Defective Interfering RNAs: Foes of Viruses and Friends of Virologists

Abstract
:1. Introduction
2. Origin and Synthesis of Defective Interfering (DI) RNAs
2.1. Mechanisms of generation of DI-RNAs
2.1.1. Replicase driven template-switching mechanism
2.1.2. Forced template-switching mechanism

2.1.3. RNA breakage and ligation mechanism
2.2. Viral replication proteins as factors influencing the formation/accumulation of DI-RNAs

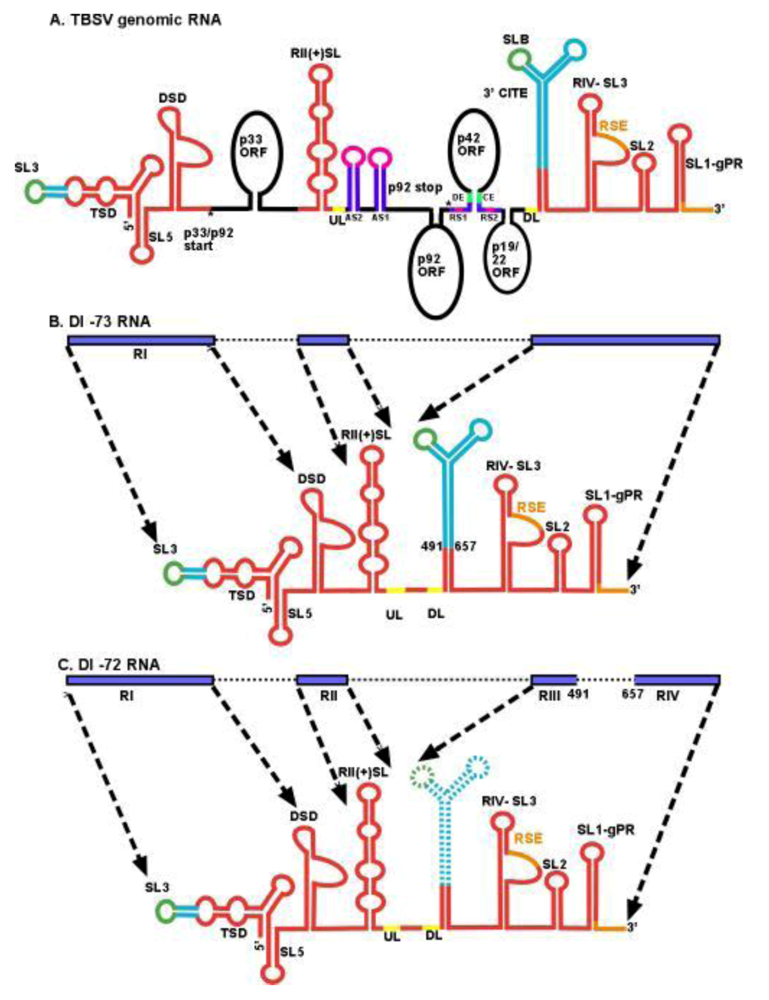
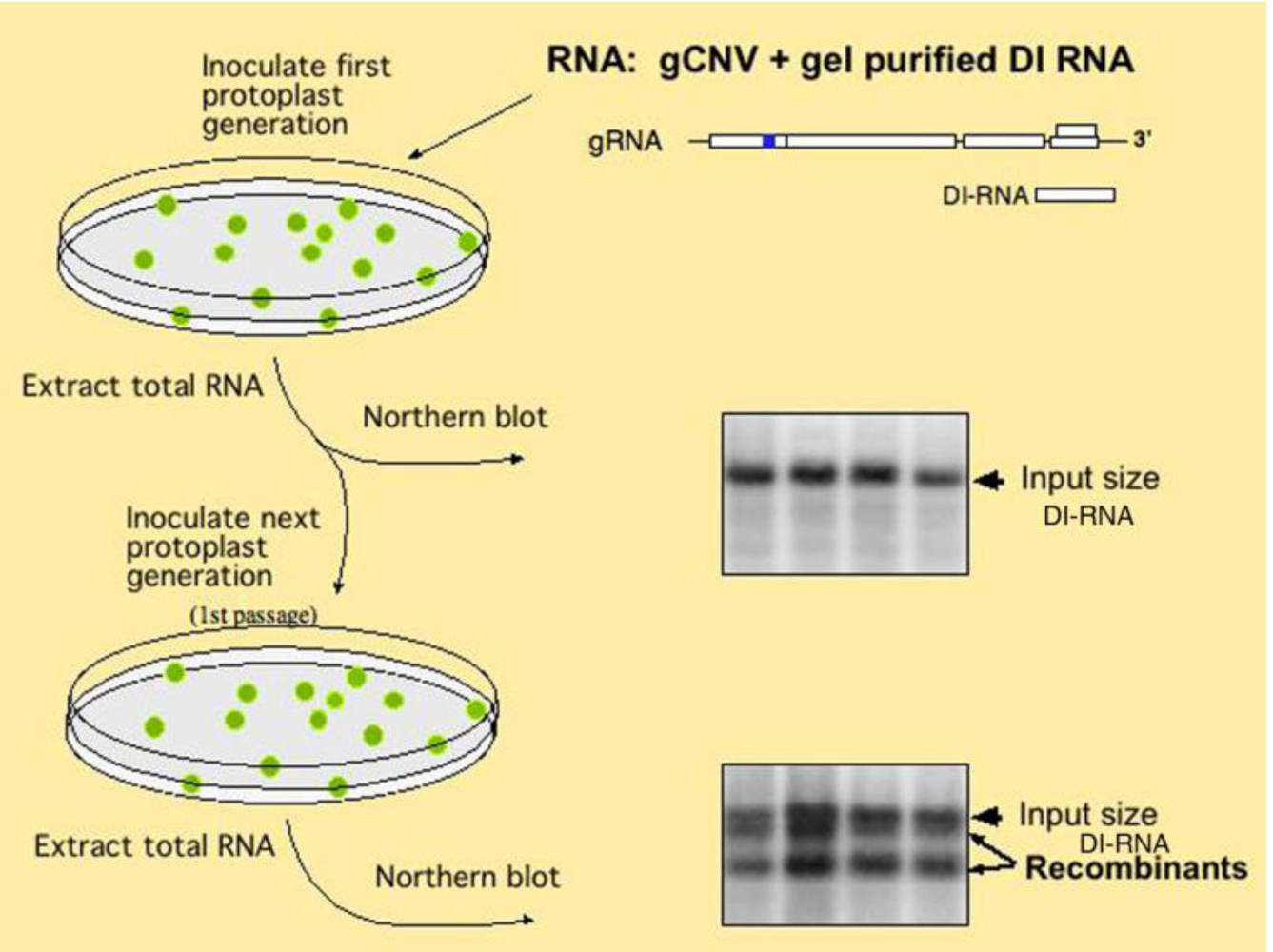
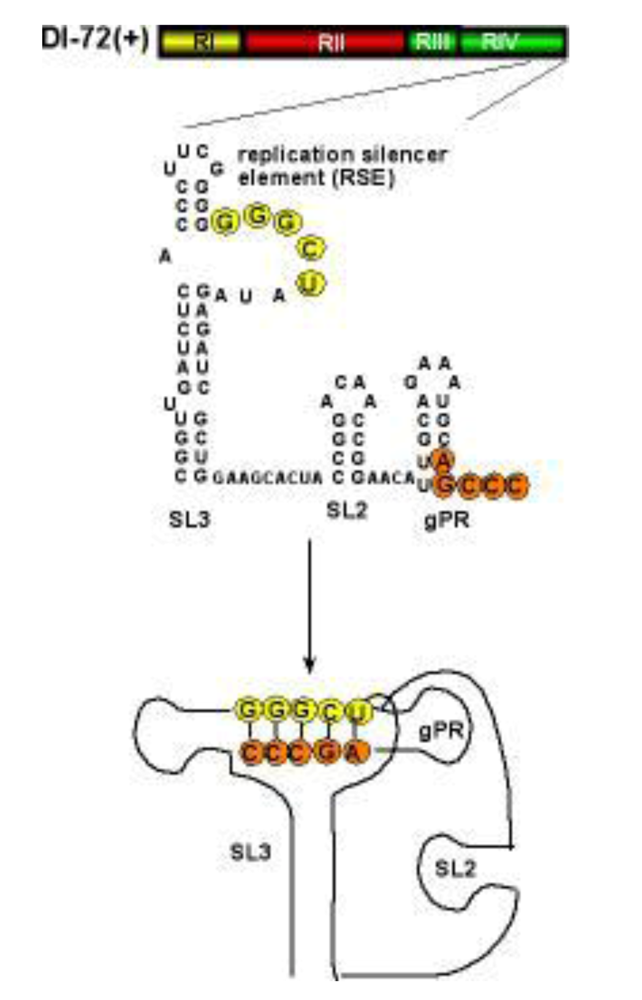
2.3. Viral RNA elements as recombination “hot spots” during DI-RNA formation
2.4. The effect of host factors on DI-RNA formation
2.5. Environmental and other factors affecting the formation of DI-RNAs
3. Continuous Evolution of DI-RNAs
3.1. Deletion
3.2. Duplication and nucleotide insertions
4. The Mechanism of Interference by DI-RNA
4.3. DI-RNA-triggered gene silencing response of the host
5. DI-RNA as a Tool in Virology and Biotechnology Applications
5.1. DI-RNAs as vectors to express recombinant proteins in plants
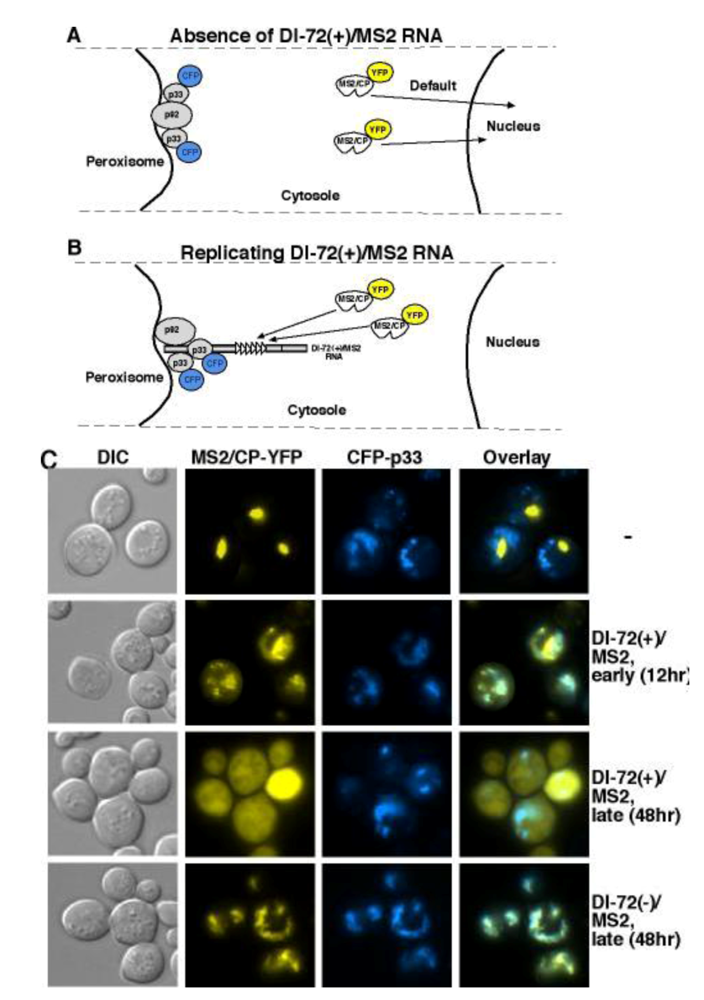
5.2. DI-RNAs as surrogate templates to study virus replication
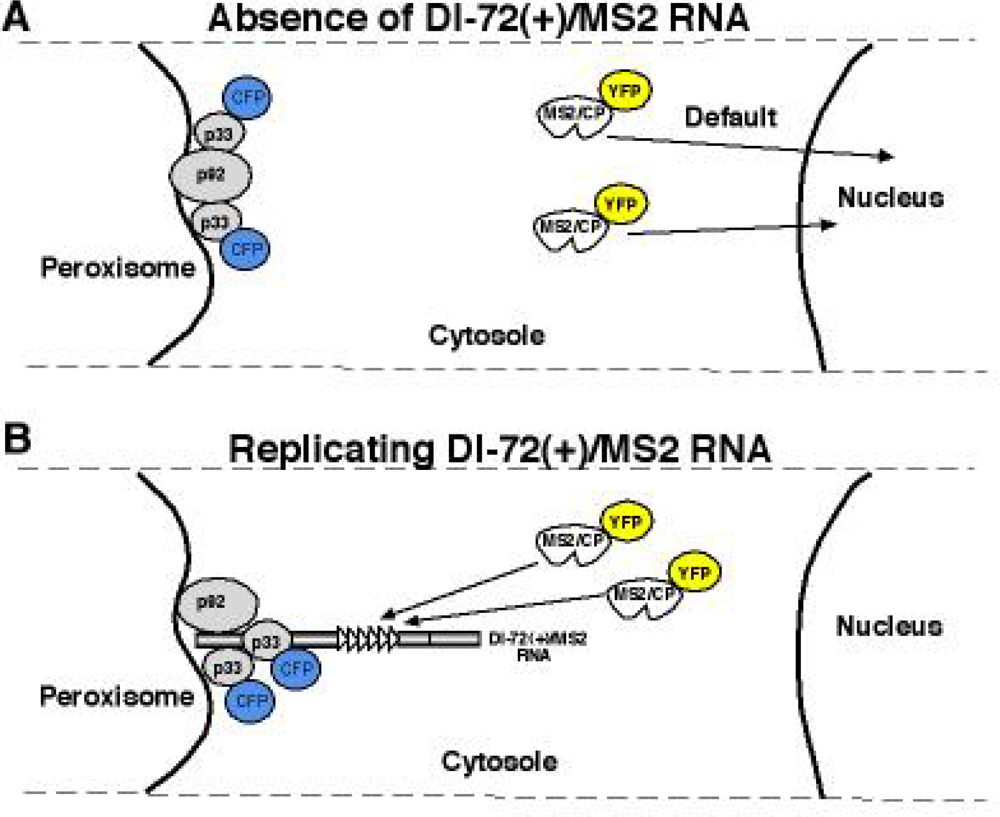
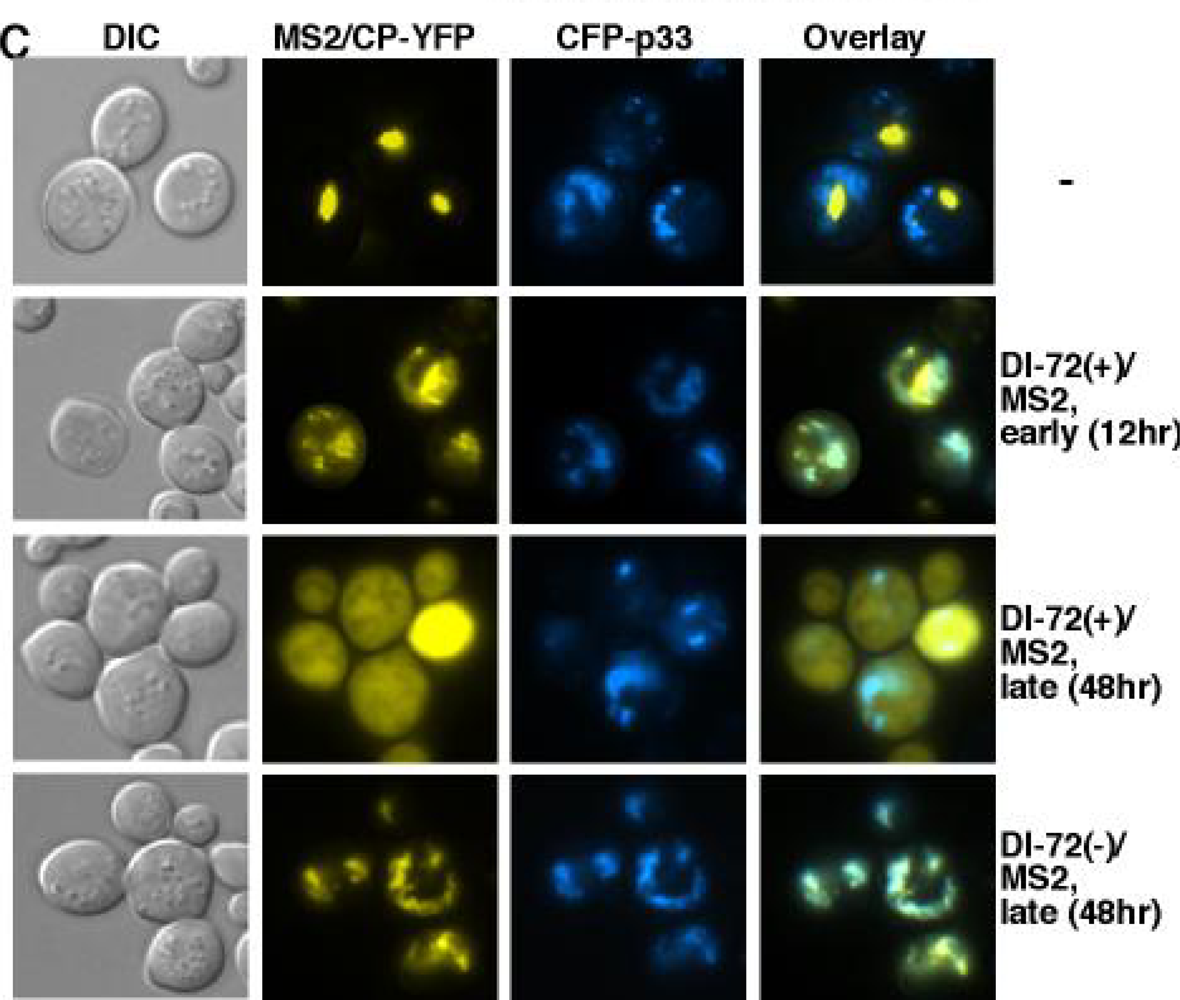
5.3. Template selection and DI-RNA recruitment into replication

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
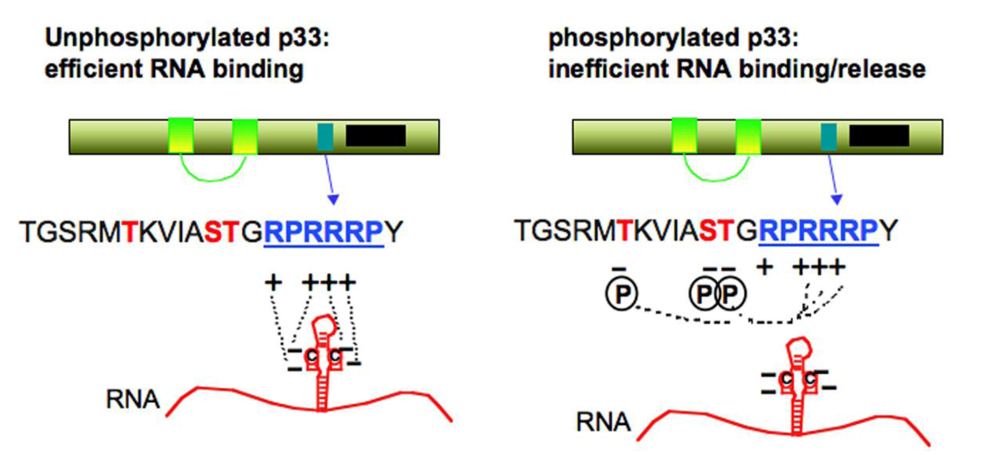
5.5. Factors affecting the replication of DI-RNA
5.6. Release of (+)DI-RNA progeny from the replicase complex and disassembly of replicase complex


6. Conclusions and Future Prospects
Acknowledgments
References
- Huang, A.S. Defective interfering viruses. Annu. Rev. Microbiol. 1973, 27, 101–117. [Google Scholar] [PubMed]
- Simon, A.E.; Roossinck, M.J.; Havelda, Z. Plant virus satellite and defective interfering RNAs: new paradigms for a new century. Annu. Rev. Phytopathol. 2004, 42, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Perrault, J. Origin and replication of defective interfering particles. Curr. Top. Microbiol. Immunol. 1981, 93, 151–207. [Google Scholar] [PubMed]
- Qiu, W.; Scholthof, K.B. Defective interfering RNAs of a satellite virus. J. Virol. 2001, 75, 5429–5432. [Google Scholar] [CrossRef] [PubMed]
- Luytjes, W.; Gerritsma, H.; Spaan, W.J. Replication of synthetic defective interfering RNAs derived from coronavirus mouse hepatitis virus-A59. Virology 1996, 216, 174–183. [Google Scholar] [CrossRef] [PubMed]
- White, K.A.; Morris, T.J. Nonhomologous RNA recombination in tombusviruses: generation and evolution of defective interfering RNAs by stepwise deletions. J. Virol. 1994, 68, 14–24. [Google Scholar] [PubMed]
- Rochon, D.M. Rapid de novo generation of defective interfering RNA by cucumber necrosis virus mutants that do not express the 20-kDa nonstructural protein. Proc. Natl. Acad. Sci. U S A 1991, 88, 11153–11157. [Google Scholar] [CrossRef] [PubMed]
- Rubino, L.; Burgyan, J.; Grieco, F.; Russo, M. Sequence analysis of cymbidium ringspot virus satellite and defective interfering RNAs. J. Gen. Virol. 1990, 71, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Pogany, J.; Romero, J.; Huang, Q.; Sgro, J.Y.; Shang, H.; Bujarski, J.J. De novo generation of defective interfering-like RNAs in broad bean mottle bromovirus. Virology 1995, 212, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Heaton, L.A.; Morris, T.J.; Simon, A.E. Turnip crinkle virus defective interfering RNAs intensify viral symptoms and are generated de novo. Proc. Natl. Acad. Sci. U S A 1989, 86, 9173–9177. [Google Scholar] [CrossRef] [PubMed]
