Impact of Two Reoviruses and Their Coinfection on the Rice RNAi System and vsiRNA Production


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Virus Materials
2.2. RNA Extraction and Virus Detection
2.3. Small RNA Sequencing and Analysis
2.4. Reverse Transcription Quantitative PCR
3. Results
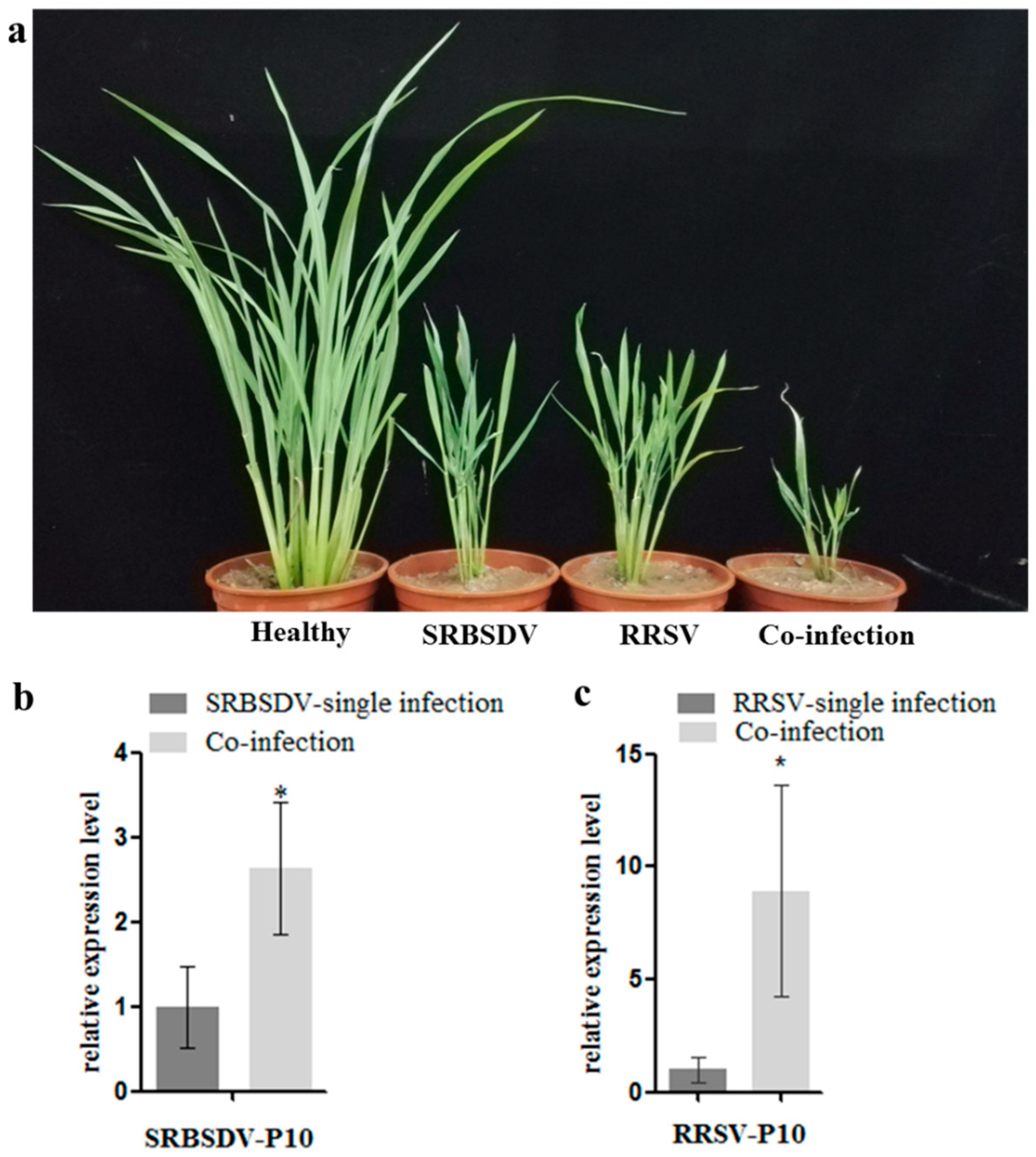
3.1. Co-Infection of SRBSDV and RRSV Exacerbated the Symptoms and Increased the Virus Titers
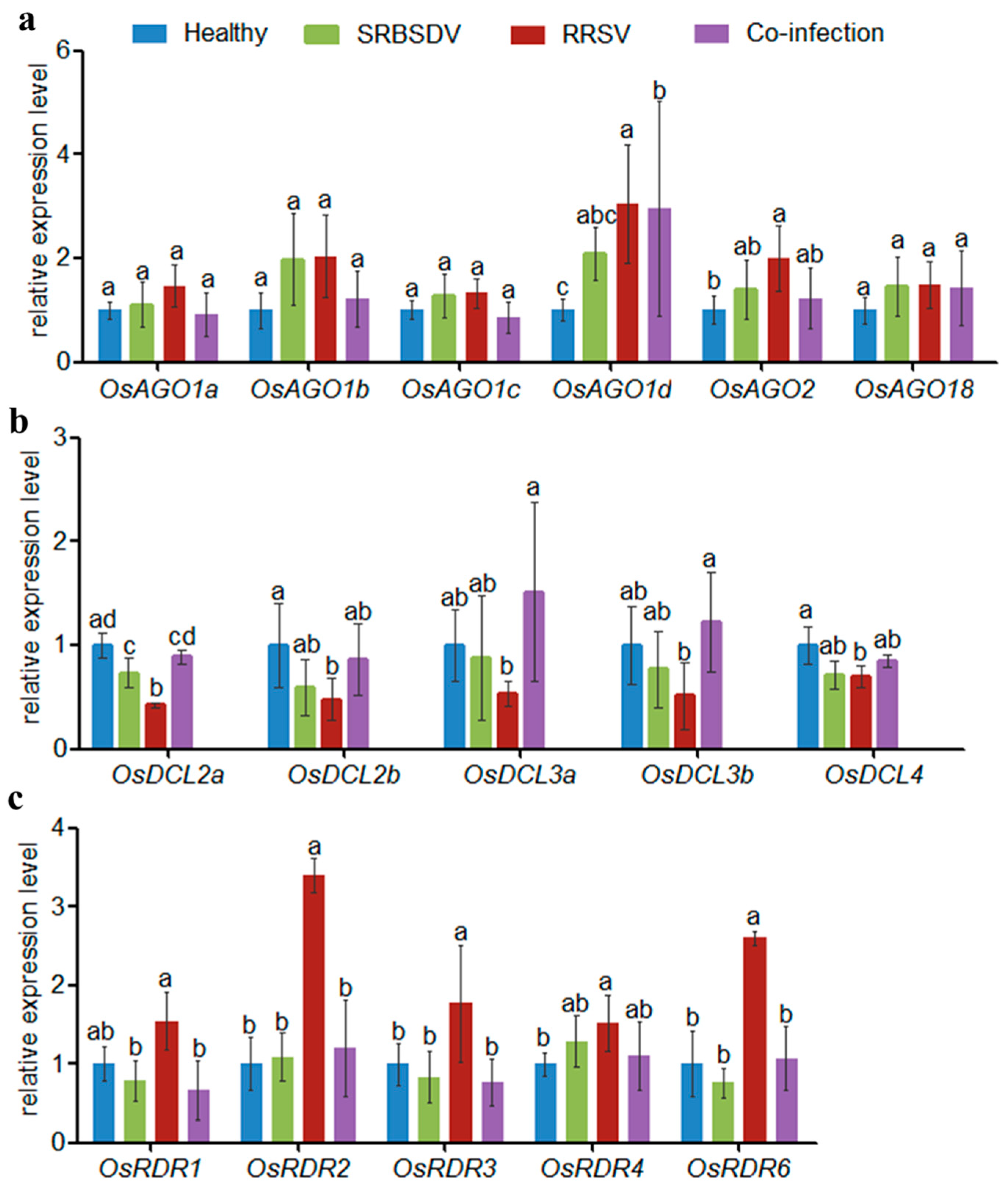
3.2. Co-Infection of SRBSDV and RRSV Altered Virus-Induced RNAi Pathway Genes
3.3. Co-Infection of SRBSDV and RRSV Altered Virus-Induced RNAi Pathway Genes
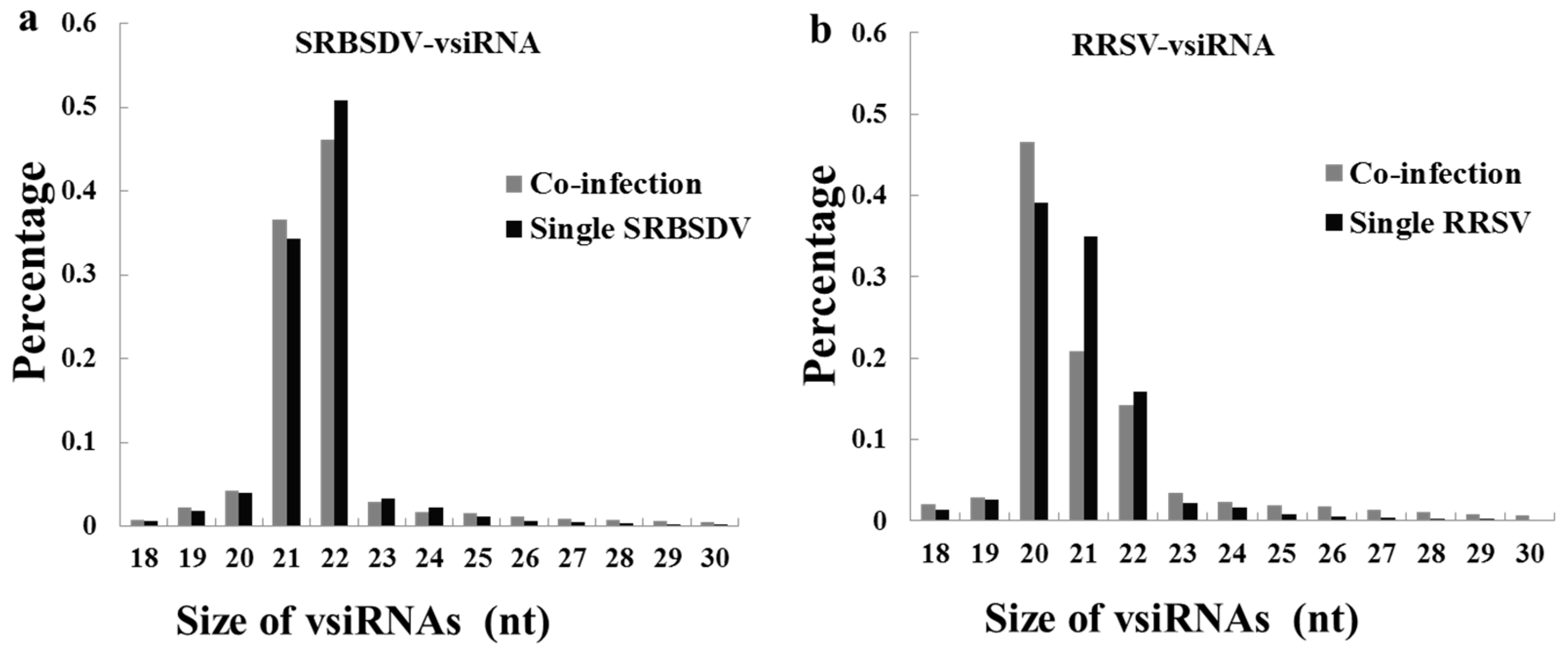
3.4. Co-Infection Changed the Length Distribution of the vsiRNAs of RRSV
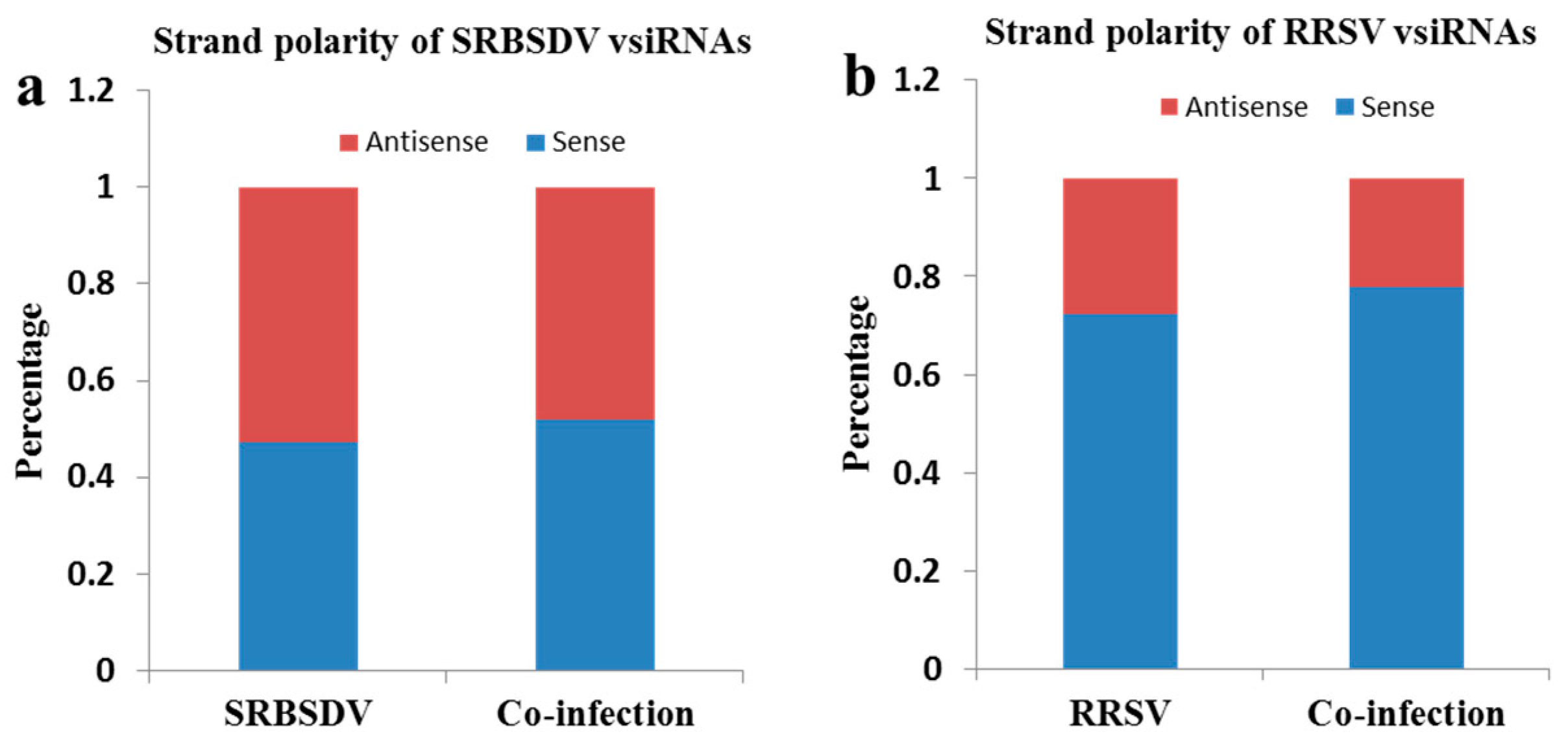
3.5. Co-Infection of SRBSDV and RRSV Had No Effect on the Strand Polarity of the vsiRNAs
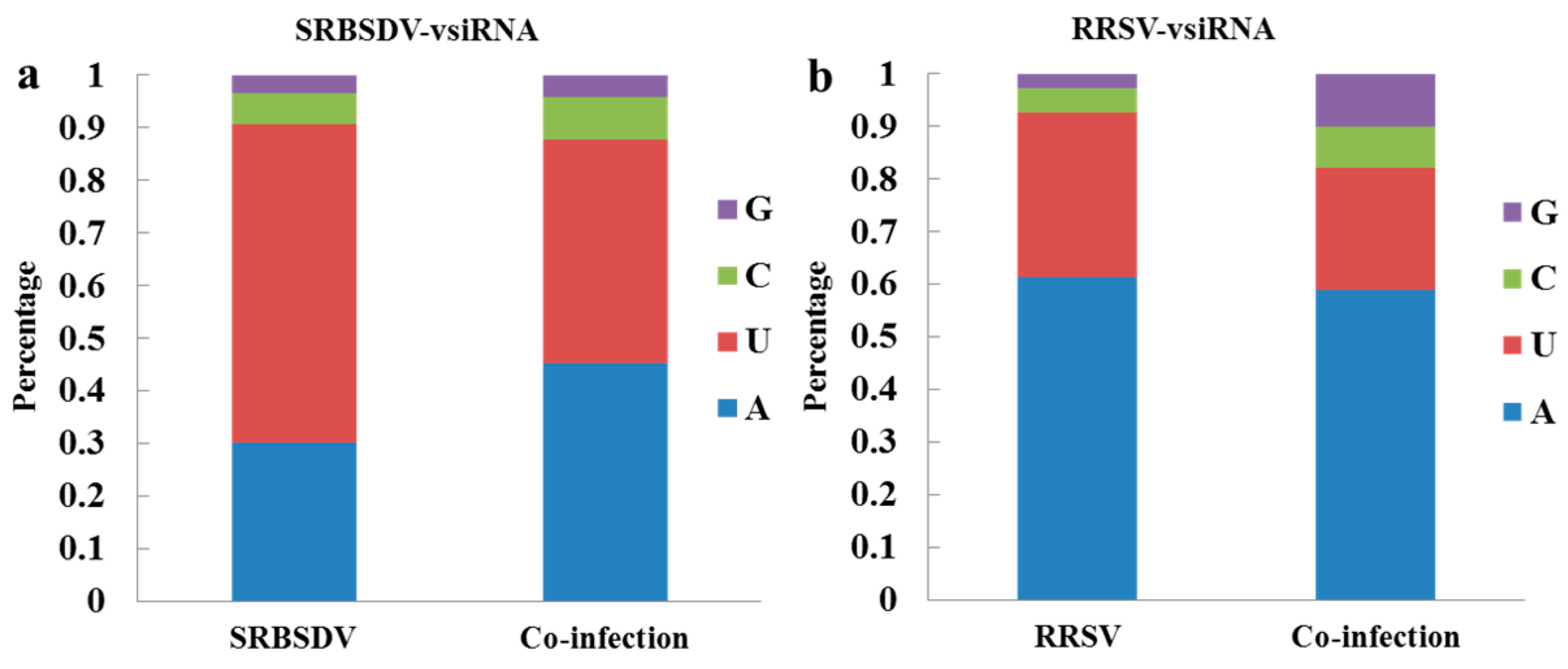
3.6. Co-Infection Changed the Bias of the 5′ Terminal Nucleotide of vsiRNAs
3.7. Co-Infection of SRBSDV and RRSV Had No Effect on vsiRNA Hotspots on the Viral Genome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buchon, N.; Vaury, C. RNAi: A defensive RNA-silencing against viruses and transposable elements. Heredity 2006, 96, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Morris, T.J. Suppressors of RNA silencing encoded by plant viruses and their role in viral infections. Febs. Lett. 2005, 579, 5958–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Z.; Zhao, Z.; Chen, L.; Li, M.; Zhou, T.; Deng, C.; Zhou, Q.; Fan, Z. Synergistic infection of two viruses MCMV and SCMV increases the accumulations of both MCMV and MCMV-derived siRNAs in maize. Sci. Rep. 2016, 6, 