Simulating Long-Term Effects of Bioenergy Extraction on Dead Wood Availability at a Landscape Scale in Sweden

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
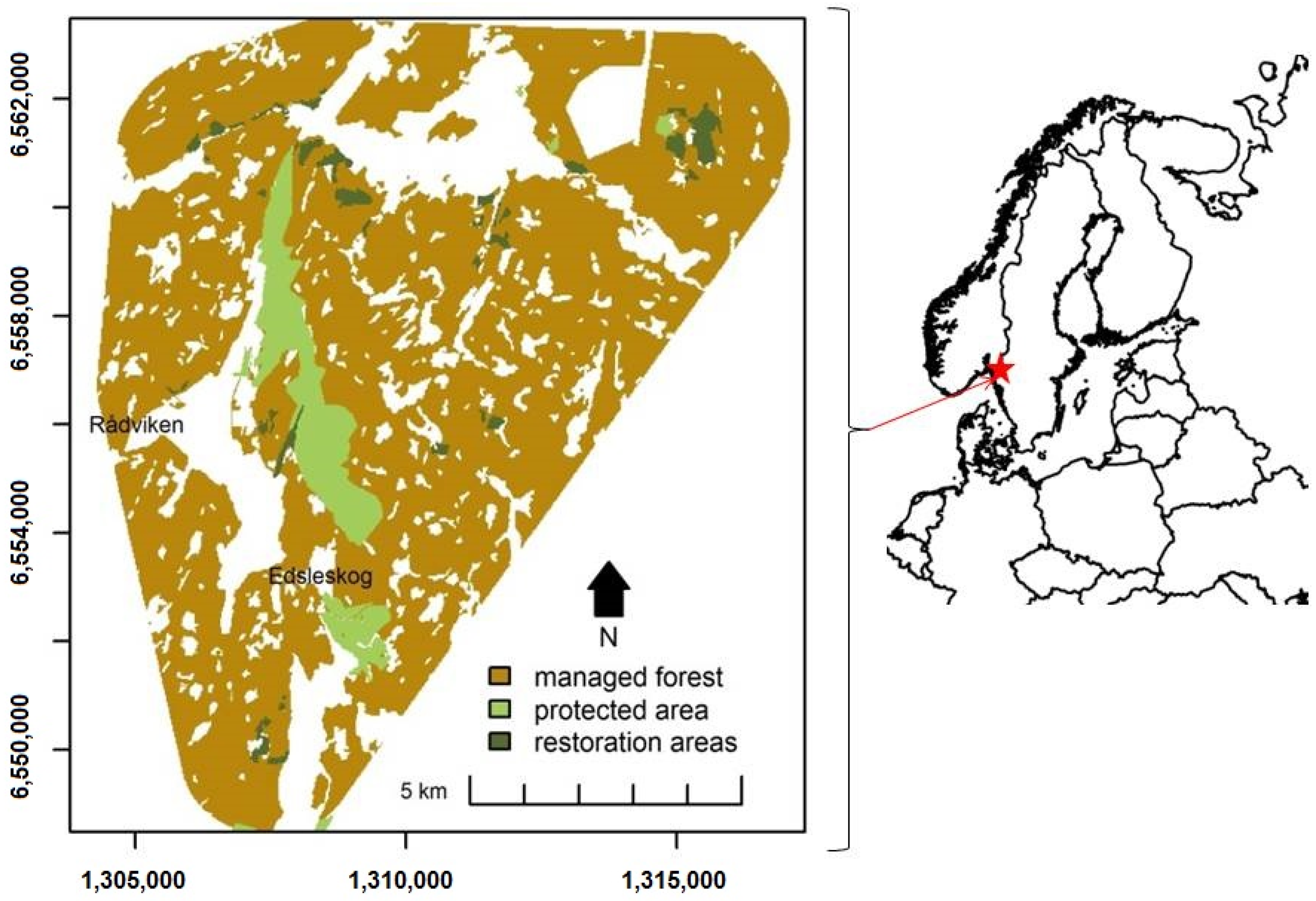