- Hillman, B.I.; Carrington, J.C.; Morris, T.J. A defective interfering RNA that contains a mosaic of a plant virus genome. Cell 1987, 51, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.; Carette, J.E.; Brown, D.J.; Bol, J.F. Serial passage of tobacco rattle virus under different selection conditions results in deletion of structural and nonstructural genes in RNA 2. J. Virol. 1996, 70, 4933–4940. [Google Scholar] [PubMed]
- Eliasco, E.; Livieratos, I.C.; Muller, G.; Guzman, M.; Salazar, L.F.; Coutts, R.H. Sequences of defective RNAs associated with potato yellow vein virus. Arch. Virol. 2006, 151, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Desvoyes, B.; Scholthof, H.B. Host-dependent recombination of a Tomato bushy stunt virus coat protein mutant yields truncated capsid subunits that form virus-like complexes which benefit systemic spread. Virology 2002, 304, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Ayllon, M.A.; Lopez, C.; Navas-Castillo, J.; Mawassi, M.; Dawson, W.O.; Guerri, J.; Flores, R.; Moreno, P. New defective RNAs from citrus tristeza virus: evidence for a replicase-driven template switching mechanism in their generation. J. Gen. Virol. 1999, 80, 817–821. [Google Scholar] [PubMed]
- White, K.A.; Morris, T.J. Defective and defective interfering RNAs of monopartite plus-strand RNA plant viruses. Curr. Top. Microbiol. Immunol. 1999, 239, 1–17. [Google Scholar] [PubMed]
- White, K.A.; Nagy, P.D. Advances in the molecular biology of tombusviruses: gene expression, genome replication, and recombination. Prog. Nucleic Acid Res. Mol. Biol. 2004, 78, 187–226. [Google Scholar] [PubMed]
- Nagy, P.D.; Simon, A.E. New insights into the mechanisms of RNA recombination. Virology 1997, 235, 1–9. [Google Scholar] [CrossRef]
- Kim, M.J.; Kao, C. Factors regulating template switch in vitro by viral RNA-dependent RNA polymerases: implications for RNA-RNA recombination. Proc. Natl. Acad. Sci. U S A 2001, 98, 4972–4977. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.P.; Nagy, P.D. Mechanism of RNA recombination in carmo- and tombusviruses: evidence for template switching by the RNA-dependent RNA polymerase in vitro. J. Virol. 2003, 77, 12033–12047. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.P.; Pogany, J.; Nagy, P.D. Mechanism of DI RNA formation in tombusviruses: dissecting the requirement for primer extension by the tombusvirus RNA dependent RNA polymerase in vitro. Virology 2002, 304, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Wierzchoslawski, R.; Bujarski, J.J. Efficient in vitro system of homologous recombination in brome mosaic bromovirus. J. Virol. 2006, 80, 6182–6187. [Google Scholar] [CrossRef] [PubMed]
- Wierzchoslawski, R.; Dzianott, A.; Kunimalayan, S.; Bujarski, J.J. A transcriptionally active subgenomic promoter supports homologous crossovers in a plus-strand RNA virus. J. Virol. 2003, 77, 6769–6776. [Google Scholar] [CrossRef] [PubMed]
