Site-Specific N-Glycosylation on the AAV8 Capsid Protein

 ,
,
Abstract
:1. Introduction
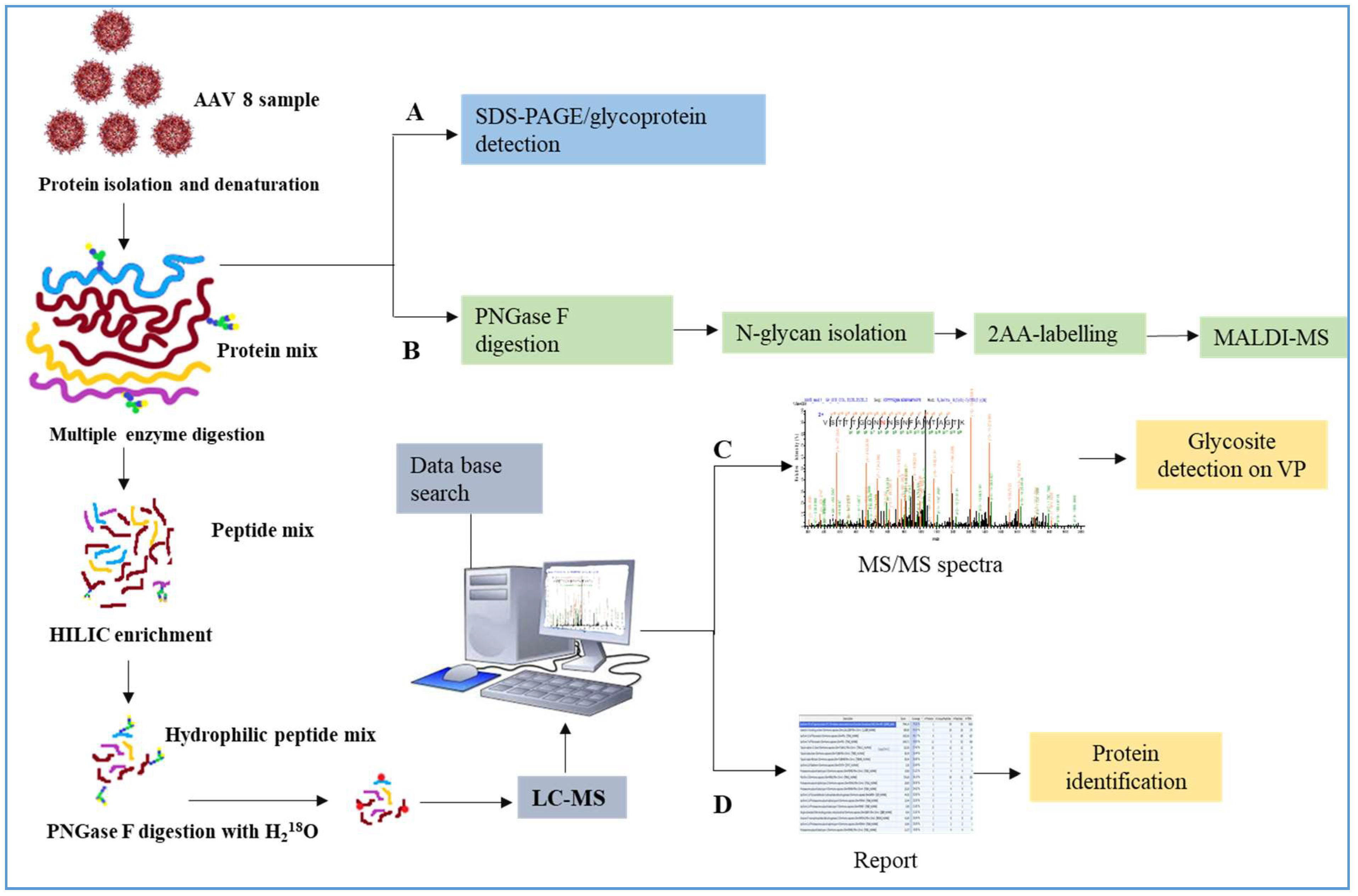
2. Materials and Methods
2.1. Source of Virus Sample
2.2. Viral Protein Sample Preparation
2.3. SDS–PAGE and Visualization of Glycoprotein Band and In-Gel-Digestion
2.4. N-Glycan Analysis
2.5. Peptide Mapping Analysis
2.6. Glycosylated Peptide Enrichment
2.7. N-Glycosite Detection (18O Labelling)
2.8. Peptide Analysis by LC-MS/MS
2.9. HCP Identification by LC-MS/MS Analysis
3. Results
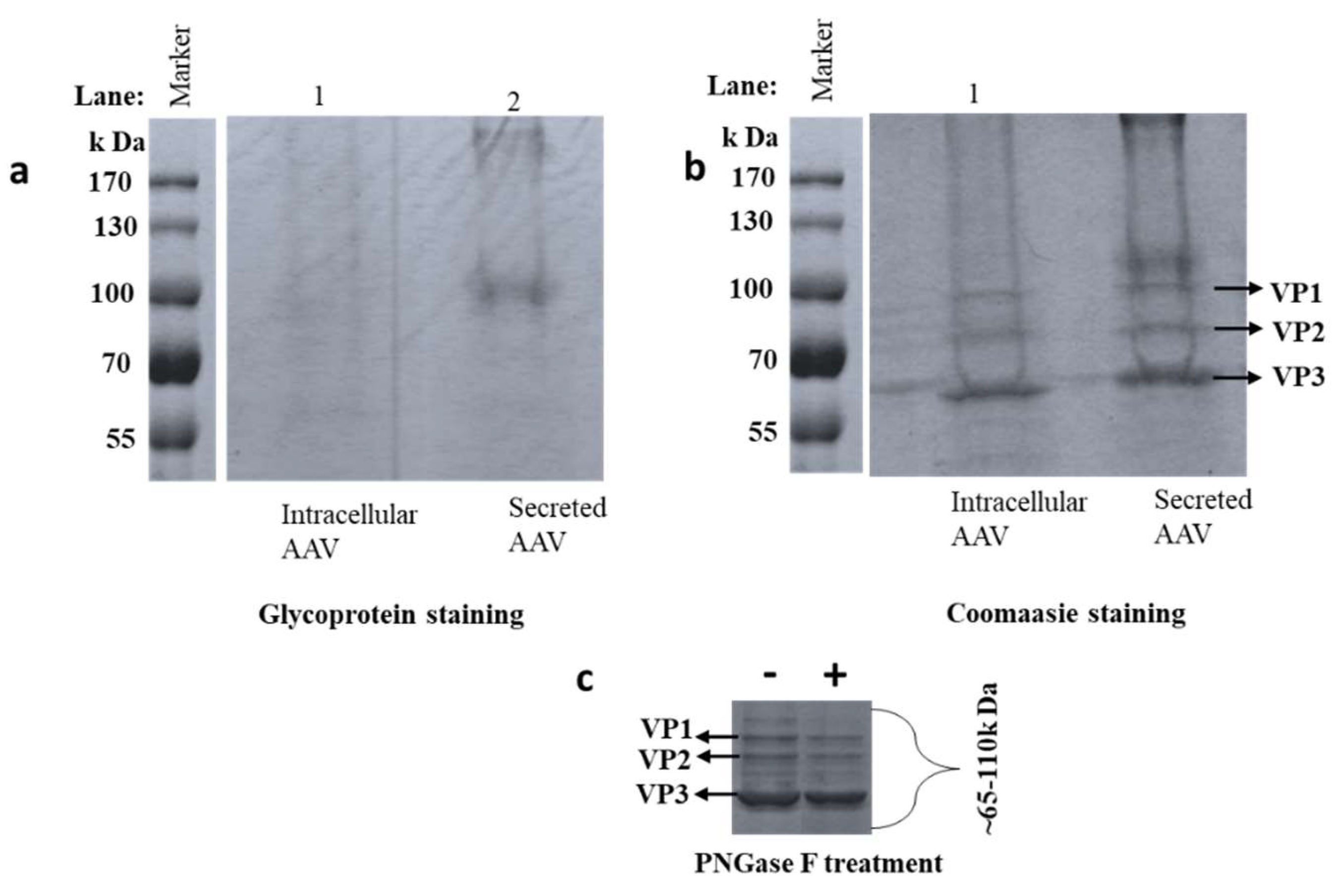
3.1. Detection of Glycosylation by SDS-PAGE
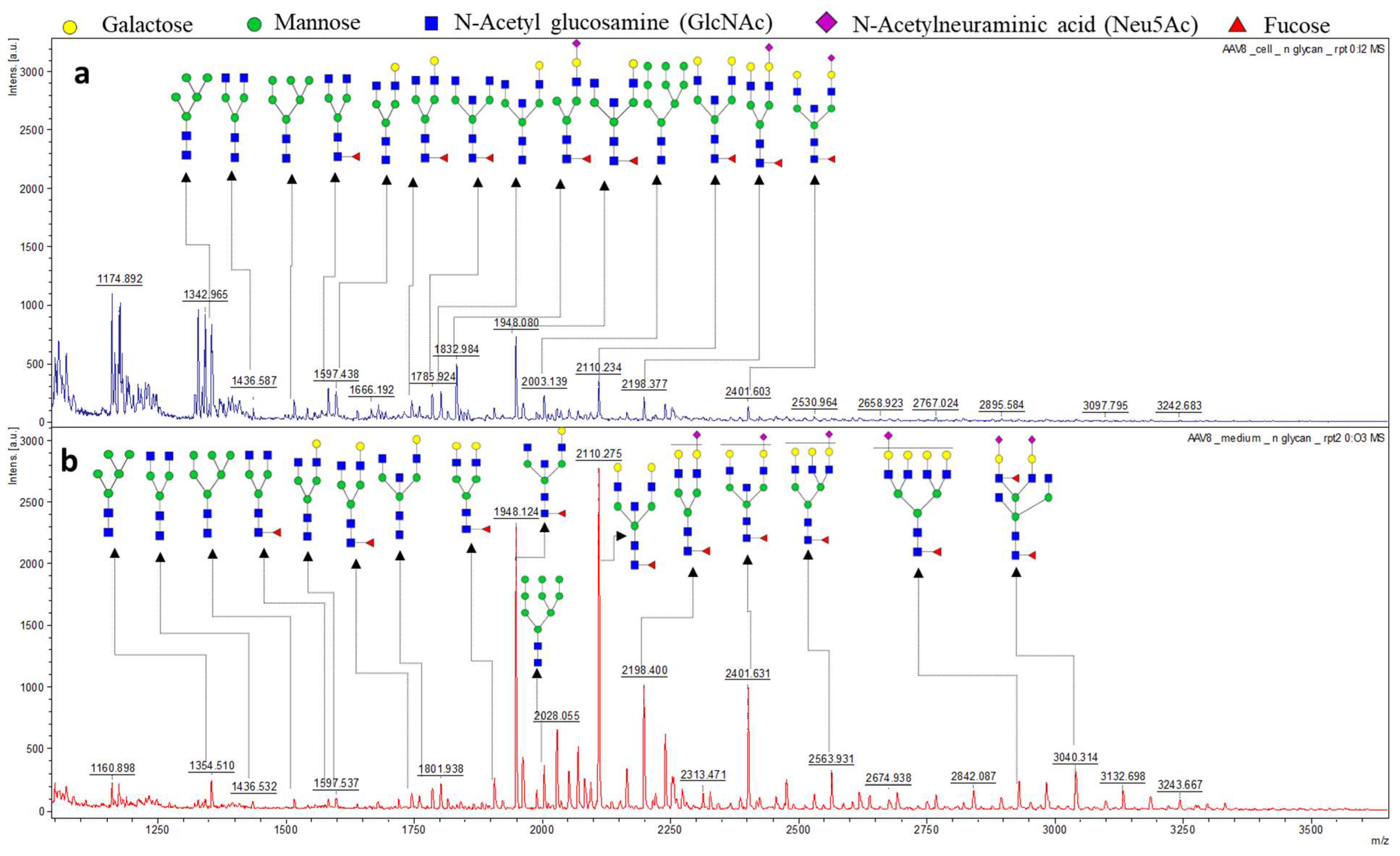
3.2. The N-Glycan Profile of Intracellular Derived AAV8 and Media Derived AAV8
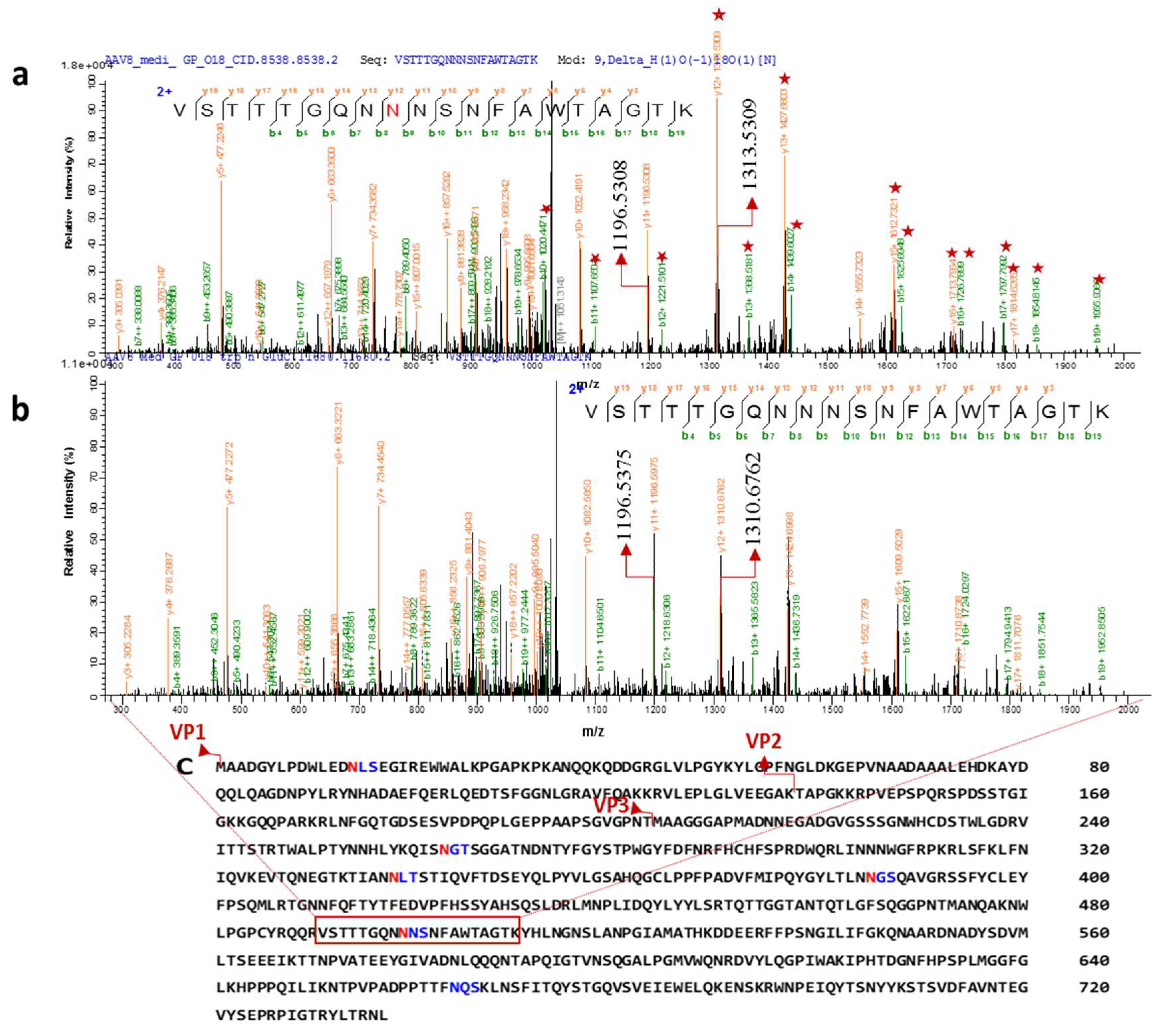
3.3. The Glycosylation Analysis at Peptide Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mueller, C.; Flotte, T.R. Clinical gene therapy using recombinant adeno-associated virus vectors. Gene Ther. 2008, 15, 858–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daya, S.; Berns, K.I. Gene therapy using adeno-associated virus vectors. Clin. Microbiol. Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S.; Zou, W.; Cheng, F.; Puschnik, A.S.; Meyer, N.L.; Ganaie, S.S.; Deng, X.; Wosen, J.E.; Davulcu, O.; Yan, Z.; et al. AAV serotypes have distinctive interactions with domains of the cellular receptor AAVR. J. Virol. 2017, 91, JVI.00391-17. [Google Scholar] [CrossRef] [PubMed]
- Earley, L.F.; Powers, J.M.; Adachi, K.; Baumgart, J.T.; Meyer, N.L.; Xie, Q.; Chapman, M.S.; Nakai, H. Adeno-Associated Virus Assembly-Activating Protein Is Not an Essential Requirement for Capsid Assembly of AAV Serotypes 4, 5 and 11. J. Virol. 2017, 91, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Vance, M.A.; Mitchell, A.; Samulski, R.J. AAV Biology, Infectivity and Therapeutic Use from Bench to Clinic. Gene Ther. Princ. Chall. 2015. [Google Scholar] [CrossRef] [Green Version]
- Sonntag, F.; Kother, K.; Schmidt, K.; Weghofer, M.; Raupp, C.; Nieto, K.; Kuck, A.; Gerlach, B.; Bottcher, B.; Muller, O.J.; et al. The Assembly-Activating Protein Promotes Capsid Assembly of Different Adeno-Associated Virus Serotypes. J. Virol. 