20520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Zhang, X.; Liu, S.; Wang, Y.; Li, D.W.; Yu, J.L.; Han, C.G. Synergistic infection of BrYV and PEMV 2 increases the accumulations of both BrYV and BrYV-derived siRNAs in Nicotiana benthamiana. Sci. Rep. 2017, 7, 45132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaucheret, H. Plant Argonautes. Trends Plant Sci. 2008, 13, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Bologna, N.G.; Voinnet, O. The diversity, biogenesis, and activities of endogenous silencing small RNAs in Arabidopsis. Annu. Rev. Plant Biol. 2014, 65, 473–503. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Maruyama, K.; Sun, L.; Kondo, H.; Tamada, T.; Suzuki, N. Different Dicer-like protein components required for intracellular and systemic antiviral silencing in Arabidopsis thaliana. Plant Signal Behav. 2015, 10, e1039214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blevins, T.; Rajeswaran, R.; Shivaprasad, P.V.; Beknazariants, D.; Si-Ammour, A.; Park, H.S.; Vazquez, F.; Robertson, D.; Meins, F., Jr.; Hohn, T.; et al. Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing. Nucleic. Acids. Res. 2006, 34, 6233–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 38, 14732–14737. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, J.; Garcia, D.; Pontier, D.; Ohnesorge, S.; Yu, A.; Garcia, S.; Braun, L.; Bergdoll, M.; Hakimi, M.A.; Lagrange, T.; et al. Argonaute quenching and global changes in Dicer homeostasis caused by a pathogen-encoded GW repeat protein. Gene Dev. 2010, 24, 904–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-Dependent RNA Polymerases and Dicer-Like Proteins in Antiviral Defense and Small Interfering RNA Biogenesis during Turnip Mosaic Virus Infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Du, P.; Wang, X.; Yu, Y.Q.; Qiu, Y.H.; Li, W.; Gal-On, A.; Zhou, C.; Li, Y.; Ding, S.W. Virus infection triggers widespread silencing of host genes by a distinct class of endogenous siRNAs inArabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14613–14618. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Li, F.; Li, W.; Ding, S. Suppression of Antiviral Silencing by Cucumber Mosaic Virus 2b Protein in Arabidopsis Is Associated with Drastically Reduced Accumulation of Three Classes of Viral Small Interfering RNAs. Plant Cell 2007, 19, 2053–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Qian, D.; Zheng, H.; Meng, L.Y.; Chen, J.; Le, W.J.; Zhou, T.; Zhou, Y.J.; Wei, C.H.; Li, Y. RNA-dependent RNA polymerase 6 of rice (Oryza sativa) plays role in host defense against negative-strand RNA virus, Rice stripe virus. Virus Res. 2012, 163, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Qian, D.; Sun, R.; Jiang, L.; Wang, Y.; Wei, C.; Zhang, Z.; Li, Y. OsRDR6 plays role in host defense against double-stranded RNA virus, Rice Dwarf Phytoreovirus. Sci. Rep. 2015, 5, 11324. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Carbonell, A.; Hoyer, J.S.; Fahlgren, N.; Gilbert, K.B.; Takeda, A.; Giampetruzzi, A.; Garcia Ruiz, M.T.; McGinn, M.G.; Lowery, N.; et al. Roles and Programming of Arabidopsis ARGONAUTE Proteins during Turnip Mosaic Virus Infection. Plos Pathog. 2015, 11, e1004755. [Google Scholar] [CrossRef] [PubMed]
- Dzianott, A.; Sztuba-Solińska, J.; Bujarski, J.J. Mutations in the Antiviral RNAi Defense Pathway Modify Brome mosaic virus RNA Recombinant Profiles. Mol. Plant Microbe Interact. 