- Pogany, J.; Nagy, P.D. Authentic replication and recombination of Tomato bushy stunt virus RNA in a cell-free extract from yeast. J. Virol. 2008, 82, 5967–5980. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.P.; Panavas, T.; Luo, G.; Nagy, P.D. Heterologous RNA replication enhancer stimulates in vitro RNA synthesis and template-switching by the carmovirus, but not by the tombusvirus, RNA-dependent RNA polymerase: implication for modular evolution of RNA viruses. Virology 2005, 341, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Simon, A.E. In vitro characterization of late steps of RNA recombination in turnip crinkle virus I. Role of motif1-hairpin structure. Virology 1998, 249, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Simon, A.E. In vitro characterization of late steps of RNA recombination in turnip crinkle virus II. The role of the priming stem and flanking sequences. Virology 1998, 249, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Bujarski, J.J.; Nagy, P.D.; Flasinski, S. Molecular studies of genetic RNA-RNA recombination in brome mosaic virus. Adv. Virus Res. 1994, 43, 275–302. [Google Scholar] [PubMed]
- Havelda, Z.; Szittya, G.; Burgyan, J. Characterization of the molecular mechanism of defective interfering RNA-mediated symptom attenuation in tombusvirus-infected plants. J. Virol. 1998, 72, 6251–6256. [Google Scholar] [PubMed]
- Jaag, H.M.; Nagy, P.D. Silencing of Nicotiana benthamiana Xrn4p exoribonuclease promotes tombusvirus RNA accumulation and recombination. Virology 2009, 386, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.P.; Jaag, H.M.; Jonczyk, M.; Serviene, E.; Nagy, P.D. Expression of the Arabidopsis Xrn4p 5'-3' exoribonuclease facilitates degradation of tombusvirus RNA and promotes rapid emergence of viral variants in plants. Virology 2007, 368, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.P.; Serviene, E.; Nagy, P.D. Suppression of viral RNA recombination by a host exoribonuclease. J. Virol. 2006, 80, 2631–2640. [Google Scholar] [CrossRef] [PubMed]
- Serviene, E.; Shapka, N.; Cheng, C.P.; Panavas, T.; Phuangrat, B.; Baker, J.; Nagy, P.D. Genome-wide screen identifies host genes affecting viral RNA recombination. Proc. Natl. Acad. Sci. U S A 2005, 102, 10545–10550. [Google Scholar] [CrossRef] [PubMed]
- White, K.A.; Morris, T.J. RNA determinants of junction site selection in RNA virus recombinants and defective interfering RNAs. RNA 1995, 1, 1029–1040. [Google Scholar] [PubMed]
- Zhang, X.; Nuss, D.L. A host dicer is required for defective viral RNA production and recombinant virus vector RNA instability for a positive sense RNA virus. Proc. Natl. Acad. Sci. U S A 2008, 105, 16749–16754. [Google Scholar] [CrossRef] [PubMed]
- Chetverin, A.B.; Chetverina, H.V.; Demidenko, A.A.; Ugarov, V.I. Nonhomologous RNA recombination in a cell-free system: evidence for a transesterification mechanism guided by secondary structure. Cell 1997, 88, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Gmyl, A.P.; Korshenko, S.A.; Belousov, E.V.; Khitrina, E.V.; Agol, V.I. Nonreplicative homologous RNA recombination: promiscuous joining of RNA pieces? Rna 2003, 9, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Dzianott, A.; Ahlquist, P.; Bujarski, J.J. Mutations in the helicase-like domain of protein 1a alter the sites of RNA-RNA recombination in brome mosaic virus. J. Virol. 1995, 69, 2547–2556. [Google Scholar] [PubMed]
