Five Decades of Structural and Compositional Changes in Managed and Unmanaged Montane Stands: A Case Study from South-East Europe


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Management Regimes
2.2. Data Collection
2.3. Data Analysis
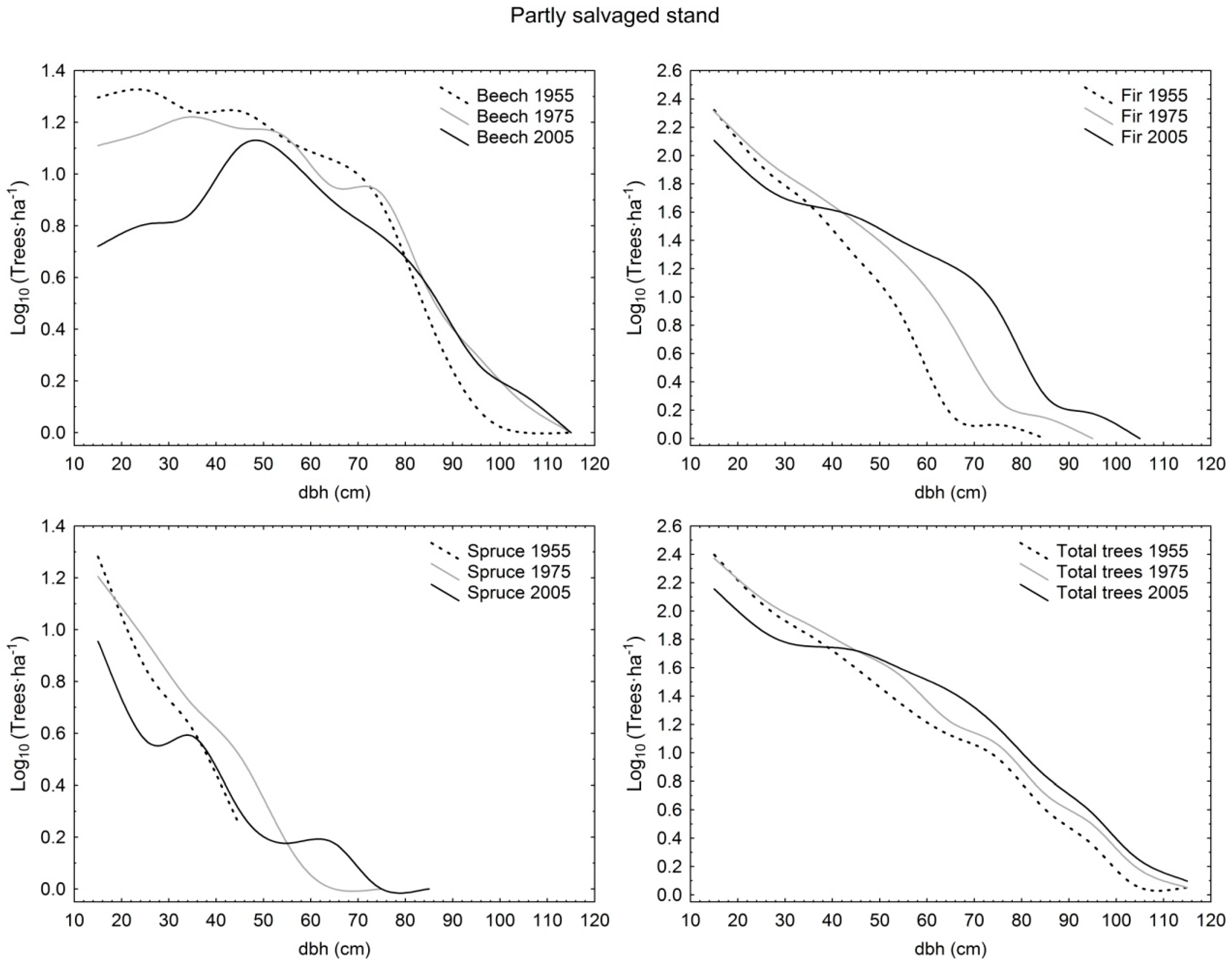
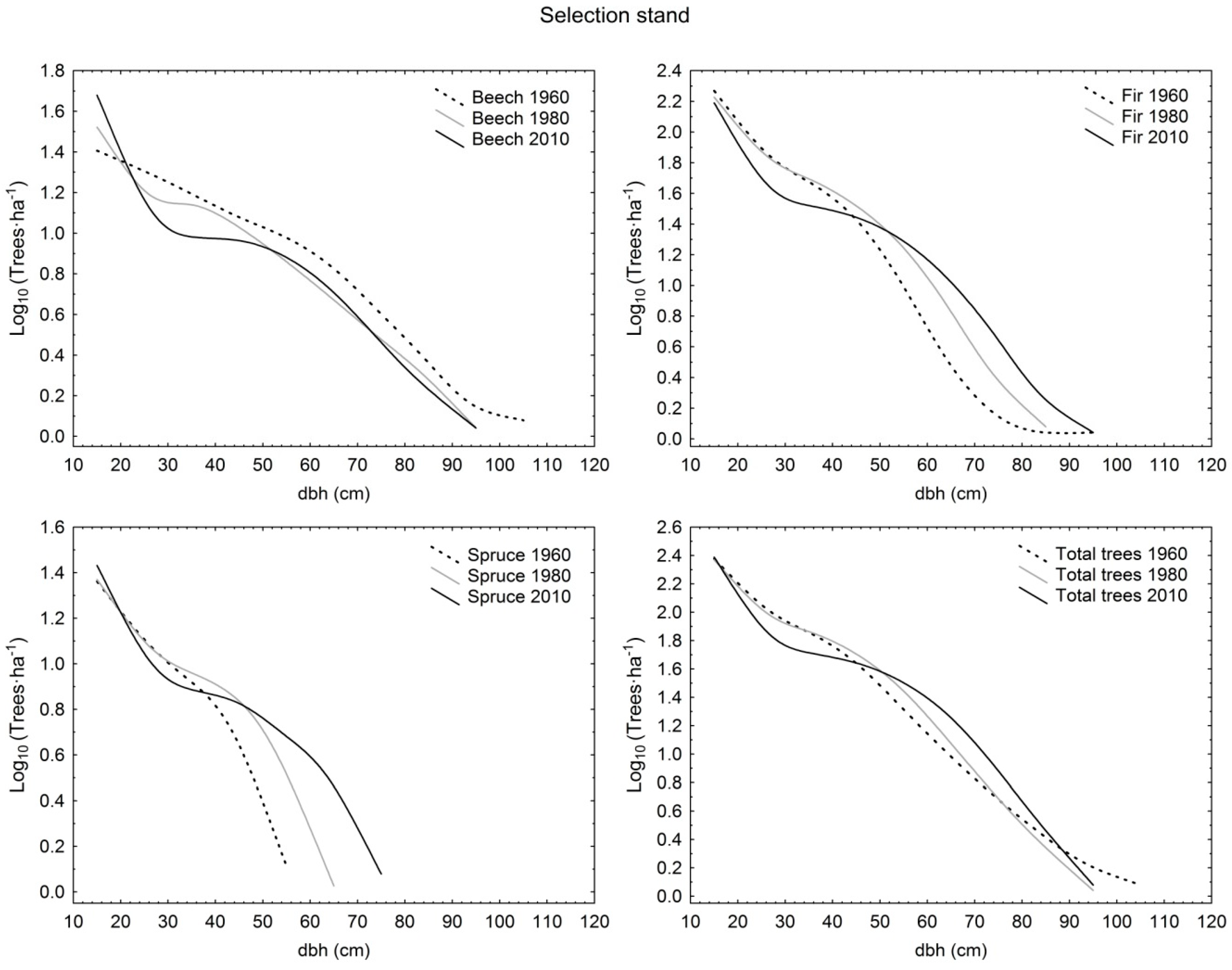
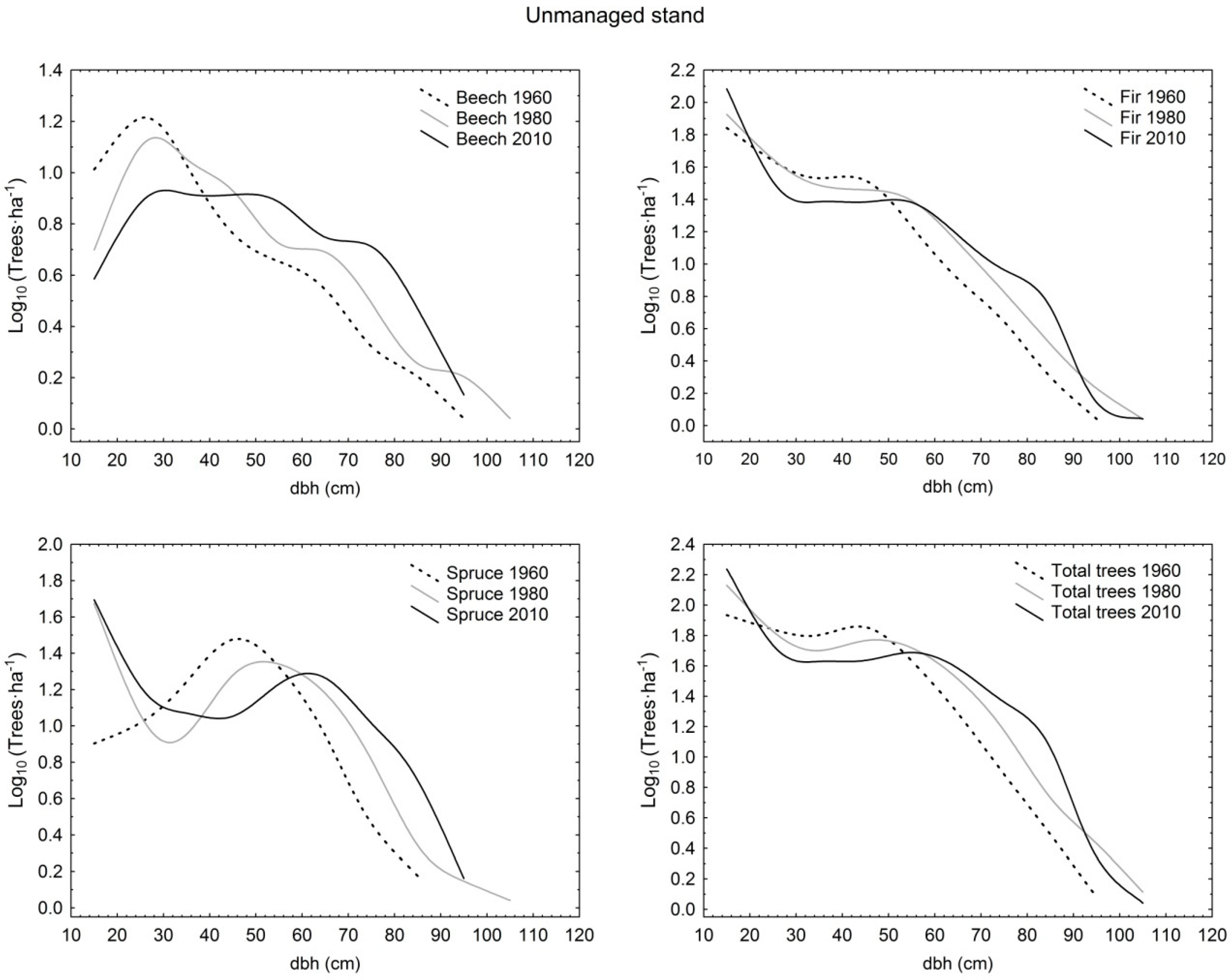
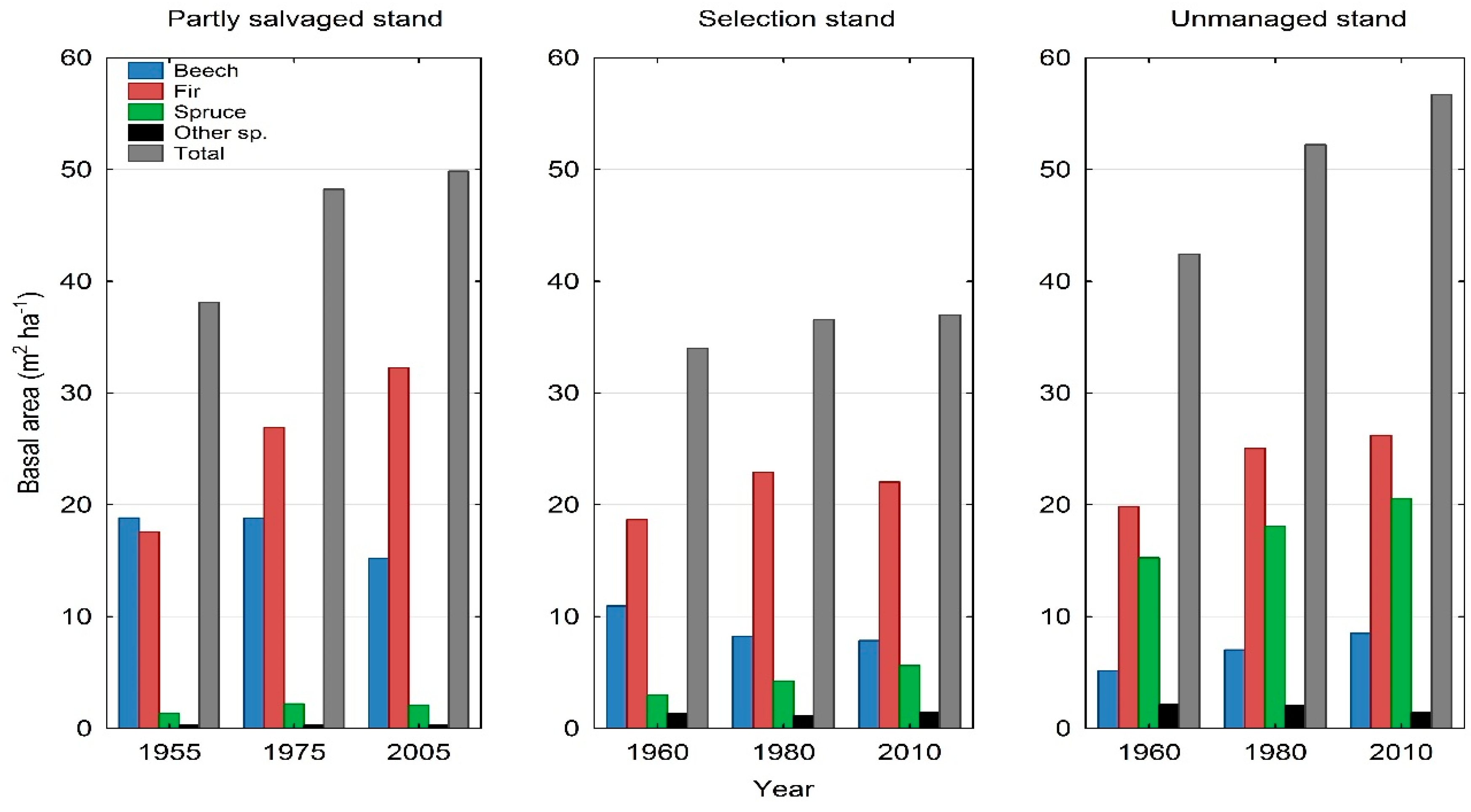
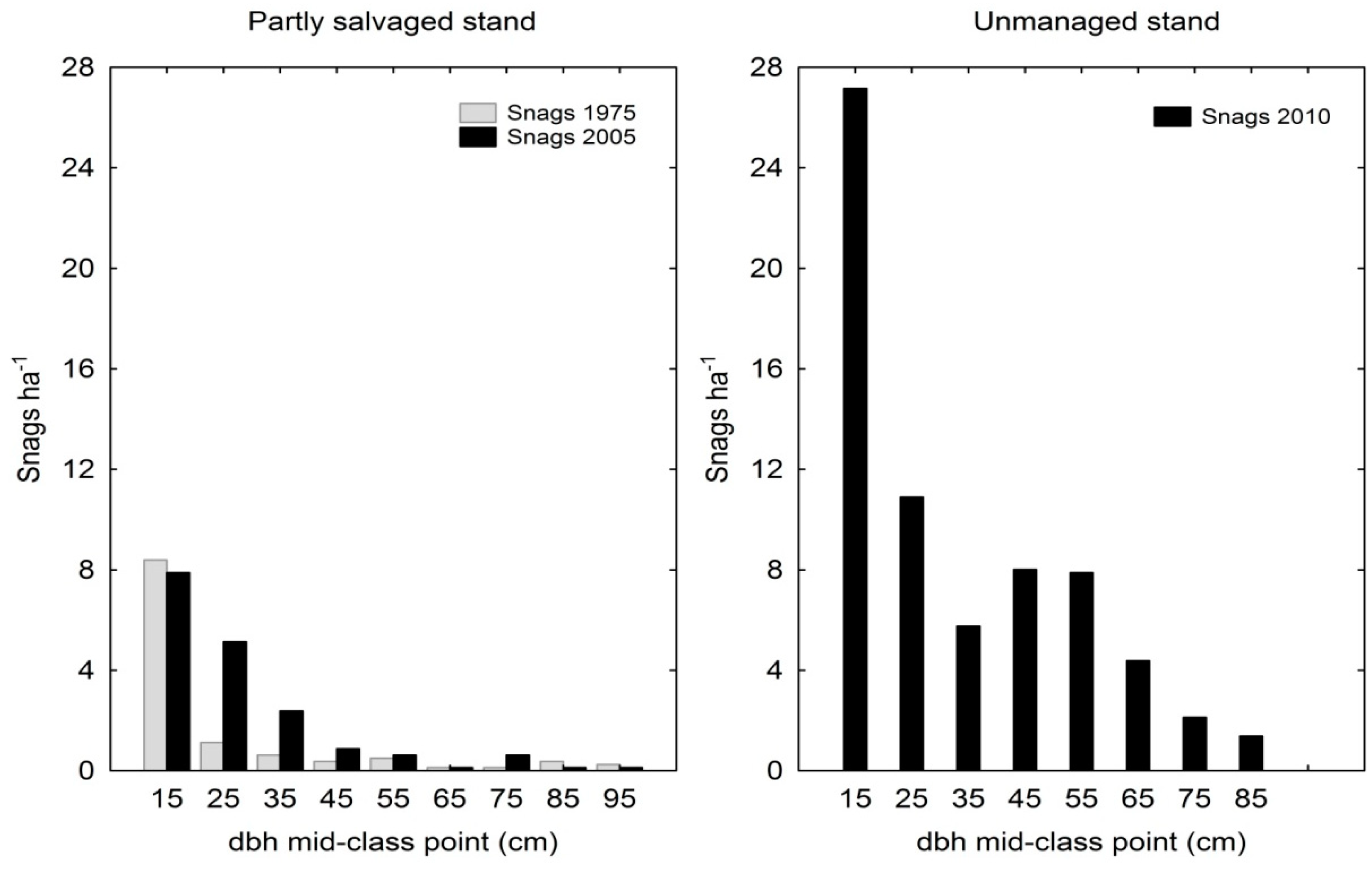
3. Results
3.1. Structural Characteristics of Live Trees
3.2. Tree Species Composition
3.3. Standing Dead Trees
4. Discussion
4.1. Structure of Living Trees
4.2. The Influence of Management Regimes on Tree Species Composition
4.3. Standing Dead Trees in Partly Salvaged Stand and Unmanaged Stand in Tara
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Partly Salvaged Stand | |||
| Inventory year | 1955 | 1975 | 2005 |
| Total number of measured trees | 4104 | 4440 | 3296 |
| Selection Stand | |||
| Inventory year | 1960 | 1980 | 2010 |
| Total number of measured trees | 5567 | 5622 | 5298 |
| Unmanaged Stand | |||
| Inventory year | 1960 | 1980 | 2010 |
| Total number of measured trees | 5467 | 6314 | 6591 |
| Partly Salvaged Stand | ||||||
|---|---|---|---|---|---|---|
| Beech 1955 | Beech 1975 | Beech 2005 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 20 | 0.35 | 13 | 0.23 | 5 | 0.09 |
| small trees | 21 | 1.04 | 15 | 0.71 | 6 | 0.31 |
| medium trees | 35 | 4.46 | 32 | 3.99 | 20 | 2.72 |
| large trees | 37 | 13.00 | 37 | 13.89 | 32 | 12.07 |
| Ʃ | 113 | 18.85 | 96 | 18.82 | 63 | 15.19 |
| Fir 1955 | Fir 1975 | Fir 2005 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 210 | 3.70 | 205 | 3.63 | 128 | 2.26 |
| small trees | 84 | 4.13 | 99 | 4.84 | 62 | 3.05 |
| medium trees | 65 | 7.41 | 91 | 10.87 | 81 | 10.12 |
| large trees | 9 | 2.36 | 27 | 7.59 | 52 | 16.81 |
| Ʃ | 367 | 17.60 | 422 | 26.93 | 323 | 32.24 |
| Spruce 1955 | Spruce 1975 | Spruce 2005 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 19 | 0.34 | 16 | 0.28 | 9 | 0.16 |
| small trees | 7 | 0.35 | 9 | 0.45 | 4 | 0.18 |
| medium trees | 6 | 0.68 | 8 | 1.01 | 6 | 0.69 |
| large trees | 0 | 0.00 | 2 | 0.45 | 3 | 1.05 |
| Ʃ | 32 | 1.36 | 35 | 2.20 | 22 | 2.09 |
| Total trees 1955 | Total trees 1975 | Total trees 2005 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 249 | 4.39 | 234 | 4.14 | 143 | 2.53 |
| small trees | 113 | 5.52 | 123 | 6.02 | 73 | 3.57 |
| medium trees | 106 | 12.62 | 131 | 15.91 | 107 | 13.58 |
| large trees | 46 | 15.56 | 66 | 22.17 | 88 | 30.18 |
| Ʃ | 513 | 38.09 | 555 | 48.23 | 412 | 49.86 |
| Selection Stand | ||||||
|---|---|---|---|---|---|---|
| Beech 1960 | Beech 1980 | Beech 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 25 | 0.43 | 32 | 0.57 | 47 | 0.83 |
