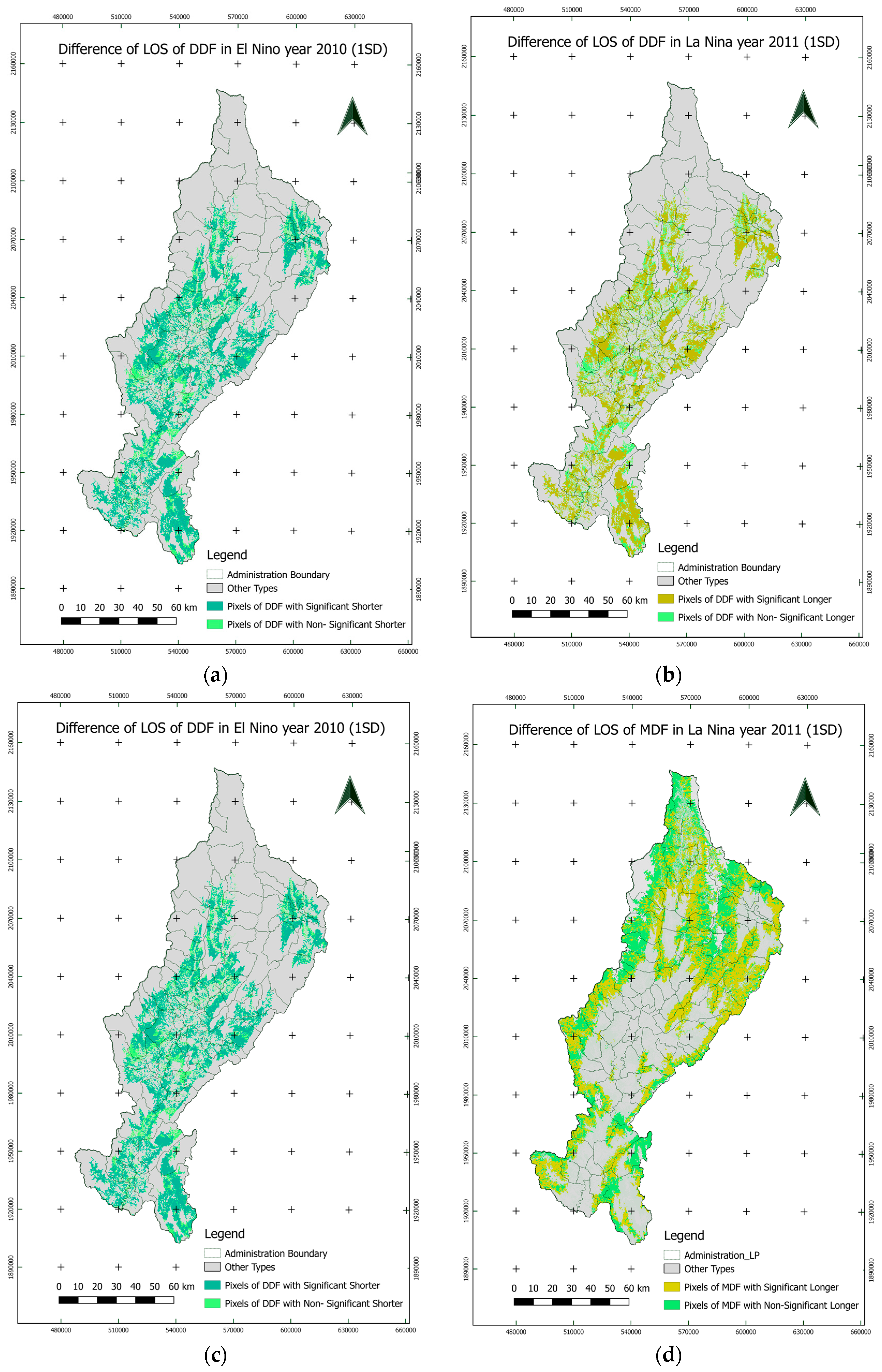
3.4.1. Temporal Variations
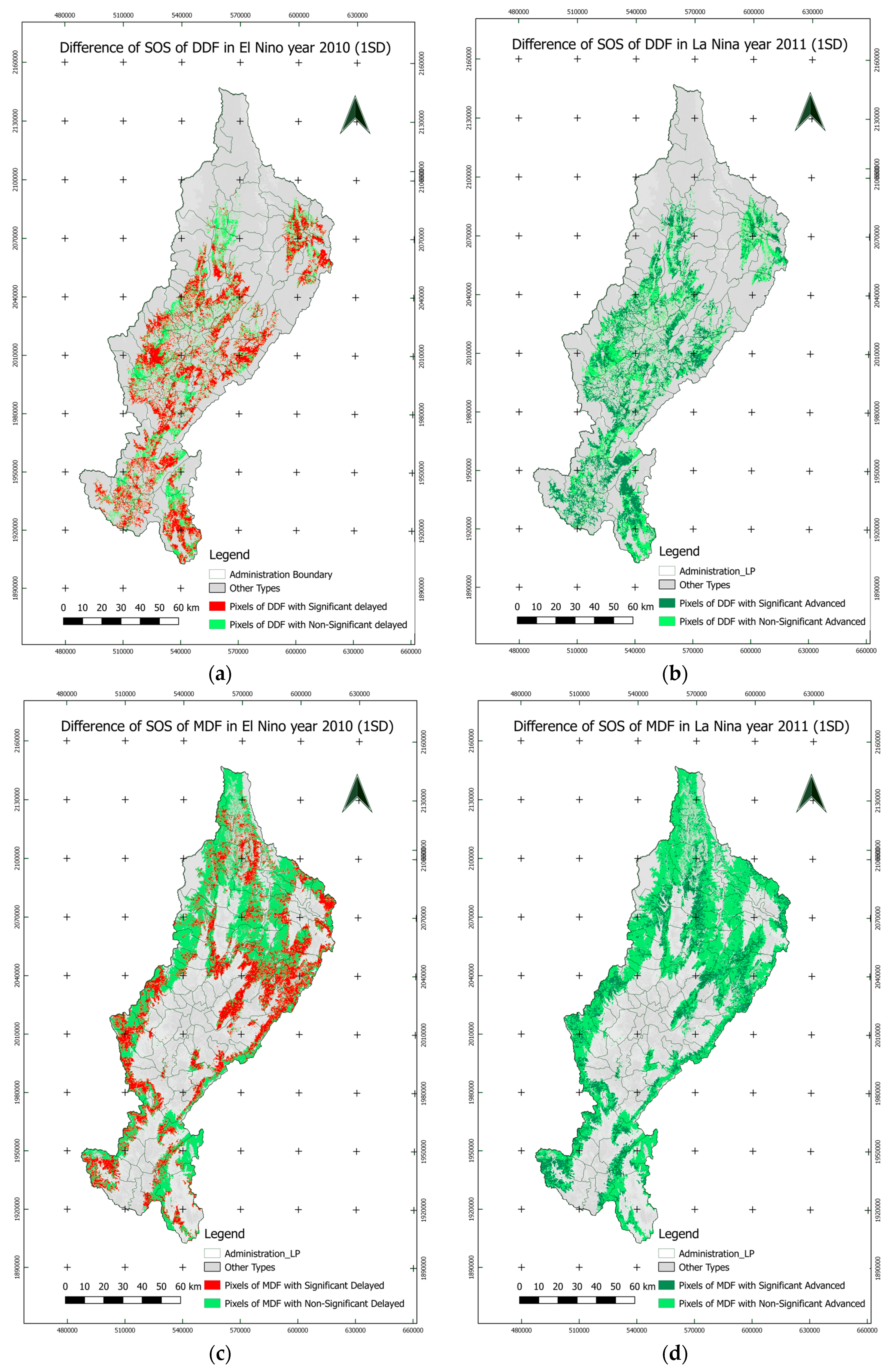
3.4.2. Spatial Variations
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- U.S. DOE. Research Priorities for Tropical Ecosystems Under Climate Change Workshop Report; DOE/SC-0153; U.S. Department of Energy Office of Science: Washington, DC, USA, 2012.