- Figlerowicz, M.; Nagy, P.D.; Bujarski, J.J. A mutation in the putative RNA polymerase gene inhibits nonhomologous, but not homologous, genetic recombination in an RNA virus. Proc. Natl. Acad. Sci. U S A 1997, 94, 2073–2078. [Google Scholar] [CrossRef] [PubMed]
- Fodor, E.; Mingay, L.J.; Crow, M.; Deng, T.; Brownlee, G.G. A single amino acid mutation in the PA subunit of the influenza virus RNA polymerase promotes the generation of defective interfering RNAs. J. Virol. 2003, 77, 5017–5020. [Google Scholar] [CrossRef] [PubMed]
- Panaviene, Z.; Nagy, P.D. Mutations in the RNA-binding domains of tombusvirus replicase proteins affect RNA recombination in vivo. Virology 2003, 317, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Panaviene, Z.; Baker, J.M.; Nagy, P.D. The overlapping RNA-binding domains of p33 and p92 replicase proteins are essential for tombusvirus replication. Virology 2003, 308, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Jaag, H.M.; Stork, J.; Nagy, P.D. Host transcription factor Rpb11p affects tombusvirus replication and recombination via regulating the accumulation of viral replication proteins. Virology 2007, 368, 388–404. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J.; Simon, A.E. RNA elements required for RNA recombination function as replication enhancers in vitro and in vivo in a plus-strand RNA virus. Embo J. 1999, 18, 5653–5665. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.; White, K.A. Enhancer-like properties of an RNA element that modulates Tombusvirus RNA accumulation. Virology 1999, 256, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Nagy, P.D. The RNA replication enhancer element of tombusviruses contains two interchangeable hairpins that are functional during plus-strand synthesis. J. Virol. 2003, 77, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Bujarski, J.J. Homologous RNA recombination in brome mosaic virus: AU-rich sequences decrease the accuracy of crossovers. J. Virol. 1996, 70, 415–426. [Google Scholar] [PubMed]
- Nagy, P.D.; Bujarski, J.J. Targeting the site of RNA-RNA recombination in brome mosaic virus with antisense sequences. Proc. Natl. Acad. Sci. U S A 1993, 90, 6390–6394. [Google Scholar] [CrossRef] [PubMed]
- Shapka, N.; Nagy, P.D. The AU-rich RNA recombination hot spot sequence of Brome mosaic virus is functional in tombusviruses: implications for the mechanism of RNA recombination. J. Virol. 2004, 78, 2288–2300. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Bujarski, J.J. Engineering of homologous recombination hotspots with AU-rich sequences in brome mosaic virus. J. Virol. 1997, 71, 3799–3810. [Google Scholar] [PubMed]
- DeStefano, J.J.; Bambara, R.A.; Fay, P.J. The mechanism of human immunodeficiency virus reverse transcriptase-catalyzed strand transfer from internal regions of heteropolymeric RNA templates. J. Biol. Chem. 1994, 269, 161–168. [Google Scholar] [PubMed]
- Nagy, P.D.; Bujarski, J.J. Silencing homologous RNA recombination hot spots with GC-rich sequences in brome mosaic virus. J. Virol. 1998, 72, 1122–1130. [Google Scholar] [PubMed]
- Havelda, Z.; Dalmay, T.; Burgyan, J. Secondary structure-dependent evolution of Cymbidium ringspot virus defective interfering RNA. J. Gen. Virol. 1997, 78, 1227–1234. [Google Scholar] [PubMed]