2011, 85, 12686–12697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonntag, F.; Schmidt, K.; Kleinschmidt, J.A. A viral assembly factor promotes AAV2 capsid formation in the nucleolus. Proc. Natl. Acad. Sci. USA 2010, 107, 10220–10225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trempe, J.P.; Carter, B.J. Alternate mRNA Splicing Is Required for Synthesis of Adeno-Associated Virus VP1 Capsid Protein. J. Virol. 1988, 62, 3356–3363. [Google Scholar] [PubMed]
- Nam, H.-J.; Lane, M.D.; Padron, E.; Gurda, B.; McKenna, R.; Kohlbrenner, E.; Aslanidi, G.; Byrne, B.; Muzyczka, N.; Zolotukhin, S.; et al. Structure of Adeno-Associated Virus Serotype 8, a Gene Therapy Vector. J. Virol. 2007, 81, 12260–12271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kronenberg, S.; Böttcher, B.; Von Der, C.W.; Bleker, S.; Kleinschmidt, J.A; Bo, B.; Lieth, C.W. Von Der A Conformational Change in the Adeno-Associated Virus Type 2 Capsid Leads to the Exposure of Hidden VP1 N Termini. J. Virol. 2005, 79, 5296–5303. [Google Scholar] [CrossRef] [PubMed]