2.1. Study Site
2.2. Forest Dynamics Simulation
2.3. Bioenergy Extraction
2.4. Analyses
3. Results
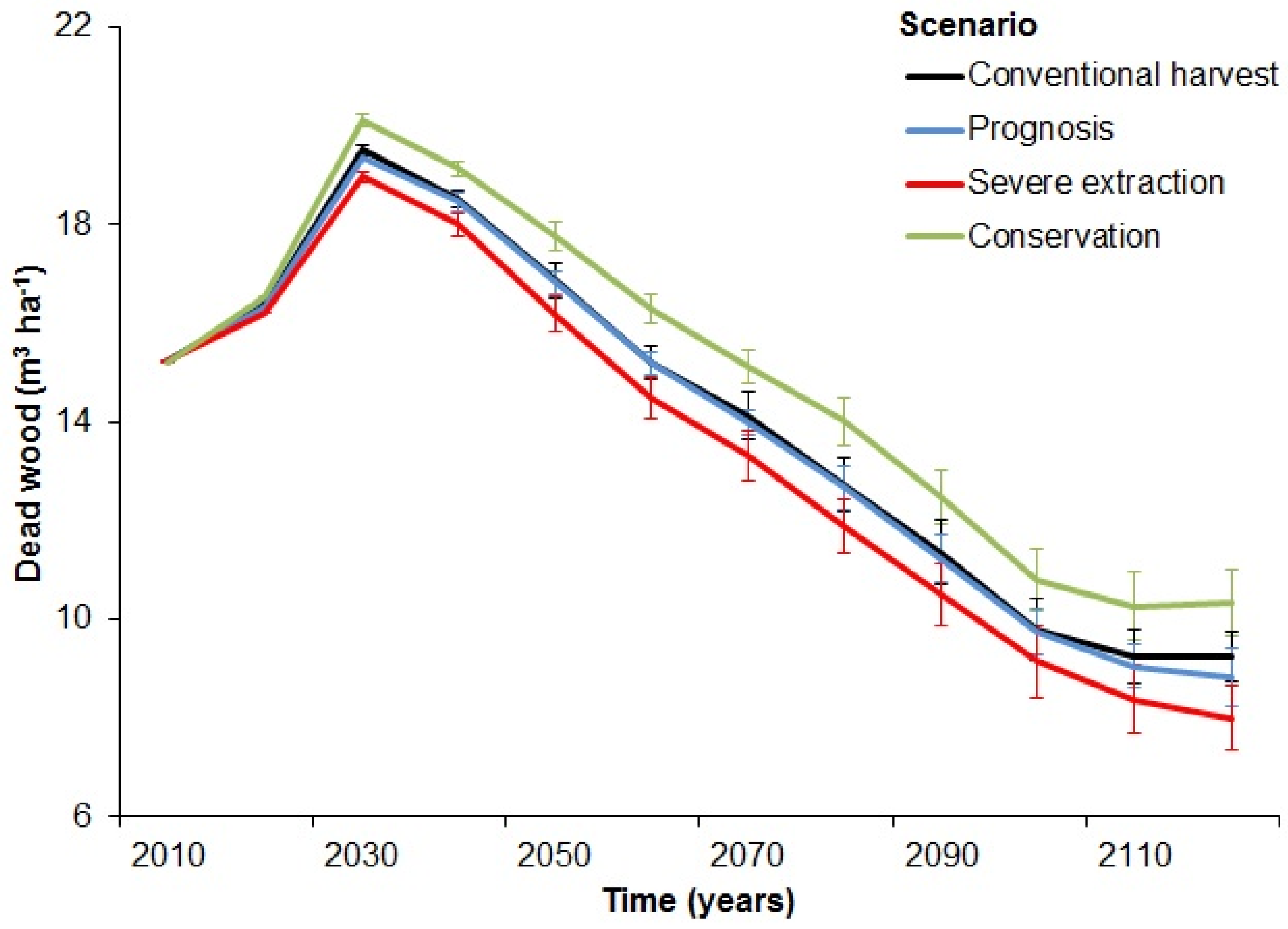
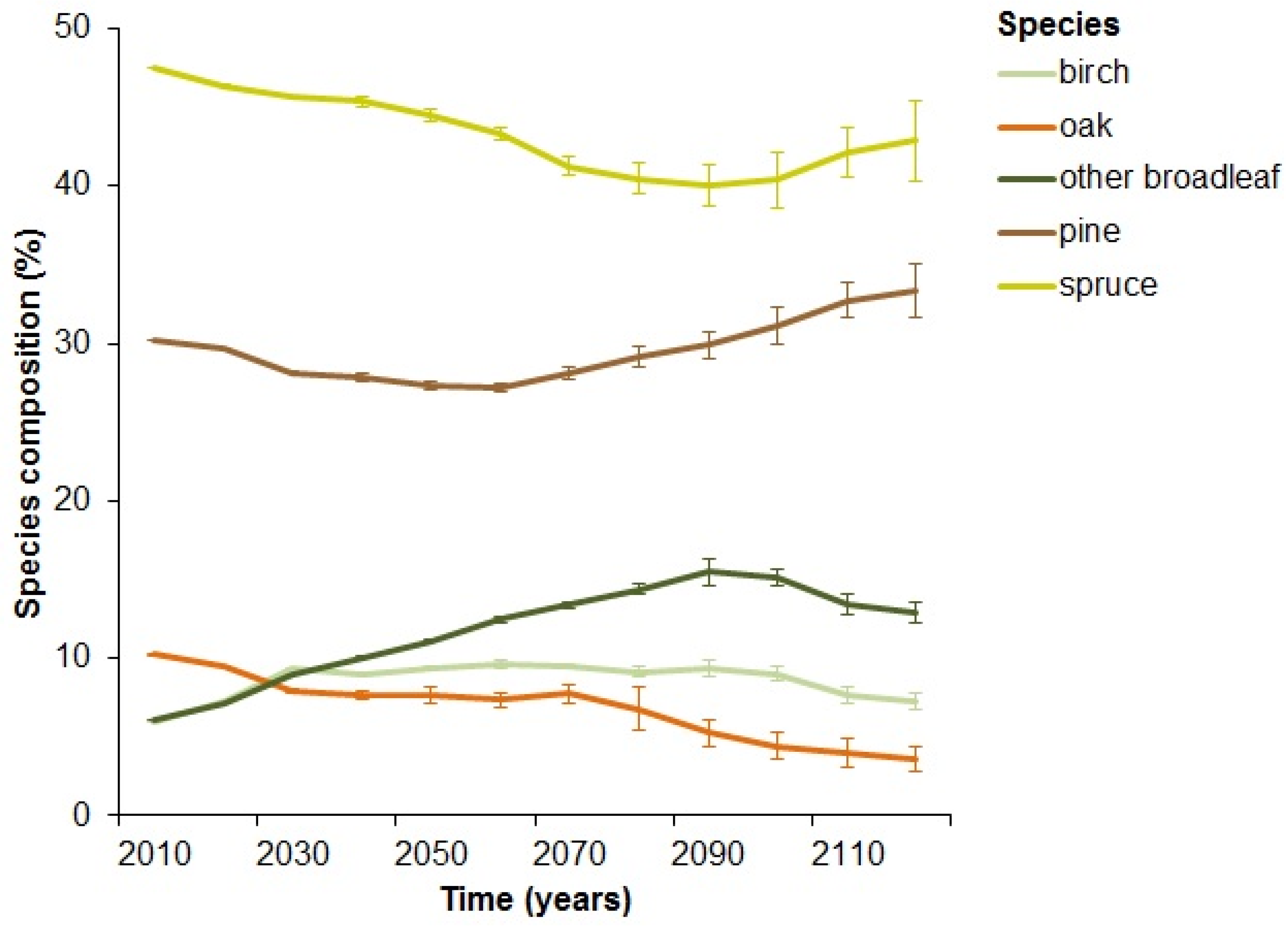