- Pogany, J.; White, K.A.; Nagy, P.D. Specific binding of tombusvirus replication protein p33 to an internal replication element in the viral RNA is essential for replication. J. Virol. 2005, 79, 4859–4869. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Nagy, P.D. Yeast as a model host to study replication and recombination of defective interfering RNA of Tomato bushy stunt virus. Virology 2003, 314, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Serviene, E.; Jiang, Y.; Cheng, C.P.; Baker, J.; Nagy, P.D. Screening of the yeast yTHC collection identifies essential host factors affecting tombusvirus RNA recombination. J. Virol. 2006, 80, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Llamas, S.; Sandoval, C.; Babin, M.; Pogany, J.; Bujarski, J.J.; Romero, J. Effect of the Host and Temperature on the Formation of Defective RNAs Associated with Broad bean mottle virus Infection. Phytopathology 2004, 94, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Omarov, R.T.; Rezende, J.A.; Scholthof, H.B. Host-specific generation and maintenance of Tomato bushy stunt virus defective interfering RNAs. Mol. Plant Microbe Interact. 2004, 17, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J. Mechanisms of plant virus evolution. Annu. Rev. Phytopathol. 1997, 35, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J. Plant RNA virus evolution. Curr. Opin. Microbiol. 2003, 6, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arenal, F.; Fraile, A.; Malpica, J.M. Variation and evolution of plant virus populations. Int. Microbiol. 2003, 6, 225–232. [Google Scholar] [CrossRef] [PubMed]
- White, K.A.; Morris, T.J. Enhanced competitiveness of tomato bushy stunt virus defective interfering RNAs by segment duplication or nucleotide insertion. J. Virol. 1994, 68, 6092–6096. [Google Scholar] [PubMed]
- Wu, B.; White, K.A. Formation and amplification of a novel tombusvirus defective RNA which lacks the 5' nontranslated region of the viral genome. J. Virol. 1998, 72, 9897–9905. [Google Scholar] [PubMed]
- Novak, J.E.; Kirkegaard, K. Coupling between genome translation and replication in an RNA virus. Genes Dev. 1994, 8, 1726–1737. [Google Scholar] [CrossRef]
- Oster, S.K.; Wu, B.; White, K.A. Uncoupled expression of p33 and p92 permits amplification of tomato bushy stunt virus RNAs. J. Virol. 1998, 72, 5845–5851. [Google Scholar] [PubMed]
- Weiland, J.J.; Dreher, T.W. Cis-preferential replication of the turnip yellow mosaic virus RNA genome. Proc. Natl. Acad. Sci. U S A 1993, 90, 6095–6099. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Silhavy, D.; Dalmay, T.; Burgyan, J. Size-dependent cell-to-cell movement of defective interfering RNAs of Cymbidium ringspot virus. J. Gen. Virol. 2002, 83, 1505–1510. [Google Scholar] [PubMed]
- Jones, R.W.; Jackson, A.O.; Morris, T.J. Defective-interfering RNAs and elevated temperatures inhibit replication of tomato bushy stunt virus in inoculated protoplasts. Virology 1990, 176, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Borja, M.; Scholthof, H.B.; Jackson, A.O.; Morris, T.J. Host effects and sequences essential for accumulation of defective interfering RNAs of cucumber necrosis and tomato bushy stunt tombusviruses. Virology 1995, 210, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.; Scholthof, H.B.; Jackson, A.O. The effect of defective interfering RNAs on the accumulation of tomato bushy stunt virus proteins and implications for disease