2012, 25, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.B.; Godon, C.; Mourrain, P.; Béclin, C.; Boutet, S.; Feuerbach, F.; Proux, F.; Vaucheret, H. Fertile hypomorphic ARGONAUTE (ago1) mutants impaired in post-transcriptional gene silencing and virus resistance. Plant Cell 2002, 14, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, C.; Moffett, P. Functional and Genetic Analysis Identify a Role for Arabidopsis ARGONAUTE5 in Antiviral RNA Silencing. Plant Cell 2015, 27, 1742–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, J.J.; Lewsey, M.G.; Patel, K.; Westwood, J.; Heimstädt, S.; Carr, J.P.; Baulcombe, D.C. An Antiviral Defense Role of AGO2 in Plants. PLoS ONE 2011, 6, e14639. [Google Scholar] [CrossRef] [PubMed]
- Jaubert, M.; Bhattacharjee, S.; Mello, A.F.S.; Perry, K.L.; Moffett, P. ARGONAUTE2 Mediates RNA-Silencing Antiviral Defenses against Potato virus X in Arabidopsis. Plant Physiol. 2011, 156, 1556–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamera, S.; Song, X.; Su, L.; Chen, X.; Fang, R. Cucumber mosaic virus suppressor 2b binds to AGO4-related small RNAs and impairs AGO4 activities. Plant J. 2012, 69, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Nonomura, K.I.; Morohoshi, A.; Nakano, M.; Eiguchi, M.; Miyao, A.; Hirochika, H.; Kurata, N. A Germ Cell Specific Gene of the ARGONAUTE Family Is Essential for the Progression of Premeiotic Mitosis and Meiosis during Sporogenesis in Rice. Plant Cell 2007, 19, 2583–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Yang, Z.; Wang, Y.; Zheng, L.; Ye, R.; Ji, Y.; Zhao, S.; Ji, S.; Liu, R.; Xu, L.; et al. Viral-inducible Argonaute18 confers broad-spectrum virus resistance in rice by sequestering a host microRNA. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitter, N.; Koundal, V.; Williams, S.; Pappu, H. Differential expression of tomato spotted wilt virus-derived viral small RNAs in infected commercial and experimental host plants. PLoS ONE 2013, 8, e76276. [Google Scholar] [CrossRef] [PubMed]