| small trees | 19 | 0.94 | 15 | 0.75 | 14 | 0.67 |
| medium trees | 26 | 3.15 | 22 | 2.77 | 17 | 2.13 |
| large trees | 19 | 6.43 | 13 | 4.17 | 13 | 4.23 |
| Ʃ | 88 | 10.95 | 83 | 8.26 | 91 | 7.86 |
| Fir 1960 | Fir 1980 | Fir 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 185 | 3.26 | 167 | 2.96 | 154 | 2.72 |
| small trees | 78 | 3.81 | 74 | 3.62 | 49 | 2.41 |
| medium trees | 72 | 8.55 | 81 | 9.76 | 59 | 7.36 |
| large trees | 11 | 3.06 | 24 | 6.57 | 32 | 9.55 |
| Ʃ | 346 | 18.68 | 346 | 22.90 | 294 | 22.03 |
| Spruce 1960 | Spruce 1980 | Spruce 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 23 | 0.41 | 23 | 0.41 | 27 | 0.48 |
| small trees | 13 | 0.63 | 13 | 0.62 | 11 | 0.54 |
| medium trees | 13 | 1.50 | 16 | 1.97 | 14 | 1.81 |
| large trees | 2 | 0.45 | 5 | 1.25 | 9 | 2.80 |
| Ʃ | 50 | 2.98 | 57 | 4.25 | 62 | 5.63 |
| Total trees 1960 | Total trees 1980 | Total trees 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 238 | 4.19 | 233 | 4.12 | 243 | 4.30 |
| small trees | 112 | 5.48 | 104 | 5.12 | 79 | 3.87 |
| medium trees | 114 | 13.61 | 122 | 14.84 | 94 | 11.70 |
| large trees | 34 | 10.69 | 43 | 12.43 | 57 | 17.09 |
| Ʃ | 497 | 33.97 | 502 | 36.50 | 473 | 36.96 |
| Unmanaged Stand | ||||||
|---|---|---|---|---|---|---|
| Beech 1960 | Beech 1980 | Beech 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 9 | 0.16 | 4 | 0.07 | 3 | 0.05 |
| small trees | 15 | 0.75 | 12 | 0.57 | 7 | 0.32 |
| medium trees | 15 | 1.70 | 18 | 2.18 | 14 | 1.84 |
| large trees | 8 | 2.56 | 12 | 4.21 | 18 | 6.30 |
| Ʃ | 47 | 5.17 | 45 | 7.03 | 42 | 8.52 |
| Fir 1960 | Fir 1980 | Fir 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 69 | 1.21 | 83 | 1.47 | 120 | 2.12 |
| small trees | 43 | 2.09 | 44 | 2.14 | 31 | 1.54 |
| medium trees | 65 | 8.24 | 58 | 7.28 | 47 | 5.93 |
| large trees | 28 | 8.32 | 45 | 14.15 | 50 | 16.64 |
| Ʃ | 204 | 19.86 | 229 | 25.04 | 248 | 26.23 |
| Spruce 1960 | Spruce 1980 | Spruce 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 7 | 0.12 | 46 | 0.81 | 48 | 0.86 |
| small trees | 9 | 0.46 | 11 | 0.52 | 15 | 0.76 |
| medium trees | 45 | 6.18 | 26 | 3.63 | 21 | 2.67 |
| large trees | 30 | 8.52 | 42 | 13.12 | 47 | 16.28 |
| Ʃ | 92 | 15.28 | 125 | 18.08 | 132 | 20.57 |
| Total trees 1960 | Total trees 1980 | Total trees 2010 | ||||
| Size category | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) | stems ha−1 | BA (m2 ha−1) |
| pole trees | 85 | 1.50 | 134 | 2.37 | 172 | 3.03 |
| small trees | 68 | 3.35 | 66 | 3.24 | 53 | 2.62 |
| medium trees | 133 | 17.22 | 107 | 13.86 | 84 | 10.72 |
| large trees | 69 | 20.37 | 103 | 32.76 | 119 | 40.34 |