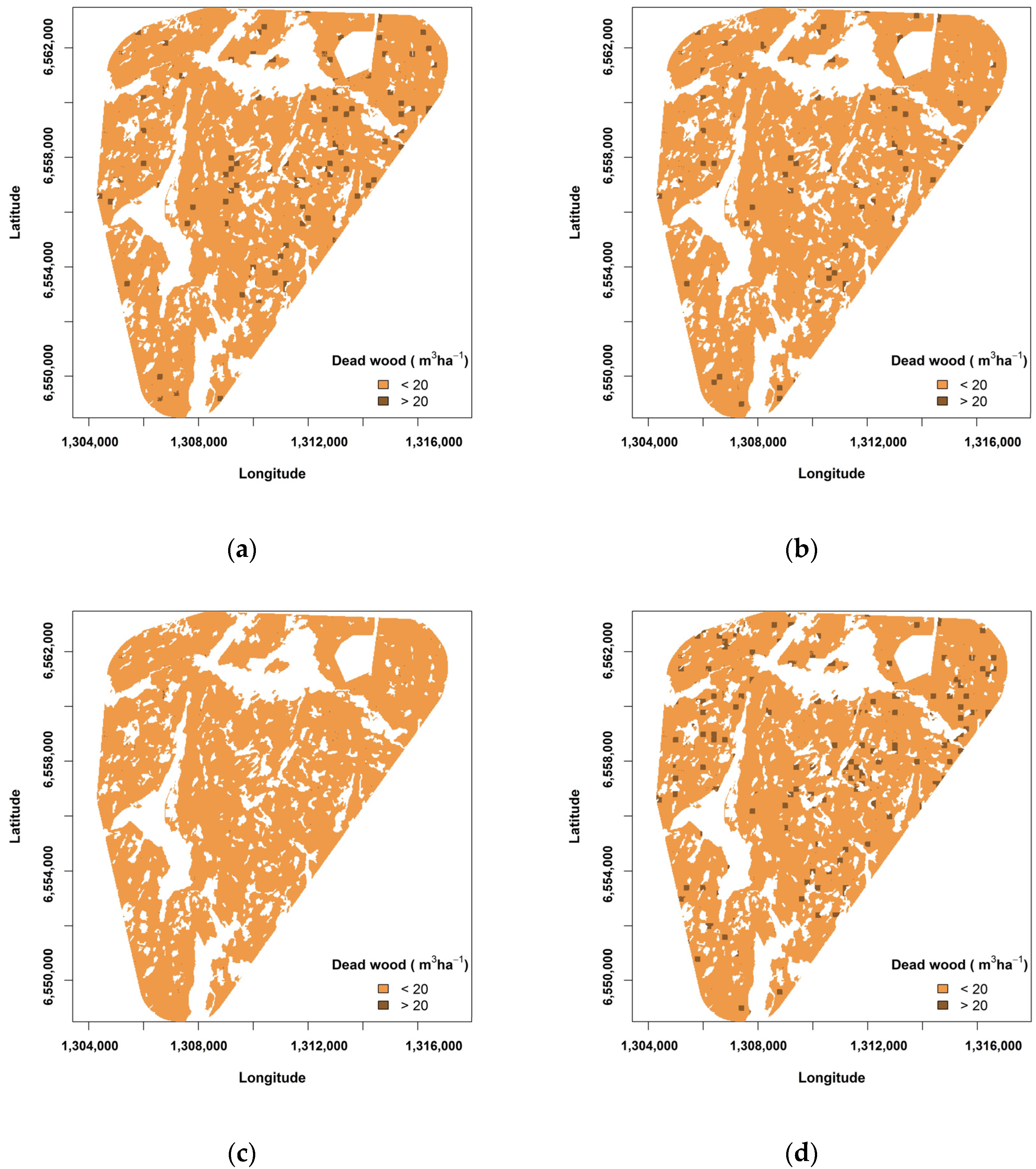
3.1. Levels of Dead Wood in the Landscape
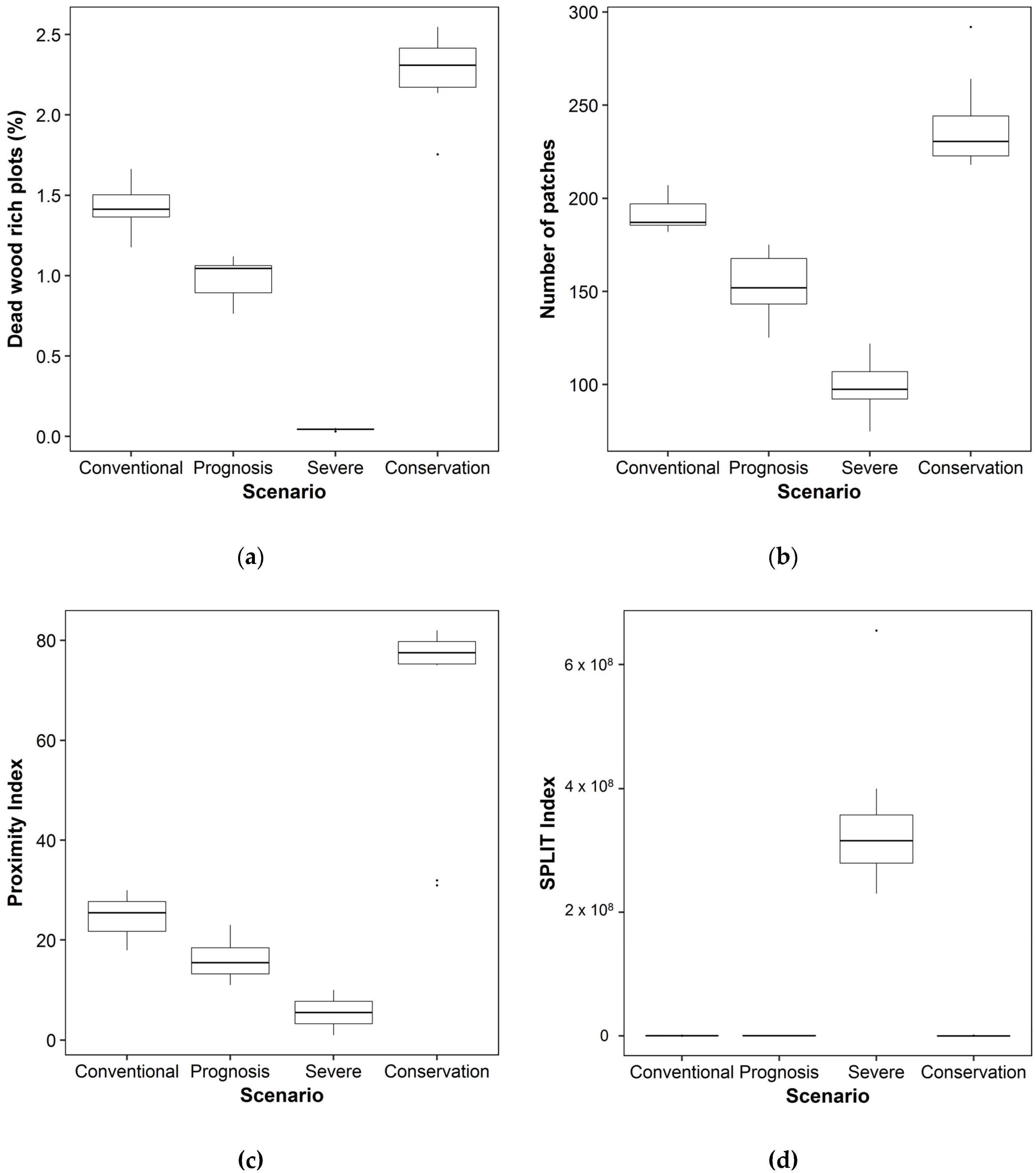
3.2. Consequences for Biodiversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Commission (2013). The 2020 Climate and Energy Package. European Commission. Available online: https://ec.europa.eu/clima/policies/strategies/2020_nl (accessed on 18 July 2018).