- Mietzsch, M.; Broecker, F.; Reinhardt, A.; Seeberger, P.H.; Heilbronn, R.; Imperiale, M.J. Differential Adeno-Associated Virus Serotype-Specific Interaction Patterns with Synthetic Heparins and Other Glycans. J. Virol. 2014, 88, 2991–3003. [Google Scholar] [CrossRef] [PubMed]
- Burger, C.; Gorbatyuk, O.S.; Velardo, M.J.; Peden, C.S.; Williams, P.; Zolotukhin, S.; Reier, P.J.; Mandel, R.J.; Muzyczka, N. Recombinant AAV viral vectors pseudotyped with viral capsids from serotypes 1, 2, and 5 display differential efficiency and cell tropism after delivery to different regions of the central nervous system. Mol. Ther. 2004, 10, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Alvira, M.R.; Somanathan, S.; Lu, Y.; Vandenberghe, L.H.; Rux, J.J.; Calcedo, R.; Sanmiguel, J.; Abbas, Z.; Wilson, J.M. Adeno-associated viruses undergo substantial evolution in primates during natural infections. Proc. Natl. Acad. Sci. USA 2003, 100, 6081–6086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.; Vandenberghe, L.H.; Alvira, M.R.; Lu, Y.; Calcedo, R.; Zhou, X.; Wilson, J.M. Clades of Adeno-Associated Viruses Are Widely Disseminated in Human Tissues. J. Virol. 