- Margaria, P.; Miozzi, L.; Ciuffo, M.; Rosa, C.; Axtell, M.J.; Pappu, H.R.; Turina, M. Comparison of small RNA profiles in Nicotiana benthamiana and Solanum lycopersicum infected by polygonum ringspot tospovirus reveals host-specific responses to viral infection. Virus Res. 2016, 211, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Li, Y.; E, Z.; Sun, F.; Du, L.; Xu, Q.; Zhou, T.; Zhou, Y.; Fan, Y. Identification of virus-derived siRNAs and their targets in RBSDV-infected rice by deep sequencing. J. Basic Microbiol. 2018, 58, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, Y.; Xia, Z.; Di, D.; Zhang, A.; Miao, H.; Zhou, T.; Fan, Z. Characterization of small interfering RNAs derived from Rice black streaked dwarf virus in infected maize plants by deep sequencing. Virus Res. 2017, 228, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Naveed, K.; Mitter, N.; Harper, A.; Dhingra, A.; Pappu, H.R. Comparative analysis of virus-specific small RNA profiles of three biologically distinct strains of Potato virus Y in infected potato (Solanum tuberosum) cv. Russet Burbank. Virus Res. 2014, 191, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Xu, D.; Xu, D.; Zhang, M. Southern rice black-streaked dwarf virus: A white-backed planthopper-transmitted fijivirus threatening rice production in Asia. Front. Microbiol. 2013, 4, 270. [Google Scholar] [CrossRef] [PubMed]
- Hibino, H.; Roechan, M.; Sudarisman, S.; Tantera, D.M. A virus disease of rice (“kerdil hampa”) transmitted by brown planthopper Nilaparvata lugens Stal in Indonesia. Contrib. Cent. Res. Inst. Agric. 1977, 35, 1–15. [Google Scholar]