- European Climate Foundation. The UN Intergovernmental Panel on Climate Change (IPCC) Fifth Assessment Report (AR5): Implications for Business; European Climate Foundation: Cambridge, UK, 2013. [Google Scholar]
- Nakagawa, M.; Tanaka, K.; Nakashizuka, T.; Ohkubo, T.; Kato, T.; Maeda, T.; Sato, K.; Miguchi, H.; Nagamasu, H.; Ogino, K.; et al. Impact of Severe Drought Associated with the 1997–1998 El Niño in a Tropical Forest in Sarawak. J. Trop. Ecol. 2000, 16, 355–367. [Google Scholar] [CrossRef]
- Asner, G.P.; Townsend, A. Satellite Observation of El Nino Effects Effects on Amazon Forest Phenology and Productivity. Geophys. Res. Lett. 2000, 27, 981–984. [Google Scholar] [CrossRef]
- FAO. Forest Resources Assessment 1990—Non-Tropical Developing Countries Mediterranean Region; FAO: Rome, Italy, 1990. [Google Scholar]
- Cavaleri, M.A.; Coble, A.P.; Ryan, M.G.; Bauerle, W.L.; Loescher, H.W.; Oberbauer, S.F. Tropical Rainforest Carbon Sink Declines during El Niño as a Result of Reduced Photosynthesis and Increased Respiration Rates. New Phytol. 2017, 216, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Rolim, S.G.; Jesus, R.M.; Nascimento, H.E.M.; Do Couto, H.T.Z.; Chambers, J.Q. Biomass Change in an Atlantic Tropical Moist Forest: The ENSO Effect in Permanent Sample Plots over a 22-Year Period. Oecologia 2005, 142, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, M.; Scheffer, M.; Ezcurra, E.; Gutiérrez, J.R.; Mohren, G.M.J. El Niño Effects on the Dynamics of Terrestrial Ecosystems. Trends Ecol. Evol. 2001, 16, 89–94. [Google Scholar] [CrossRef]
- Pau, S.; Okin, G.S.; Gillespie, T.W. Asynchronous Response of Tropical Forest Leaf Phenology to Seasonal and El Niño-Driven Drought. PLoS ONE 2010, 5, e11325. [Google Scholar] [CrossRef] [PubMed]
- Slik, J.W.F. El Nino Droughts and Their Effects on Tree Species Composition and Diversity in Tropical Rain Forests. Oecologia 2004, 141, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Patra, P.K.; Ishizawa, M.; Maksyutov, S.; Nakazawa, T.; Inoue, G. Role of Biomass Burning and Climate Anomalies for Land-Atmosphere Carbon Fluxes Based on Inverse Modeling of Atmospheric CO2. Global Biogeochem. Cycles 2005, 19, 1–10. [Google Scholar] [CrossRef]