| Ʃ | 355 | 42.43 | 410 | 52.22 | 428 | 56.71 |
| LTIba Index Values | |||
|---|---|---|---|
| Partly Salvaged Stand | Selection Stand | Unmanaged Stand | |
| Beech | −0.07 | −0.34 | 1.46 |
| Fir | 6.12 | 2.12 | 1.00 |
| Spruce | na | 5.19 | 0.91 |
| All species | 0.94 | 0.60 | 0.98 |
References
- Davis, M.B. Extent and location. In Eastern Old-Growth Forests: Prospects for Rediscovery and Recovery; Davis, M.B., Ed.; Island Press: Washington, DC, USA, 1996; pp. 18–36. [Google Scholar]
- Parviainen, J.; Bücking, W.; Vandekerkhove, K.; Schuck, A.; Päivinen, R. Strict forest reserves in Europe: Efforts to enhance biodiversity and research on forests left for free development in Europe (EU-COST-Action E4). Forestry 2000, 73, 107–118. [Google Scholar] [CrossRef]
- Global Forest Resource Assessment 2010. Available online: http://www.fao.org/docrep/013/i1757e/i1757e.pdf (accessed on 17 May 2018).
- Keeton, W.S. Managing for late-successional/old-growth characteristics in northern hardwood-conifer forests. For. Ecol. Manag. 2006, 235, 129–142. [Google Scholar] [CrossRef]
- Côté, P.; Tittler, R.; Messier, C.; Kneeshaw, D.D.; Fall, A.; Fortin, M.J. Comparing different forest zoning options for landscape-scale management of the boreal forest: Possible benefits of the TRIAD. For. Ecol. Manag. 2010, 259, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Schütz, J.P.; Saniga, M.; Diaci, J.; Vrška, T. Comparing close-to-nature silviculture with processes in pristine forests: Lessons from Central Europe. Ann. For. Sci. 2016, 73, 911–921. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Ford, S.E.; Keeton, W.S. Enhanced carbon storage through management for old-growth characteristics in northern hardwood-conifer forests. Ecosphere 2017, 8, e01721. [Google Scholar] [CrossRef] [Green Version]
- Kimmins, J.P.; Rempel, R.; Welham, C.; Seely, B.; Rees, V.K. Biophysical sustainability, process-based monitoring and forest ecosystem management decision support systems. For. Chron. 2007, 83, 502–514. [Google Scholar] [CrossRef] [Green Version]
- Paillet, Y.; Pernot, C.; Boulanger, V.; Debaive, N.; Fuhr, M.; Gilg, O.; Gosselin, F. Quantifying the recovery of old-growth attributes in forest reserves: A first reference for France. For. Ecol. Manag. 2015, 346, 51–64. [Google Scholar] [CrossRef]
- Alessandrini, A.; Biondi, F.; Di, A.; Ziaco, E.; Piovesan, G. Tree size distribution at increasing spatial scales converges to the rotated sigmoid curve in two old-growth beech stands of the Italian Apennines. For. Ecol. Manag. 2011, 262, 1950–1962. [Google Scholar] [CrossRef]