attenuation. Virology 1995, 211, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Hannon, G.J. RNA interference. Nature 2002, 418, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Post-transcriptional RNA silencing in plant-microbe interactions: a touch of robustness and versatility. Curr. Opin. Plant Biol. 2008, 11, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, D.; Molnar, A.; Lucioli, A.; Szittya, G.; Hornyik, C.; Tavazza, M.; Burgyan, J. A viral protein suppresses RNA silencing and binds silencing-generated, 21- to 25-nucleotide double-stranded RNAs. Embo J. 2002, 21, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Havelda, Z.; Hornyik, C.; Valoczi, A.; Burgyan, J. Defective interfering RNA hinders the activity of a tombusvirus-encoded posttranscriptional gene silencing suppressor. J. Virol. 2005, 79, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Molnar, A.; Silhavy, D.; Hornyik, C.; Burgyan, J. Short defective interfering RNAs of tombusviruses are not targeted but trigger post-transcriptional gene silencing against their helper virus. Plant Cell 2002, 14, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Hornyik, C.; Havelda, Z.; Burgyan, J. Identification of sequence elements of tombusvirus-associated defective interfering RNAs required for symptom modulation. Arch. Virol. 2006, 151, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Hou, H.; Qiu, W. Integrity of nonviral fragments in recombinant Tomato bushy stunt virus and defective interfering RNA is influenced by silencing and the type of inserts. Mol. Plant Microbe Interact. 2005, 18, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Viral vectors for the expression of proteins in plants. Curr. Opin. Biotechnol. 2007, 18, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Winterhagen, P.; Dubois, C.; Sinn, M.; Wetzel, T.; Reustle, G.M. Gene silencing and virus resistance based on defective interfering constructs in transgenic Nicotiana benthamiana is not linked to accumulation of siRNA. Plant Physiol. Biochem. 2009. [Google Scholar] [CrossRef]
- Qiu, W.; Scholthof, H.B. Using vectors derived from tomato bushy stunt virus (TBSV) and TBSV defective interfering RNAs (DIs). Curr. Protoc. Microbiol. 2007, Chapter 16, Unit 16I 14. [Google Scholar] [PubMed]
- Dasgupta, R.; Cheng, L.L.; Bartholomay, L.C.; Christensen, B.M. Flock house virus replicates and expresses green fluorescent protein in mosquitoes. J. Gen. Virol. 2003, 84, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, C.; Pogany, J.; Bujarski, J.; Romero, J. Use of a defective RNA of broad bean mottle bromovirus for stable gene expression in legumes. Arch. Virol. 2008, 153, 1755–1758. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J. Multiple roles of viral replication proteins in plant RNA virus replication. Methods Mol. Biol. 2008, 451, 55–68. [Google Scholar] [PubMed]
- Pantaleo, V.; Rubino, L.; Russo, M. Replication of Carnation Italian ringspot virus defective interfering RNA in Saccharomyces cerevisiae. J. Virol. 2003, 77, 2116–2123. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Pogany, J.; Panavas, T.; Xu, K.; Esposito, A.M.; Kinzy, T.G.; Nagy, P.D. Translation elongation factor 1A is a component of the tombusvirus replicase complex and affects the stability of the p33 replication co-factor. Virology 2009, 385, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Barajas, D.; Panavas, T.; Herbst, D.A.; Nagy, P.D. Cdc34p Ubiquitin-Conjugating Enzyme Is a Component of the Tombusvirus Replicase Complex and Ubiquitinates p33 Replication Protein. J. Virol. 2008, 82, 6911–6926. [Google Scholar] [CrossRef] [PubMed]