- Li, J.; Fan, K.; Xu, Z. Asymmetric Response in Northeast Asia of Summer NDVI to the Preceding ENSO Cycle. Clim. Dyn. 2016, 47, 2765–2783. [Google Scholar] [CrossRef]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate Change, Phenology, and Phenological Control of Vegetation Feedbacks to the Climate System. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Saigusa, N.; Yamamoto, S.; Hirata, R.; Ohtani, Y.; Ide, R.; Asanuma, J.; Gamo, M.; Hirano, T.; Kondo, H.; Kosugi, Y.; et al. Temporal and Spatial Variations in the Seasonal Patterns of CO2 Flux in Boreal, Temperate, and Tropical Forests in East Asia. Agric. For. Meteorol. 2008, 148, 700–713. [Google Scholar] [CrossRef]
- Nanda, A.; Prakash, H.M.; Suresh, H.S.; Murthy, Y.L.K. Canopy, Understorey Leaf Phenology and Seasonality in Tropical Dry Forest, Southern India. Adv. For. Lett. 2016, 5. [Google Scholar] [CrossRef]
- Igarashi, Y.; Kumagai, T.; Yoshifuji, N.; Sato, T.; Tanaka, N.; Tanaka, K.; Suzuki, M.; Tantasirin, C. Environmental Control of Canopy Stomatal Conductance in a Tropical Deciduous Forest in Northern Thailand. Agric. For. Meteorol. 2015, 202, 1–10. [Google Scholar] [CrossRef]
- Royal Forest Department of Thailand. Forestry in Thailand; Royal Forest Department of Thailand: Bangkok, Thailand, 2009. [Google Scholar]
- Vaidhayakarn, C.; Maxwell, J.F. Ecological Status of the Lowland Deciduous Forest in Chang Kian. J. Sci. Technol. 2010, 4, 268–317. [Google Scholar]
- Kirtphaiboon, S.; Wongwises, P.; Limsakul, A.; Sooktawee, S.; Humphries, U. Rainfall Variability over Thailand Related to the El Nino-Southern Oscillation (ENSO). J. Sustain. Energy Environ. 2014, 5, 37–42. [Google Scholar]
- Maxwell, J.F.; Elliot, S.; Anusarnsunthorn, V. The Vegetation of Jae Sawn National Park, Lampang Province, Thailand. Nat. Hist. Bull. Siam Soc. 1997, 45, 71–97. [Google Scholar]
- Land Processes Distributed Active Archive Center - LP DAAC, USGS. Available online: https://lpdaac.usgs.gov/products/modis_products_table/mod09q1 (accessed on 01 July 2017).
- Land Processes Distributed Active Archive Center - LP DAAC, USGS. Available online: https://lpdaac.usgs.gov/products/modis_products_table/mod09a1 (accessed on 01 July 2017).