- Geijer, E.; Andersson, J.; Bostedt, G.; Brannlund, R.; Hjältèn, J. Safeguarding species richness vs. increasing the use of renewable energy-The effect of stump harvesting on two environmental goals. J. For. Econ. 2014, 20, 111–125. [Google Scholar] [CrossRef]
- Jonsell, M.; Schroeder, M. Proportions of saproxylic beetle populations that utilise clear-cut stumps in a boreal landscape-Biodiversity implications for stump harvest. For. Ecol. Manag. 2014, 334, 313–320. [Google Scholar] [CrossRef]
- Andersson, J.; Hjältèn, J.; Dynesius, M. Wood-inhabiting beetles in low stumps, high stumps and logs on boreal clear-cuts: Implications for dead wood management. PLoS ONE 2015, 10, e0118896. [Google Scholar] [CrossRef] [PubMed]
- Bouget, C.; Lassauce, A.; Jonsell, M. Effects of fuelwood harvesting on biodiversity—A review focused on the situation in Europe. Can. J. For. Res. 2012, 42, 1421–1432. [Google Scholar] [CrossRef]
- Work, T.T.; Andersson, J.; Ranius, T.; Hjältén, J. Defining stump harvesting retention targets required to maintain saproxylic beetle biodiversity. For. Ecol. Manag. 2016, 371, 90–102. [Google Scholar] [CrossRef]
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forest as an example. Ecol. Bull. 2001, 49, 11–41. [Google Scholar]
- Grove, S.J. Saproxylic insect ecology and the sustainable management of forests. Annu. Rev. Ecol. Syst. 2002, 33, 1–23. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012; pp. 1–509. ISBN 9781139025843. [Google Scholar]
- Esseen, P.-A.; Ehnström, B.; Ericson, L.; Sjöberg, K. Boreal forests. Ecol. Bull. 1997, 46, 16–47. [Google Scholar]
- Kuuluvainen, T.; Aakala, T. Natural forest dynamics in boreal Fennoscandia: A review and classification. Silva Fenn. 2011, 45, 823–841. [Google Scholar] [CrossRef]
- Ranius, T.; Fahrig, L. Targets for maintenance of dead wood for biodiversity conservation based on extinction thresholds. Scand. J. For. Res. 2006, 21, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Bütler, R. A review of habitat thresholds for dead wood, A baseline for management recommendations in European forests. Eur. J. For. Res. 2010, 129, 981–992. [Google Scholar] [CrossRef]
- Work, T.T.; Hibbert, A. Estimating species loss of saproxylic flies under scenarios of reduced coarse woody material CWM, in eastern boreal mixedwood forests. Ecosphere 2011, 2, 1–11. [Google Scholar] [CrossRef]
- Fridman, J.; Walheim, M. Amount, structure, and dynamics of dead wood on managed forestland in Sweden. For. Ecol. Manag. 2000, 131, 23–36. [Google Scholar] [CrossRef]
- Anonymous. Skogsdata 2016; Institutionen för skoglig resurshushållning SLU: Umeå, Sweden, 2016; pp. 1–146. [Google Scholar]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Ódor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests, meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Penttila, R.; Siitonen, J.; Kuusinen, M. Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol. Conserv. 2004, 117, 271–283. [Google Scholar] [CrossRef]
- Kruys, N.; Jonsson, B.G. Fine woody debris is important for species richness on logs in managed boreal spruce forests of northern Sweden. Can. J. For. Res. 1999, 29, 1295–1299. [Google Scholar] [CrossRef]
- Hjältén, J.; Stenbacka, F.; Andersson, J. Saproxylic beetle assemblages on low stumps, high tumps and logs: Implications for environmental effects of stump harvesting. For. Ecol. Manag. 2010, 26, 1149–1155. [Google Scholar] [CrossRef]
- Persson, T.; Egnell, G. Stump harvesting for bioenergy: A review of climatic and environmental impacts in northern Europe and America. WIRES Energy Environ. 2018, e307. [Google Scholar] [CrossRef]
- Kommissionen-mot-oljeberoende. På väg Mot ett Oljefritt Sverige; Statsrådsberedningen: Stockholm, Sweden. 2006; pp. 1–45. [Google Scholar]
- Hof, A.R.; Hjältén, J. Are we restoring enough? Simulating impacts of restoration efforts on the suitability of forest landscapes for a locally critically endangered umbrella species. Restor. Ecol. 2017. [Google Scholar] [CrossRef]
- Scheller, R.M.; Domingo, J.B.; Sturtevant, B.R.; Williams, J.S.; Rudy, A.; Gustafson, E.J.; Mladenoff, D.J. Design, development, and application of LANDIS-II: A spatial landscape simulation model with flexible temporal and spatial resolution. Ecol. Model. 2007, 201, 409–419. [Google Scholar] [CrossRef]
- Scheller, R.M.; Mladenoff, D.J. A forest growth and biomass module for a landscape simulation model, LANDIS: Design, validation, and application. Ecol. Model. 2004, 180, 211–229. [Google Scholar] [CrossRef]
- Pennanen, J.; Kuuluvainen, T. A spatial simulation approach to natural forest landscape dynamics in boreal Fennoscandia. For. Ecol. Manag. 2002, 164, 157–175. [Google Scholar] [CrossRef]
- Scheller, R.M.; Mladenoff, D.J. A spatially interactive simulation of climate change, harvesting, wind, and tree species migration and projected changes to forest composition and biomass in northern Wisconsin, USA. Glob. Chang. Biol. 2005, 11, 307–321. [Google Scholar] [CrossRef]
- Dymond, C.C.; Beukema, S.; Scheller, R. LANDIS-II Forest Carbon Succession Extension v2.0 User Guide. Self-published. 2015. Available online: www.landis-ii.org (accessed on 23 March 2016).