- Diaci, J.; Rozenbergar, D.; Anic, I.; Mikac, S.; Saniga, M.; Kucbel, S.; Visnjic, C.; Ballian, D. Structural dynamics and synchronous silver fir decline in mixed old-growth mountain forests in Eastern and Southeastern Europe. Forestry 2011, 84, 479–491. [Google Scholar] [CrossRef] [Green Version]
- Motta, R.; Garbarino, M.; Berretti, R.; Meloni, F.; Nosenzo, A.; Vacchiano, G. Development of old-growth characteristics in uneven-aged forests of the Italian Alps. Eur. J. For. Res. 2015, 134, 19–31. [Google Scholar] [CrossRef]
- Dieler, J.; Uhl, E.; Biber, P.; Müller, J.; Rötzer, T.; Pretzsch, H. Effect of forest stand management on species composition, structural diversity, and productivity in the temperate zone of Europe. Eur. J. For. Res. 2017, 136, 739–766. [Google Scholar] [CrossRef]
- Spies, T.A.; Franklin, J.F. The Diversity and Maintenance of Old-Growth Forests. In Biodiversity in Managed Landscapes: Theory and Practice; Szaro, R.C., Johnston, D., Eds.; Oxford University Press: New York, NY, USA, 1996; pp. 296–314. ISBN 0-19-507958-2. [Google Scholar]
- Gronewold, C.A.; D’Amato, A.W.; Palik, B.J. The influence of cutting cycle and stocking level on the structure and composition of managed old-growth northern hardwoods. For. Ecol. Manag. 2010, 259, 1151–1160. [Google Scholar] [CrossRef]
- Hanson, J.J.; Lorimer, C.G.; Halpin, C.R.; Palik, B.J. Ecological forestry in an uneven-aged, late-successional forest: Simulated effects of contrasting treatments on structure and yield. For. Ecol. Manag. 2012, 270, 94–107. [Google Scholar] [CrossRef]
- Curtis, R.; Marshall, D.D. Douglas-Fir Rotations-Time for Reappraisal? West. J. Appl. For. 1993, 8, 81–85. [Google Scholar]
- Silver, E.J.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Bradford, J.B. Structure and development of old-growth, unmanaged second-growth, and extended rotation Pinus resinosa forests in Minnesota, USA. For. Ecol. Manag. 2013, 291, 110–118. [Google Scholar] [CrossRef]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of european forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Pretzsch, H. Diversity and productivity in forests: Evidence from long-term experimental plots. In Forest Diversity and Function: Temperate and Boreal Systems; Scherer-Lorenzen, M., Korner, C., Schulze, E.-D., Eds.; Springer: Berlin, Germany, 2005; Volume 176, pp. 41–64. ISBN 978-3-540-22191-3. [Google Scholar]
- Fischer, C.; Mölder, A. Trend to increasing structural diversity in German forests: Results from National Forest Inventories 2002 and 2012. Ann. For. Sci. 2017, 74, 80. [Google Scholar] [CrossRef]
- Spathelf, P.; Bolte, A.; van der Maaten, E. Is Close-to-Nature Silviculture (CNS) an adequate concept to adapt forests to climate change? Landbauforsch. App. Agric. For. Res. 2015, 65, 161–170. [Google Scholar] [CrossRef]