- Hoan, N.T.; Tateishi, R. Global MODIS 250 M Dataset for 10 Years (2003–2012) User’s Manual; Chiba University: Chiba, Japan, 2013. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- The Land Processes Distributed Active Archive Center. Available online: https://lpdaac.usgs.gov/data_ Access/glovis (accessed on 10 April 2017).
- The Shuttle Radar Topographic Mission (SRTM). Available online: https://lta.cr.usgs.gov/SRTM (accessed on 22 October 2017).
- Wu, Q.; Jin, Y.; Fan, H. Evaluating and Comparing Performances of Topographic Correction Methods Based on Multi-Source DEMs and Landsat-8 OLI Data. Int. J. Remote Sens. 2016, 37, 4712–4730. [Google Scholar] [CrossRef]
- Pimple, U.; Sitthi, A.; Simonetti, D.; Pungkul, S.; Leadprathom, K.; Chidthaisong, A. Topographic Correction of Landsat TM-5 and Landsat OLI-8 Imagery to Improve the Performance of Forest Classification in the Mountainous Terrain of Northeast Thailand. Sustainability 2017, 9, 258. [Google Scholar] [CrossRef]
- Feng, S.; Hu, Q.; Qian, W. Quality Control of Daily Meteorological Data in China, 1951–2000: A New Dataset. Int. J. Climatol. 2004, 24, 853–870. [Google Scholar] [CrossRef]
- Peterson, T.C.; Russell, V.; Richard, S.; Vyachevslav, R. Global Historical Climatology Network (GHCN) Quality Control of Monthly Temperature Data. Int. J. Climatol. 1998, 1179, 1169–1179. [Google Scholar] [CrossRef]
- Friedl, M.A.; Gray, J.M.; Melaas, E.K.; Richardson, A.D.; Hufkens, K.; Keenan, T.F.; Bailey, A.; O’Keefe, J.A. Tale of Two Springs: Using Recent Climate Anomalies to Characterize the Sensitivity of Temperate Forest Phenology to Climate Change. Environ. Res. Lett. 2014, 9, 054006. [Google Scholar] [CrossRef]
- Wolf, S.; Baldocchi, D.; Wolf, S.; Keenan, T.F.; Fisher, J.B.; Baldocchi, D.D.; Desai, A.R. Warm Spring Reduced Carbon Cycle Impact of the 2012 US Summer Drought Warm Spring Reduced Carbon Cycle Impact of the 2012 US. Summer Drought. 2016, 113, 5880–5885. [Google Scholar]
- Bruce, C.M.; Hilbert, D.W. Pre-Processing Methodology for Application to Landsat TM/ETM + Imagery of the Wet Tropics; Cooperative Research Centre for Tropical Rainforest Ecology and Management, Rainforest CRC: Cairns, Australia, 2004. [Google Scholar]