- Serva, S.; Nagy, P.D. Proteomics analysis of the tombusvirus replicase: Hsp70 molecular chaperone is associated with the replicase and enhances viral RNA replication. J. Virol. 2006, 80, 2162–2169. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Serviene, E.; Gal, J.; Panavas, T.; Nagy, P.D. Identification of essential host factors affecting tombusvirus RNA replication based on the yeast Tet promoters Hughes Collection. J. Virol. 2006, 80, 7394–7404. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Serviene, E.; Brasher, J.; Nagy, P.D. Yeast genome-wide screen reveals dissimilar sets of host genes affecting replication of RNA viruses. Proc. Natl. Acad. Sci. U S A 2005, 102, 7326–7331. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J. Yeast as a model host to dissect functions of viral and host factors in tombusvirus replication. Virology 2006, 344, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D. Yeast as a model host to explore plant virus-host interactions. Annu. Rev. Phytopathol. 2008, 46, 217–242. [Google Scholar] [CrossRef] [PubMed]
- Monkewich, S.; Lin, H.X.; Fabian, M.R.; Xu, W.; Na, H.; Ray, D.; Chernysheva, O.A.; Nagy, P.D.; White, K.A. The p92 polymerase coding region contains an internal RNA element required at an early step in Tombusvirus genome replication. J. Virol. 2005, 79, 4848–4858. [Google Scholar] [CrossRef] [PubMed]
- Pogany, J.; Stork, J.; Li, Z.; Nagy, P.D. In vitro assembly of the Tomato bushy stunt virus replicase requires the host Heat shock protein 70. Proc. Natl. Acad. Sci. U S A 2008, 105, 19956–19961. [Google Scholar] [CrossRef] [PubMed]
- Pogany, J.; Fabian, M.R.; White, K.A.; Nagy, P.D. A replication silencer element in a plus-strand RNA virus. Embo J. 2003, 22, 5602–5611. [Google Scholar] [CrossRef] [PubMed]
- Panaviene, Z.; Panavas, T.; Nagy, P.D. Role of an internal and two 3'-terminal RNA elements in assembly of tombusvirus replicase. J. Virol. 2005, 79, 10608–10618. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Hawkins, C.M.; Panaviene, Z.; Nagy, P.D. The role of the p33:p33/p92 interaction domain in RNA replication and intracellular localization of p33 and p92 proteins of Cucumber necrosis tombusvirus. Virology 2005, 338, 81–95. [Google Scholar] [CrossRef] [PubMed]
- McCartney, A.W.; Greenwood, J.S.; Fabian, M.R.; White, K.A.; Mullen, R.T. Localization of the tomato bushy stunt virus replication protein p33 reveals a peroxisome-to-endoplasmic reticulum sorting pathway. Plant Cell 2005, 17, 3513–3531. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Di Franco, A.; Martelli, G.P. Cytopathology in the identification and classification of tombusviruses. Intervirology 1987, 28, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.B.; Sasvari, Z.; Nagy, P.D. The host Pex19p plays a role in peroxisomal localization of tombusvirus replication proteins. Virology 2008, 379, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.; Stork, J.; Pogany, J.; Nagy, P.D. A temperature sensitive mutant of heat shock protein 70 reveals an essential role during the early steps of tombusvirus replication. Virology 2009. [Google Scholar] [CrossRef]