- Bončina, A. History, current status and future prospects of uneven-aged forest management in the Dinaric region: An overview. Forestry 2011, 84, 467–478. [Google Scholar] [CrossRef]
- Keren, S.; Diaci, J.; Motta, R.; Govedar, Z. Stand structural complexity of mixed old-growth and adjacent selection forests in the Dinaric Mountains of Bosnia and Herzegovina. For. Ecol. Manag. 2017, 400, 531–541. [Google Scholar] [CrossRef]
- Bosela, M.; Tobin, B.; Šeben, V.; Petráš, R.; Larocque, G.R. Different mixtures of Norway spruce, silver fir, and European beech modify competitive interactions in central European mature mixed forests. Can. J. For. Res. 2015, 45, 1577–1586. [Google Scholar] [CrossRef]
- Bankovic, S.; Medarevic, M.; Pantic, D.; Petrovic, N.; Sljukic, B.; Obradovic, S. The growing stock of the Republic of Serbia: State and problems. Bull. Fac. For. 2009, 100, 7–30. [Google Scholar] [CrossRef]
- Medarević, M.; Banković, S.; Pantić, D.; Obradović, S. Effects of the control method (Goč variety) in selection forest management in western Serbia. Arch. Biol. Sci. 2010, 62, 407–418. [Google Scholar] [CrossRef]
- Diaci, J.; Rozenbergar, D.; Fidej, G.; Nagel, T.A. Challenges for uneven-aged silviculture in restoration of post-disturbance forests in central Europe: A synthesis. Forests 2017, 8, 378. [Google Scholar] [CrossRef]
- Lang, K.D.; Schulte, L.A.; Guntenspergen, G.R. Windthrow and salvage logging in an old-growth hemlock-northern hardwoods forest. For. Ecol. Manag. 2009, 259, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Fidej, G.; Rozman, A.; Diaci, J. Drivers of regeneration dynamics following salvage logging and different silvicultural treatments in windthrow areas in Slovenia. For. Ecol. Manag. 2018, 409, 378–389. [Google Scholar] [CrossRef]
- Thorn, S.; Bässler, C.; Svoboda, M.; Müller, J. Effects of natural disturbances and salvage logging on biodiversity—Lessons from the Bohemian Forest. For. Ecol. Manag. 2017, 388, 113–119. [Google Scholar] [CrossRef]
- Thorn, S.; Bässler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.Y.; Cobb, T.; Donato, D.C.; et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 2018, 55, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.A.; Mikac, S.; Dolinar, M.; Klopcic, M.; Keren, S.; Svoboda, M.; Diaci, J.; Boncina, A.; Paulic, V. The natural disturbance regime in forests of the Dinaric Mountains: A synthesis of evidence. For. Ecol. Manag. 2017, 388, 29–42. [Google Scholar] [CrossRef]
- Klopčič, M.; Simončič, T.; Bončina, A. Comparison of regeneration and recruitment of shade-tolerant and light-demanding tree species in mixed uneven-aged forests: Experiences from the Dinaric region. Forestry 2015, 88, 552–563. [Google Scholar] [CrossRef]