- Foga, S.; Scaramuzza, P.L.; Guo, S.; Zhu, Z.; Dilley, R.D.; Beckmann, T.; Schmidt, G.L.; Dwyer, J.L.; Joseph Hughes, M.; Laue, B. Cloud Detection Algorithm Comparison and Validation for Operational Landsat Data Products. Remote Sens. Environ. 2017, 194, 379–390. [Google Scholar] [CrossRef]
- Zhu, Z.; Woodcock, C.E. Object-Based Cloud and Cloud Shadow Detection in Landsat Imagery. Remote Sens. Environ. 2012, 118, 83–94. [Google Scholar] [CrossRef]
- Robinson, N.P.; Allred, B.W.; Jones, M.O.; Moreno, A.; Kimball, J.S.; Naugle, D.E.; Erickson, T.A.; Richardson, A.D. A Dynamic Landsat Derived Normalized Difference Vegetation Index (NDVI) Product for the Conterminous United States. Remote Sens. 2017, 9, 863. [Google Scholar] [CrossRef]
- Pimple, U.; Simonetti, D.; Sitthi, A.; Pungkul, S.; Leadprathom, K.; Skupek, H.; Som-ard, J.; Gond, V.; Towprayoon, S. Google Earth Engine Based Three Decadal Landsat Imagery Analysis for Mapping of Mangrove Forests and Its Surroundings in the Trat Province of Thailand. J. Comput. Commun. 2018, 6, 247–264. [Google Scholar] [CrossRef]
- Otsu, N.A. Threshold Selection Method from Gray-Level Histograms. IEEE Trans. Syst. Man. Cybern. 1979, 9, 62–66. [Google Scholar] [CrossRef]
- Millard, K.; Richardson, M. On the Importance of Training Data Sample Selection in Random Forest Image Classification: A Case Study in Peatland Ecosystem Mapping. Remote Sens. 2015, 7, 8489–8515. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, P.; Foody, G.M.; Herold, M.; Stehman, S.V.; Woodcock, C.E.; Wulder, M.A. Good Practices for Estimating Area and Assessing Accuracy of Land Change. Remote Sens. Environ. 2014, 148, 42–57. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT—A Program for Analyzing Time-Series of Satellite Sensor Data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Xu, L.; Li, B.; Yuan, Y.; Gao, X.; Zhang, T. A Temporal-Spatial Iteration Method to Reconstruct NDVI Time Series Datasets. Remote Sens. 2015, 7, 8906–8924. [Google Scholar] [CrossRef] [Green Version]