- Dymond, C.C.; Beukema, S.; Nitschke, C.R.; Coates, K.D.; Scheller, R.M. Carbon sequestration in managed temperate coniferous forests under climate change. Biogeosciences 2016, 13, 1933–1947. [Google Scholar] [CrossRef]
- Scheller, R.M.; Domingo, J.B. LANDIS-II Model v6 0–User Guide; Portland State University: Portland, OR, USA, 2011; pp. 1–21. [Google Scholar]
- Scheller, R.M.; Sturtevant, B.R.; Gustafson, E.J.; Miranda, B.R.; Zollner, P.A.; Mladenoff, D.J.; Domingo, J.B. Base Harvest v2.2 LANDIS-II Extension User Guide; Portland State University: Portland, OR, USA, 2015; pp. 1–28. [Google Scholar]
- Bergeron, Y.; Flannigan, M.; Gauthier, S.; Leduc, A.; Lefort, P. Past, current and future fire frequency in the Canadian boreal forest, Implications for sustainable forest management. Ambio 2004, 33, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Zackrisson, O. Influence of forest fires on the North Swedish boreal forest. Oikos 1977, 29, 22–32. [Google Scholar] [CrossRef]
- Östlund, L.; Zackrisson, O.; Axelsson, A.L. The history and transformation of a Scandinavian boreal forest landscape since the 19th century. Can. J. For. Res. 1997, 27, 1198–1206. [Google Scholar] [CrossRef]
- Linder, P.; Östlund, L. Structural changes in three mid-boreal Swedish forest landscapes, 1885–1996. Biol. Conserv. 1998, 851, 9–19. [Google Scholar] [CrossRef]
- Reese, H.; Nilsson, M.; Pahen, T.G.; Hagner, O.; Joyce, S.; Tingelof, U.; Egberth, M.; Olsson, H. Countrywide estimates of forest variables using satellite data and field data from the national forest inventory. Ambio 2003, 32, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Korzukhin, M.; Rubinia, A.; Bonan, G.; Solomon, A.; Antonovsky, M. The Silvics of Some East European and Siberian Boreal Forest Tree Species; International Institute for Applied Systems Analysis: Laxenburg, Austria, 1989; pp. 1–27. [Google Scholar]
- Perala, D.A.; Alm, A.A. Reproductive ecology of birch: A review. For. Ecol. Manag. 1990, 32, 1–38. [Google Scholar] [CrossRef]
- Prentice, I.C.; Helmisaari, H. Silvics of north European trees, compilation, comparisons and implications for forest succession modelling. For. Ecol. Manag. 1991, 42, 79–93. [Google Scholar] [CrossRef]
- Hofgaard, A. Structure and regeneration patterns in a virgin Picea abies forest in northern Sweden. J. Veg. Sci. 1993, 4, 601–608. [Google Scholar] [CrossRef]
- Reyes, O.; Casal, M.; Trabaud, L. The influence of population; fire and time of dissemination on the germination of Betula pendula seeds. Plant Ecol. 1997, 133, 201–208. [Google Scholar] [CrossRef]
- Welander, N.; Ottosson, B. The influence of shading on growth and morphology in seedlings of Quercus robur L. and Fagus sylvatica L. For. Ecol. Manag. 1998, 107, 117–126. [Google Scholar] [CrossRef]
- Næsset, E. Decomposition rate constants of Picea abies logs in southeastern Norway. Can. J. For. Res. 1999, 293, 372–381. [Google Scholar] [CrossRef]
- Berg, B.; Johansson, M.B.; Meentemeyer, V. Litter decomposition in a transect of Norway spruce forests, substrate quality and climate control. Can. J. For. Res. 2000, 307, 1136–1147. [Google Scholar] [CrossRef]
- Tinner, W.; Conedera, M.; Gobet, E.; Hubschmid, P.; Wehrli, M.; Ammann, B. A palaeoecological attempt to classify fire sensitivity of trees in the southern Alps. The Holocene 2000, 10, 565–574. [Google Scholar] [CrossRef]
- Deiller, A.F.; Walter, J.M.N.; Trémoličres, M. Regeneration strategies in a temperate hardwood floodplain forest of the Upper Rhine, sexual versus vegetative reproduction of woody species. For. Ecol. Manag. 2003, 180, 215–225. [Google Scholar] [CrossRef]
- Pensa, M.; Jalkanan, R. Variation in needle longevity is related to needle-fascicle production rate in Pinus sylvestris. Tree Physiol. 2005, 25, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, H.; Hynynen, J.; Siitonen, J.; Sievänen, R. Predicting the decomposition of Scots pine; Norway spruce; and birch stems in Finland. Ecol. Appl. 2006, 16, 1865–1879. [Google Scholar] [CrossRef]
- Withington, J.M.; Reich, P.B.; Oleksyn, J.; Eissenstat, D.M. Comparisons of structure and life span in roots and leaves among temperate trees. Ecol. Monogr. 2006, 763, 381–397. [Google Scholar] [CrossRef]
- Chalupka, W. Reproduction. In Biology and Ecology of Norway Spruce; Tjoelker, M.G., Boratynski, A., Bugala, W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 97–114. ISBN 978-1-4020-4841-8. [Google Scholar]
- Modrzynski, J. Outline of Ecology. In Biology and Ecology of Norway Spruce; Tjoelker, M.G., Boratynski, A., Bugala, W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 195–253. ISBN 978-1-4020-4841-8. [Google Scholar]