- Klopčič, M.; Mina, M.; Bugmann, H.; Bončina, A. The prospects of silver fir (Abies alba Mill.) and Norway spruce (Picea abies (L.) Karst) in mixed mountain forests under various management strategies, climate change and high browsing pressure. Eur. J. For. Res. 2017, 136, 1071–1090. [Google Scholar] [CrossRef]
- Vasić, M. Tara planina, Šumarska Studija; N. štamparija „Davidović“: Belgrade, Serbia, 1908. [Google Scholar]
- Milojković, D. Razvoj gazdovanja i uređivanja šuma GJ Tara. Šumarstvo 1986, 1–2, 11–27. [Google Scholar]
- Obradović, S. Stanje i Razvoj Sastojina Bukve, jele i Smrče Prašumskog Porekla u Srbiji kao Osnov za Planiranje i obezbeđivanje Prirodi Bliskog Gazdovanja. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 2015. [Google Scholar]
- Pantic, D.; Medarevic, M.; Bankovic, S.; Obradovic, S.; Sljukic, B.; Pesic, B. Structural, production and dynamic characteristics of the strict forest reserve “Račanska šljivovica” on mt. Tara. Bull. Fac. For. 2011, 93–114. [Google Scholar] [CrossRef]
- Bončina, A.; Čavlović, J.; Čurović, M.; Govedar, Z.; Klopčič, M.; Medarević, M. A comparative analysis of recent changes in Dinaric uneven-aged forests of the NW Balkans. Forestry 2014, 87, 71–84. [Google Scholar] [CrossRef]
- Janowiak, M.K.; Nagel, L.M.; Webster, C.R. Spatial Scale and Stand Structure in Northern Hardwood Forests: Implications for Quantifying Diameter Distributions. For. Sci. 2008, 54, 497–506. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002; ISBN 0-387-95364-7. [Google Scholar]
- Zenner, E.K.; Peck, J.L.E. Floating neighborhoods reveal contribution of individual trees to high sub-stand scale heterogeneity. For. Ecol. Manag. 2018, 412, 29–40. [Google Scholar] [CrossRef]
- Lafond, V.; Cordonnier, T.; De Coligny, F.; Courbaud, B. Reconstructing harvesting diameter distribution from aggregate data. Ann. For. Sci. 2012, 69, 235–243. [Google Scholar] [CrossRef]
- Klopcic, M.; Boncina, A. Stand dynamics of silver fir (Abies alba Mill.)-European beech (Fagus sylvatica L.) forests during the past century: A decline of silver fir? Forestry 2011, 84, 259–271. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Bobiec, A.; Eycott, A.E. Lack of demographic equilibrium indicates natural, large-scale forest dynamics, not a problematic forest conservation policy—A reply to Brzeziecki et al. J. Veg. Sci. 2017, 28, 218–222. [Google Scholar] [CrossRef]
- Fassnacht, K.S.; Bronson, D.R.; Palik, B.J.; Amato, A.W.D.; Lorimer, C.G.; Martin, K.J. Accelerating the Development of Old-Growth Characteristics in Second-Growth Northern Hardwoods; General Technical Report; U.S. Depoartment of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2015.