- Suepa, T.; Qi, J.; Lawawirojwong, S.; Messina, J.P. Understanding Spatio-Temporal Variation of Vegetation Phenology and Rainfall Seasonality in the Monsoon Southeast Asia. Environ. Res. 2016, 147, 621–629. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Fan, G.F.; Zhang, X.W.; Li, Z.Q.; Gao, D.W. Vegetation Phenological Variation and Its Response to Climate Changes in Zhejiang Province. J. Nat. Resour. 2013, 2, 220–233. [Google Scholar]
- Chen, J.; Chen, J.; Jo, P. A Simple Method for Reconstructing a High- Quality NDVI Time-Series Data Set Based on the Savitzky-Golay Filter A Simple Method for Reconstructing a High-Quality NDVI Time-Series Data Set Based on the Savitzky—Golay Filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Palacios-Orueta, A.; Huesca, M.; Whiting, M.L.; Litago, J.; Khanna, S.; Garcia, M.; Ustin, S.L. Derivation of Phenological Metrics by Function Fitting to Time-Series of Spectral Shape Indexes AS1 and AS2: Mapping Cotton Phenological Stages Using MODIS Time Series. Remote Sens. Environ. 2012, 126, 148–159. [Google Scholar] [CrossRef]
- Yoshifuji, N.; Igarashi, Y.; Tanaka, N.; Tanaka, K.; Sato, T.; Tantasirin, C.; Suzuki, M. Inter-Annual Variation in the Response of Leaf-out Onset to Soil Moisture Increase in a Teak Plantation in Northern Thailand. Int. J. Biometeorol. 2014, 58, 2025–2029. [Google Scholar] [CrossRef] [PubMed]
- Dannenberg, M.P.; Song, C.; Hwang, T.; Wise, E.K. Empirical Evidence of El Nino-Southern Oscillation Influence on Land Surface Phenology and Productivity in the Western United States. Remote Sens. Environ. 2015, 159, 167–180. [Google Scholar] [CrossRef]
- Potithep, S.; Nagai, S.; Nasahara, K.N.; Muraoka, H.; Suzuki, R. Two Separate Periods of the LAI-VIs Relationships Using in Situ Measurements in a Deciduous Broadleaf Forest. Agric. For. Meteorol. 2013, 169, 148–155. [Google Scholar] [CrossRef]
- Borchert, R. Water Status and Development of Tropical Trees During Seasonal Drought. Trees. 1994, 8, 115–124. [Google Scholar] [CrossRef]
- Elliott, S.; Baker, P.J.; Borchert, R. Leaf Flushing during the Dry Season: The Paradox of Asian Monsoon Forests. Glob. Ecol. Biogeogr. 2006, 15, 248–257. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