- Proença, V.A.M. Galicio-Portuguese Oak Forest of Quercus robur and Quercus pyrenaica, Biodiversity Patterns and Forest Response to Fire. Ph.D. Dissertation, Universidade de Lisboa, Lisbon, Portugal, 2009; p. 172. [Google Scholar]
- Kurz, W.A.; Dymond, C.C.; White, T.M.; Stinson, G.; Shaw, C.H.; Rampley, G.J.; Smyth, C.; Simpson, B.N.; Neilson, E.T.; Trofymow, J.A.; et al. CBM-CFS3, a model of carbon-dynamics in forestry and land-use change implementing IPCC standards. Ecol. Model. 2009, 2204, 480–504. [Google Scholar] [CrossRef]
- Conedera, M.; Lucini, L.; Valese, E.; Ascoli, D.; Pezzati, G.B. Fire resistance and vegetative recruitment ability of different deciduous trees species after low-to moderate-intensity surface fires in southern Switzerland. In Proceedings of the VI International Conference on Forest Fire Research, Coimbra, Portugal, 15–18 November 2010; Viegas, D.X., Ed.; University of Coimbra: Coimbra, Portugal, 2010; pp. 15–18. [Google Scholar]
- Hansson, K.; Helmisaari, H.S.; Sah, S.P.; Lange, H. Fine root production and turnover of tree and understorey vegetation in Scots pine; silver birch and Norway spruce stands in SW Sweden. For. Ecol. Manag. 2013, 309, 58–65. [Google Scholar] [CrossRef]
- Anonymous. Skogsvårds Lagstiftningen, Gällande Regler 1 April 2017; Skogsstyrelsen: Jönköping, Sweden, 2017; pp. 1–92. ISBN 978-91-87535-06-2. [Google Scholar]
- Nilsson, P.; Cory, N. Forestry Statistics 2013; Swedish University of Agricultural Sciences: Umeå, Sweden, 2013; pp. 1–158. [Google Scholar]
- Sandström, F.; Petersson, H.; Kruys, N.; Ståhl, G. Biomass conversion factors density and carbon concentration, by decay classes for dead wood of Pinus sylvestris, Picea abies and Betula spp. in boreal forests of Sweden. For. Ecol. Manag. 2007, 243, 19–27. [Google Scholar] [CrossRef]
- Krankina, O.N.; Harmon, M.E. Dynamics of the dead wood carbon pool in northwestern Russian boreal forests. Water Air Soil Pollut. 1995, 82, 227–238. [Google Scholar] [CrossRef]
- Sollins, P.; Cline, S.P.; Verhoeven, T.; Sachs, D.; Spycher, G. Patterns of log decay in old-growth Douglas-fir forests. Can. J. For. Res. 1987, 17, 1585–1595. [Google Scholar] [CrossRef]
- Groombridge, B. Global Biodiversity, Status of the Earth’s Living Resources; Chapman and Hall: New York, NY, USA, 1992; pp. 1–585. ISBN 978-94-011-2282-5. [Google Scholar]
- Gustafson, E.J.; Parker, G.R. Relationships between landcover proportion and indices of landscape spatial pattern. Landsc. Ecol. 1992, 7, 101–110. [Google Scholar] [CrossRef]
- McGarial, K.; Marks, B. FRAGSTAT, Spatial Pattern Analysis Program for Quantifying Landscape Structure; United States Department of Agriculture Pacific Northwest Research Station: Portland, OR, USA, 1995; p. 122. [Google Scholar] [CrossRef]
- McGarical, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4, Spatial Pattern Analysis Program for Categorical and Continuous Maps; University of Massachusetts: Amherst, MA, USA, 2012; Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 18 March 2018).
- The R Foundation for Statistical Computing Platform. R version 3.2.2. 2015. Available online: http://www.r-project.org/ (accessed on 6 December 2015).
- Hijmans, R.J.; van Etten, J.; Cheng, J.; Mattiuzzi, M.; Sumner, M.; Greenberg, J.A.; Perpinan Lamigueiro, O.; Bevan, A.; Racine, E.B.; Ashton Shortridge, A.; et al. Raster: Geographic Data Analysis and Modeling; R package version 2.6-7. 2017. Available online: https://cran.r-project.org/web/packages/raster/ (accessed on 27 July 2018).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Lindenmayer, D.B.; Franklin, J.F.; Lõhmus, A.; Baker, S.C.; Bauhus, J.; Beese, W.; Brodie, A.; Kiehl, B.; Kouki, J.; Martínez Pastur, G.; et al. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012, 5, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, B.G.; Ekström, M.; Esseen, P.A.; Grafström, A.; Ståhl, G.; Westerlund, B. Dead wood availability in managed Swedish forests–Policy outcomes and implications for biodiversity. For. Ecol. Manag. 2016, 376, 174–182. [Google Scholar] [CrossRef]
- Aakala, T.; Kuuluvainen, T.; Wallenius, T.; Kauhanen, H. Contrasting patterns of tree mortality in late-successional Picea abies stands in two areas in northern Fennoscandia. J. Veg. Sci. 2009, 20, 1016–1026. [Google Scholar] [CrossRef]
- Stenbacka, F.; Hjältén, J.; Hilszczański, J.; Dynesius, M. Saproxylic and non-saproxylic beetle assemblages in boreal spruce forests of different age and forestry intensity. Ecol. Appl. 2010, 20, 2310–2321. [Google Scholar] [CrossRef] [PubMed]