- Brzeziecki, B.; Pommerening, A.; Miścicki, S.; Drozdowski, S.; Zybura, H. A common lack of demographic equilibrium among tree species in Bialowieża National Park (NE Poland): Evidence from long-term plots. J. Veg. Sci. 2016, 27, 460–469. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Niemczyk, M.; Olszowska, G. The influence of stand structure on European yew Taxus baccata populations in its natural habitats in central Poland. Pol. J. Ecol. 2017, 65, 369–384. [Google Scholar] [CrossRef]
- Čavlović, J.; Božić, M.; Bončina, A. Stand structure of an uneven-aged fir-beech forest with an irregular diameter structure: Modeling the development of the Belevine forest, Croatia. Eur. J. For. Res. 2006, 125, 325–333. [Google Scholar] [CrossRef]
- Petritan, I.C.; Commarmot, B.; Hobi, M.L.; Petritan, A.M.; Bigler, C.; Abrudan, I.V.; Rigling, A. Structural patterns of beech and silver fir suggest stability and resilience of the virgin forest Sinca in the Southern Carpathians, Romania. For. Ecol. Manag. 2015, 356, 184–195. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Gersonde, R.F. Stocking control concepts in uneven-aged silviculture. Forestry 2004, 77, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Schütz, J.P. Die Plenterung und ihre Unterschiedlichen Formen Skript zu Vorlesung Waldbau II und Waldbau IV; ETH Zürich: Zürich, Switzerland, 2002. [Google Scholar]
- Goodburn, J.M.; Lorimer, C.G. Population structure in old-growth and managed northern hardwoods: An examination of the balanced diameter distribution concept. For. Ecol. Manag. 1999, 118, 11–29. [Google Scholar] [CrossRef]
- Motta, R.; Berretti, R.; Castagneri, D.; Dukić, V.; Garbarino, M.; Govedar, Z.; Lingua, E.; Maunaga, Z.; Meloni, F. Toward a definition of the range of variability of central European mixed Fagus–Abies–Picea forests: The nearly steady-state forest of Lom (Bosnia and Herzegovina). Can. J. For. Res. 2011, 41, 1871–1884. [Google Scholar] [CrossRef]
- Schwartz, J.W.; Nagel, L.M.; Webster, C.R. Effects of uneven-aged management on diameter distribution and species composition of northern hardwoods in Upper Michigan. For. Ecol. Manag. 2005, 211, 356–370. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Hasenauer, H.; Kindermann, G. Sustainability in multi-aged stands: An analysis of long-term plenter systems. Forestry 2007, 80, 163–181. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rötzer, T. Indicating Forest Ecosystem and Stand Productivity: From Deductive to Inductive Concepts. In Ecological Forest Management Handbook, Series of Applied Ecology and Environmental Management; Laroque, G.R., Ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2016; pp. 415–444. [Google Scholar]
- Jaloviar, P.; Saniga, M.; Kucbel, S.; Pittner, J.; Vencurik, J.; Dovciak, M. Seven decades of change in a European old-growth forest following a stand-replacing wind disturbance: A long-term case study. For. Ecol. Manag. 2017, 399, 197–205. [Google Scholar] [CrossRef]
- Keren, S.; Motta, R.; Govedar, Z.; Lucic, R.; Medarevic, M.; Diaci, J. Comparative structural dynamics of the Janj mixed old-growth mountain forest in Bosnia and Herzegovina: Are conifers in a long-term decline? Forests 2014, 5, 1243–1266. [Google Scholar] [CrossRef]
- Marinšek, A.; Diaci, J. A comparison of structural characteristics and ecological factors between forest reserves and managed silver fir: Norway spruce forests in Slovenia. Ekológia (Bratislava) 2011, 30, 51–66. [Google Scholar] [CrossRef]
- Szwagrzyk, J.; Szewczyk, J.; Maciejewski, Z. Shade-tolerant tree species from temperate forests differ in their competitive abilities: A case study from Roztocze, south-eastern Poland. For. Ecol. Manag. 2012, 282, 28–35. [Google Scholar] [CrossRef]
- Vrška, T.; Adam, D.; Hort, L.; Kolář, T.; Janík, D. European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) rotation in the Carpathians - A developmental cycle or a linear trend induced by man? For. Ecol. Manag. 2009, 258, 347–356. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and silviculture of silver fir (Abies alba Mill.): A review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Keren, S.; Diaci, J. Comparing the quantity and structure of deadwood in selection managed and old-growth forests in South-East Europe. Forests 2018, 9, 76. [Google Scholar] [CrossRef]