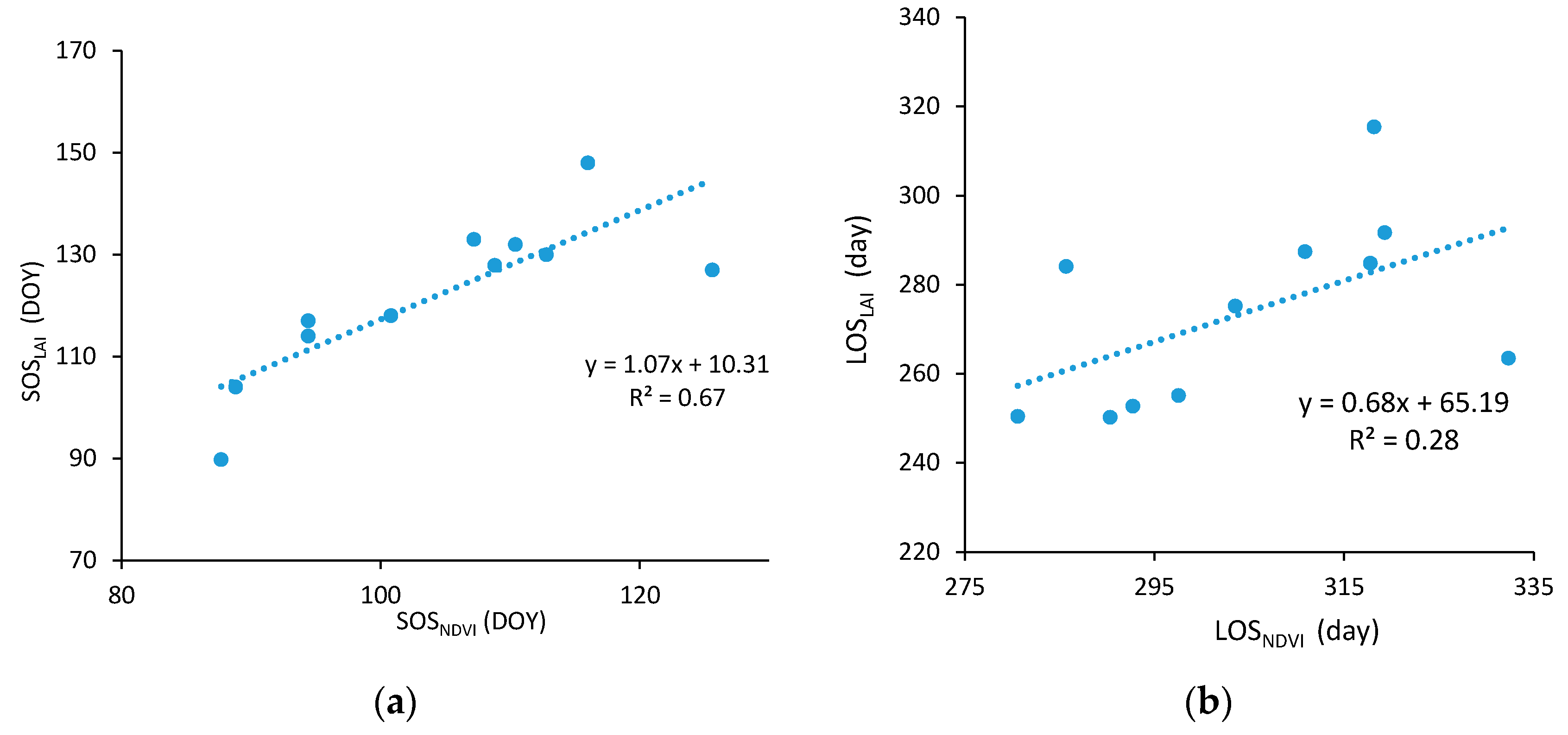{kind=link}
{kind=link}
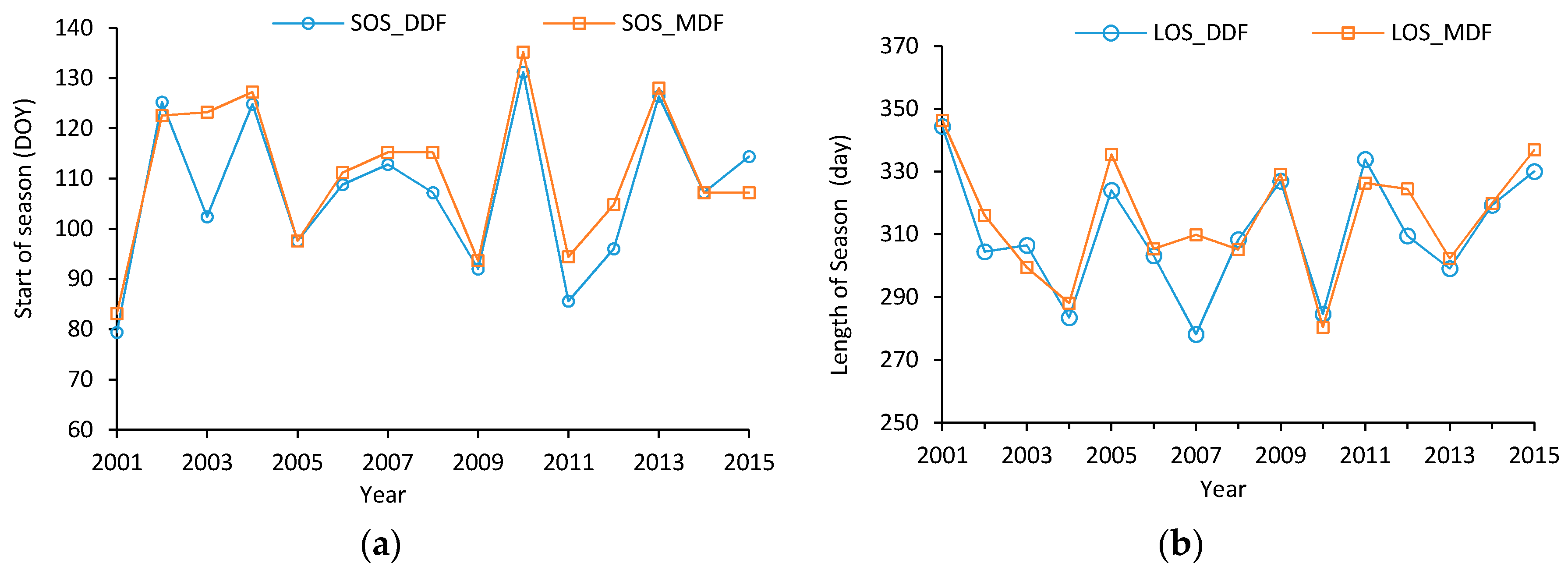{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Landsat Path/Row | Date Acquired |
|---|---|---|
| 1 | Landsat OLI-8 130/47 | 16-01-2017 |
| 2 | Landsat OLI-8 130/47 | 16-01-2017 |
| 3 | Landsat OLI-8 131/47 | 08-02-2017 |
| 4 | Landsat OLI-8 131/47 | 08-02-2017 |
| Neutral Year (Mean ± 1SD) | El Niño Year (2010) | La Niña Year (2011) | The Area Size Delayed by El Niño (2010) (±Ha, 1SD) | The Areasize Advanced by La Niña (2011) (±Ha, 1SD) | |
|---|---|---|---|---|---|
| (DOY) | 106.4 ± 7.1 | 131.8 | 86.5 | 168,337.5 (75.3%) | 147,718.8 (66.0%) |
| (DOY) | 106.9 ± 6.9 | 126.9 | 92.4 | 210,662.5 (65.8%) | 153,762.5 (48.0%) |
| (Day) | 306.9 ± 13.9 | 279.0 | 321.9 | 214,806.3 (96.0%) | 221,818.8 (99.2%) |
| (Day) | 319.0 ± 12.5 | 292.7 | 327.3 | 307,262.5 (96.0%) | 307,931.3 (96.2%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diem, P.K.; Pimple, U.; Sitthi, A.; Varnakovida, P.; Tanaka, K.; Pungkul, S.; Leadprathom, K.; LeClerc, M.Y.; Chidthaisong, A. Shifts in Growing Season of Tropical Deciduous Forests as Driven by El Niño and La Niña during 2001–2016. Forests 2018, 9, 448. https://doi.org/10.3390/f9080448
Diem PK, Pimple U, Sitthi A, Varnakovida P, Tanaka K, Pungkul S, Leadprathom K, LeClerc MY, Chidthaisong A. Shifts in Growing Season of Tropical Deciduous Forests as Driven by El Niño and La Niña during 2001–2016. Forests. 2018; 9(8):448. https://doi.org/10.3390/f9080448
Chicago/Turabian StyleDiem, Phan Kieu, Uday Pimple, Asamaporn Sitthi, Pariwate Varnakovida, Katsunori Tanaka, Sukan Pungkul, Kumron Leadprathom, Monique Y. LeClerc, and Amnat Chidthaisong. 2018. "Shifts in Growing Season of Tropical Deciduous Forests as Driven by El Niño and La Niña during 2001–2016" Forests 9, no. 8: 448. https://doi.org/10.3390/f9080448
APA StyleDiem, P. K., Pimple, U., Sitthi, A., Varnakovida, P., Tanaka, K., Pungkul, S., Leadprathom, K., LeClerc, M. Y., & Chidthaisong, A. (2018). Shifts in Growing Season of Tropical Deciduous Forests as Driven by El Niño and La Niña during 2001–2016. Forests, 9(8), 448. https://doi.org/10.3390/f9080448