- Panaviene, Z.; Panavas, T.; Serva, S.; Nagy, P.D. Purification of the cucumber necrosis virus replicase from yeast cells: role of coexpressed viral RNA in stimulation of replicase activity. J. Virol. 2004, 78, 8254–8263. [Google Scholar] [CrossRef] [PubMed]
- Gursinsky, T.; Schulz, B.; Behrens, S.E. Replication of Tomato bushy stunt virus RNA in a plant in vitro system. Virology 2009, 390, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Pogany, J.; Na, H.; Nicholson, B.L.; Nagy, P.D.; White, K.A. A Discontinuous RNA Platform Mediates RNA Virus Replication: Building an Integrated Model for RNA-based Regulation of Viral Processes. PLoS Pathog. 2009, 5, e1000323. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.; Stork, J.; Nagy, P.D. A key role for heat shock protein 70 in the localization and insertion of tombusvirus replication proteins to intracellular membranes. J. Virol. 2009, 83, 3276–3287. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Pogany, J.; Nagy, P.D. Analysis of minimal promoter sequences for plus-strand synthesis by the Cucumber necrosis virus RNA-dependent RNA polymerase. Virology 2002, 296, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Panavas, T.; Nagy, P.D. Mechanism of stimulation of plus-strand synthesis by an RNA replication enhancer in a tombusvirus. J. Virol. 2005, 79, 9777–9785. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.; Nagy, P.D. Tomato bushy stunt virus co-opts the RNA-binding function of a host metabolic enzyme for viral genomic RNA synthesis. Cell Host Microbe 2008, 3, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Stork, J.; Panaviene, Z.; Nagy, P.D. Inhibition of in vitro RNA binding and replicase activity by phosphorylation of the p33 replication protein of Cucumber necrosis tombusvirus. Virology 2005, 343, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Shapka, N.; Stork, J.; Nagy, P.D. Phosphorylation of the p33 replication protein of Cucumber necrosis tombusvirus adjacent to the RNA binding site affects viral RNA replication. Virology 2005, 343, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Barajas, D.; Li, Z.; Nagy, P.D. The Nedd4-type Rsp5p ubiquitin ligase inhibits tombusvirus replication via regulating degradation of the p92 replication protein and decreasing the activity of the tombusvirus replicase. J. Virol. 2009. [Google Scholar] [CrossRef]
- Na, H.; White, K.A. Structure and prevalence of replication silencer-3' terminus RNA interactions in Tombusviridae. Virology 2006, 345, 305–316. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Pathak, K.B.; Nagy, P.D. Defective Interfering RNAs: Foes of Viruses and Friends of Virologists. Viruses 2009, 1, 895-919. https://doi.org/10.3390/v1030895
Pathak KB, Nagy PD. Defective Interfering RNAs: Foes of Viruses and Friends of Virologists. Viruses. 2009; 1(3):895-919. https://doi.org/10.3390/v1030895
Chicago/Turabian StylePathak, Kunj B., and Peter D. Nagy. 2009. "Defective Interfering RNAs: Foes of Viruses and Friends of Virologists" Viruses 1, no. 3: 895-919. https://doi.org/10.3390/v1030895
APA StylePathak, K. B., & Nagy, P. D. (2009). Defective Interfering RNAs: Foes of Viruses and Friends of Virologists. Viruses, 1(3), 895-919. https://doi.org/10.3390/v1030895