- Motta, R.; Garbarino, M.; Berretti, R.; Bjelanovic, I.; Borgogno Mondino, E.; Čurović, M.; Keren, S.; Meloni, F.; Nosenzo, A. Structure, spatio-temporal dynamics and disturbance regime of the mixed beech–silver fir–Norway spruce old-growth forest of Biogradska Gora (Montenegro). Plant Biosyst. 2015, 149, 966–975. [Google Scholar] [CrossRef]
- Marage, D.; Lemperiere, G. The management of snags: A comparison in managed and unmanaged ancient forests of the Southern French Alps. Ann. For. Sci. 2005, 62, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Mason, F.; Nardi, G.; Whitmore, D. Recherches sur la restauration des habitats du bois mort: L’exemple du LIFE “Bosco della Fontana” (Italie). In Bois Mort et à Cavités, Une clé Pour Des Forêts Vivantes; Vallauri, D., André, J., Dodelin, B., Eynard-Machet, R., Rambaud, D., Eds.; Tec & Doc: Paris, France, 2005; pp. 285–291. [Google Scholar]





| Partly Salvaged Stand (1955) | Selection Stand (1960) | Unmanaged Stand (1960) | ||||||||||
| Fir | Spruce | Beech | Total | Fir | Spruce | Beech | Total | Fir | Spruce | Beech | Total | |
| RMSE | 0.14 | 0.06 | 0.09 | 0.07 | 0.03 | 0.03 | 0.06 | 0.08 | 0.07 | 0.13 | 0.09 | 0.06 |
| Adjusted R2 | 0.97 | 0.98 | 0.96 | 0.99 | 0.98 | 0.99 | 0.99 | 0.94 | 0.99 | 0.91 | 0.95 | 0.98 |
| p | 0.0000 | 0.0054 | 0.0000 | 0.0000 | 0.0012 | 0.0032 | 0.0000 | 0.0015 | 0.0000 | 0.0015 | 0.0000 | 0.0000 |
| AIC | −10.3 | −8.5 | −17.4 | −22.9 | −12.0 | −15.9 | −39.3 | −25.2 | −19.5 | −3.1 | −13.7 | −16.0 |
| Shape | NE | NE | IQ | NE | RS | NE | IQ | NE | IQ | UNI | IQ | IQ |
| Partly Salvaged Stand (1975) | Selection Stand (1980) | Unmanaged Stand (1980) | ||||||||||
| Fir | Spruce | Beech | Total | Fir | Spruce | Beech | Total | Fir | Spruce | Beech | Total | |
| RMSE | 0.08 | 0.07 | 0.05 | 0.05 | 0.06 | 0.03 | 0.06 | 0.07 | 0.06 | 0.24 | 0.13 | 0.05 |
| Adjusted R2 | 0.99 | 0.98 | 0.98 | 0.99 | 0.98 | 0.99 | 0.98 | 0.93 | 0.98 | 0.80 | 0.86 | 0.98 |
| p | 0.0000 | 0.0000 | 0.0004 | 0.0000 | 0.0001 | 0.0001 | 0.0000 | 0.0018 | 0.0001 | 0.023 | 0.0004 | 0.0001 |
| AIC | −19.0 | −13.7 | −27.7 | −29.2 | −11.3 | −16.1 | −23.9 | −18.4 | −14.5 | 3.8 | −6.2 | −9.5 |
| Shape | NE | NE | UNI | NE | RS | RS | NE | NE | RS | IQ | IQ | RS |
| Partly Salvaged Stand (2005) | Selection Stand (2010) | Unmanaged Stand (2010) | ||||||||||
| Fir | Spruce | Beech | Total | Fir | Spruce | Beech | Total | Fir | Spruce | Beech | Total | |
| RMSE | 0.11 | 0.08 | 0.07 | 0.09 | 0.03 | 0.06 | 0.09 | 0.09 | 0.10 | 0.07 | 0.06 | 0.06 |
| Adjusted R2 | 0.98 | 0.94 | 0.95 | 0.98 | 0.99 | 0.99 | 0.97 | 0.98 | 0.95 | 0.97 | 0.95 | 0.99 |
| p | 0.0000 | 0.0004 | 0.0000 | 0.0000 | 0.0001 | 0.0001 | 0.0005 | 0.0001 | 0.0017 | 0.0001 | 0.0000 | 0.0000 |
| AIC | −13.8 | −11.5 | −21.6 | −18.3 | −8.4 | −19.5 | −11.3 | −12.2 | −13.3 | −16.7 | −21.8 | −21.1 |
| Shape | RS | IQ | UNI | IQ | RS | RS | RS | RS | RS | RS | UNI | RS |
| Stem Density Per Hectare (%) | ||||||
|---|---|---|---|---|---|---|
| Stand Category | Year | Beech (%) | Fir (%) | Spruce (%) | Maple (%) | Other sp. (%) |
| Partly Salvaged Stand | 1955 | 21.9 | 71.5 | 6.3 | 0.3 | 0.0 |
| 1975 | 17.4 | 76.0 | 6.4 | 0.2 | 0.0 | |
| 2005 | 15.3 | 78.6 | 5.4 | 0.7 | 0.0 | |
| Selection Stand | 1960 | 17.8 | 69.5 | 10.1 | 1.5 | 1.1 |
| 1980 | 16.5 | 68.8 | 11.3 | 1.7 | 1.7 | |
| 2010 | 19.2 | 62.2 | 13.1 | 3.6 | 1.9 | |
| Unmanaged Stand | 1960 | 13.2 | 57.3 | 25.9 | 0.2 | 3.4 |
| 1980 | 11.0 | 56.0 | 30.5 | 0.2 | 2.3 | |
| 2010 | 9.7 | 58.1 | 30.9 | 0.2 | 1.1 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keren, S.; Medarević, M.; Obradović, S.; Zlokapa, B. Five Decades of Structural and Compositional Changes in Managed and Unmanaged Montane Stands: A Case Study from South-East Europe. Forests 2018, 9, 479. https://doi.org/10.3390/f9080479
Keren S, Medarević M, Obradović S, Zlokapa B. Five Decades of Structural and Compositional Changes in Managed and Unmanaged Montane Stands: A Case Study from South-East Europe. Forests. 2018; 9(8):479. https://doi.org/10.3390/f9080479
Chicago/Turabian StyleKeren, Srđan, Milan Medarević, Snežana Obradović, and Brane Zlokapa. 2018. "Five Decades of Structural and Compositional Changes in Managed and Unmanaged Montane Stands: A Case Study from South-East Europe" Forests 9, no. 8: 479. https://doi.org/10.3390/f9080479
APA StyleKeren, S., Medarević, M., Obradović, S., & Zlokapa, B. (2018). Five Decades of Structural and Compositional Changes in Managed and Unmanaged Montane Stands: A Case Study from South-East Europe. Forests, 9(8), 479. https://doi.org/10.3390/f9080479