- Shao, C.; Lü, H.; Wu, J.; Gong, Z. Nucleic Acid Binding Activity of Pns6 Encoded by Genome Segment 6 of Rice Ragged Stunt Oryzavirus. Acta Biochim. Biophys. Sin. 2004, 36, 457–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyaya, N.M.; Ramm, K.; Gellatly, J.A.; Li, Z.; Kositratana, W.; Waterhouse, P.M. Rice ragged stunt oryzavirus genome segments S7 and S10 encode non-structural proteins of M(r) 68,025 (Pns7) and M(r) 32,364 (Pns10). Arch. Virol. 1997, 142, 1719–1726. [Google Scholar] [CrossRef]
- Li, S.; Zhang, T.; Zhu, Y.; Zhou, G. Co-infection of two reoviruses increases both viruses accumulation in rice by up-regulating of viroplasm components and movement proteins bilaterally and RNA silencing suppressor unilaterally. Virol. J. 2017, 14, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, H.; Zhou, G. Synergism Between Southern rice black-streaked dwarf virus andRice ragged stunt virus Enhances Their Insect Vector Acquisition. Phytopathology 2014, 104, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhou, G. Characteristics of siRNAs derived from Southern rice black-streaked dwarf virus in infected rice and their potential role in host gene regulation. Virol. J. 2017, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lv, K.; Wang, M.; Zhou, G. Investigation of viruses infecting rice in southern China using a multiplex RT-PCR assay. Crop Prot. 2017, 91, 8–12. [Google Scholar] [CrossRef]
- Tatineni, S.; Alexander, J.; Gupta, A.K.; French, R. Asymmetry in Synergistic Interaction between Wheat streak mosaic virus and Triticum mosaic virus in Wheat. Mol. Plant Microbe Interact. 2018. [Google Scholar] [CrossRef] [PubMed]