- Johansson, V.; Felton, A.; Ranius, T. Long-term landscape scale effects of bioenergy extraction on dead wood-dependent species. For. Ecol. Manag. 2016, 371, 103–113. [Google Scholar] [CrossRef]
- Green, P.; Peterken, G.F. Variation in the amount of dead wood in the woodlands of the Lower Wye Valley, UK in relation to the intensity of management. For. Ecol. Manag. 2017, 98, 229–238. [Google Scholar] [CrossRef]
- Toivanen, T.; Kotiaho, J.S. The preferences of saproxylic beetle species for different dead wood types created in forest restoration treatments. Can. J. For. Res. 2010, 40, 445–464. [Google Scholar] [CrossRef]
- Hägglund, R.; Hjältén, J. Substrate specific restoration promotes saproxylic beetle diversity in boreal forest set-asides. For. Ecol. Manag. 2018, 425, 45–58. [Google Scholar] [CrossRef]
- Schmitz, H.; Bousack, H. Modelling a historic oil-tank fire allows an estimation of the sensitivity of the infrared receptors in pyrophilous Melanophila beetles. PLoS ONE 2012, 7, e37627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, J.; Isacsson, G. A comparison of the saproxylic beetle fauna between lowland and upland beech forests in southern Sweden. Ecol. Bull. 2010, 53, 131–139. [Google Scholar]
- Ranius, T. Measuring the dispersal of saproxylic insects, a key characteristic for their conservation. Popul. Ecol. 2006, 48, 177–188. [Google Scholar] [CrossRef]
- Schiegg, K. Effects of dead wood volume and connectivity on saproxylic insect species diversity. Ecoscience 2000, 7, 290–298. [Google Scholar] [CrossRef]
- Hanski, I. Insect conservation in boreal forests. J. Insect Conserv. 2008, 12, 451–454. [Google Scholar] [CrossRef]
- Hof, A.R.; Svahlin, A. The potential effect of climate change on the geographical distribution of insect pest species in the Swedish boreal forest. Scand. J. For. Res. 2016, 31, 29–39. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Müller, J.; Bußler, H.; Goßner, M.; Rettelbach, T.; Duelli, P. The European spruce bark beetle Ips typographus in a national park: from pest to keystone species. Biodivers. Conserv. 2008, 17, 2979. [Google Scholar] [CrossRef]
- Martikainen, P.; Siitonen, J.; Kaila, L.; Punttila, P.; Rauh, J. Bark beetles (Coleoptera, Scolytidae) and associated beetle species in mature managed and old-growth boreal forests in southern Finland. For. Ecol. Manag. 1999, 116, 233–245. [Google Scholar] [CrossRef]
- Hjältén, J.; Hägglund, R.; Johansson, T.; Roberge, J.-M.; Dynesius, M.; Olsson, J. Forest restoration by burning and gap cutting yield distinct immediate effects on saproxylic beetles. Biol. Conserv. 2017, 26, 1623–1640. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | TL | LL | SM | ST | ED | MD | VR | MIN | MAX |
|---|---|---|---|---|---|---|---|---|---|
| Picea abies (L.) Karsten | 500 | 5 | 50 | 3 | 50 | 400 | 0.2 | 0 | 500 |
| Pinus sylvestris L. | 500 | 3 | 10 | 1 | 50 | 1000 | 0 | 0 | 0 |
| Betula pendula Roth. | 250 | 1 | 10 | 2 | 50 | 200 | 0.4 | 0 | 200 |
| Quercus robur L. | 1000 | 1 | 40 | 3 | 20 | 3000 | 0.2 | 0 | 500 |
| Other broadleaf | 250 | 1 | 10 | 3 | 40 | 1000 | 0.2 | 0 | 250 |
| Biomass Pools | Conventional Harvest | Prognosis | Severe Extraction | Conservation |
|---|---|---|---|---|
| Live stemwood of merchantable size | 95% | 95% | 95% | 80% |
| Branches and stumps of newly harvested trees, small trees | 0% | 30% | 100% | 0% |
| Foliage | 0% | 0% | 100% | 0% |
| Coarse roots | 0% | 10% | 10% | 0% |
| Fine roots | 0% | 0% | 0% | 0% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hof, A.R.; Löfroth, T.; Rudolphi, J.; Work, T.; Hjältén, J. Simulating Long-Term Effects of Bioenergy Extraction on Dead Wood Availability at a Landscape Scale in Sweden. Forests 2018, 9, 457. https://doi.org/10.3390/f9080457
Hof AR, Löfroth T, Rudolphi J, Work T, Hjältén J. Simulating Long-Term Effects of Bioenergy Extraction on Dead Wood Availability at a Landscape Scale in Sweden. Forests. 2018; 9(8):457. https://doi.org/10.3390/f9080457
Chicago/Turabian StyleHof, Anouschka R., Therese Löfroth, Jörgen Rudolphi, Timothy Work, and Joakim Hjältén. 2018. "Simulating Long-Term Effects of Bioenergy Extraction on Dead Wood Availability at a Landscape Scale in Sweden" Forests 9, no. 8: 457. https://doi.org/10.3390/f9080457
APA StyleHof, A. R., Löfroth, T., Rudolphi, J., Work, T., & Hjältén, J. (2018). Simulating Long-Term Effects of Bioenergy Extraction on Dead Wood Availability at a Landscape Scale in Sweden. Forests, 9(8), 457. https://doi.org/10.3390/f9080457