- Szittya, G.; Moxon, S.; Pantaleo, V.; Toth, G.; Rusholme Pilcher, R.L.; Moulton, V.; Burgyan, J.; Dalmay, T. Structural and functional analysis of viral siRNAs. PLoS Pathog. 2010, 6, e1000838. [Google Scholar] [CrossRef] [PubMed]
- Molnar, A.; Csorba, T.; Lakatos, L.; Várallyay, E.; Lacomme, C.; Burgyán, J. Plant virus-derived small interfering RNAs originate predominantly from highly structured single-stranded viral RNAs. J. Virol. 2005, 79, 7812–7818. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Lu, R. Virus-derived siRNAs and piRNAs in immunity and pathogenesis. Curr. Opin. Virol. 2011, 1, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Huang, L.; Fu, S.; Wu, J.; Zhou, X. Population diversity of rice stripe virus-derived siRNAs in 474 three different hosts and RNAi-based antiviral immunity in Laodelphgax striatellus. PLoS ONE 2012, 7, e46238. [Google Scholar]
- Deleris, A.; Gallego-Bartolome, J.; Bao, J.; Kasschau, K.D.; Carrington, J.C.; Voinnet, O. Hierarchical action and inhibition of plant Dicer-like proteins in antiviral defense. Science 2017, 313, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Gu, L.; Song, X.; Cui, X.; Lu, Z.; Zhou, M.; Wang, L.; Hu, F.; Zhai, J.; Meyers, B.C.; et al. Dicer-like 3 produces transposable element-associated 24-nt siRNAs that control agricultural traits in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 3877–3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Li, P.; Zhai, J.; Zhou, M.; Ma, L.; Liu, B.; Jeong, D.; Nakano, M.; Cao, S.; Liu, C.; et al. Roles of DCL4 and DCL3b in rice phased small RNA 483 biogenesis. Plant J. 2012, 69, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Schuck, J.; Gursinsky, T.; Pantaleo, V.; Burgyan, J.; Behrens, S.E. AGO RISC mediated antiviral RNA silencing in a plant in vitro system. Nucleic. Acids. Res. 2013, 9, 5090–5103. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, T.A.; Howell, M.D.; Cuperus, J.T.; Li, D.; Hansen, J.E.; Alexander, A.L.; Chapman, E.J.; Fahlgren, N.; Allen, E.; Carrington, J.C. Specificity of ARGONAUTE7-miR390 interaction and dual functionality in TAS3 trans-acting siRNA formation. Cell 2008, 133, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Iwasaki, S.; Watanabe, T.; Utsumi, M.; Watanabe, Y. The Mechanism Selecting the Guide Strand from Small RNA Duplexes is Different among Argonaute Proteins. Plant Cell Physiol. 2008, 49, 493–500. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Reads | ||
|---|---|---|---|
| SRBSDV | RRSV | Co-Infection | |
| Total raw reads | 31,080,566 | 31,970,208 | 28,219,238 |
| Total clean reads | 27,486,888 | 27,299,736 | 24,177,550 |
| SRBSDV-derived siRNAs | 474,032 | - | 69,192 |
| RRSV-derived siRNAs | - | 1,110,229 | 346,402 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhang, T.; Huang, X.; Zhou, G. Impact of Two Reoviruses and Their Coinfection on the Rice RNAi System and vsiRNA Production. Viruses 2018, 10, 594. https://doi.org/10.3390/v10110594
Li Z, Zhang T, Huang X, Zhou G. Impact of Two Reoviruses and Their Coinfection on the Rice RNAi System and vsiRNA Production. Viruses. 2018; 10(11):594. https://doi.org/10.3390/v10110594
Chicago/Turabian StyleLi, Zhanbiao, Tong Zhang, Xiuqin Huang, and Guohui Zhou. 2018. "Impact of Two Reoviruses and Their Coinfection on the Rice RNAi System and vsiRNA Production" Viruses 10, no. 11: 594. https://doi.org/10.3390/v10110594
APA StyleLi, Z., Zhang, T., Huang, X., & Zhou, G. (2018). Impact of Two Reoviruses and Their Coinfection on the Rice RNAi System and vsiRNA Production. Viruses, 10(11), 594. https://doi.org/10.3390/v10110594