2004, 78, 6381–6388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rayaprolu, V.; Kruse, S.; Kant, R.; Venkatakrishnan, B.; Movahed, N.; Brooke, D.; Lins, B.; Bennett, A.; Potter, T.; McKenna, R.; et al. Comparative Analysis of Adeno-Associated Virus Capsid Stability and Dynamics. J. Virol. 2013, 87, 13150–13160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Zhang, H.; Franco, L.M.; Young, S.P.; Schneider, A.; Bird, A.; Amalfitano, A.; Chen, Y.T.; Koeberl, D.D. Efficacy of an adeno-associated virus 8-pseudotyped vector in glycogen storage disease type II. Mol. Ther. 2005, 11, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Akache, B.; Grimm, D.; Pandey, K.; Yant, S.R.; Xu, H.; Kay, M.A. The 37/67-Kilodalton Laminin Receptor Is a Receptor for Adeno-Associated Virus Serotypes 8, 2, 3, and 9. J. Virol. 2006, 80, 9831–9836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, R.; Tetreault, R.; Gao, G.; Wang, L.; Bell, P.; Chandler, R.; Wilson, J.M.; Kazazian, H.H. Total correction of hemophilia A mice with canine FVIII using an AAV 8 serotype. Blood 2004, 103, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Vandendriessche, T.; Thorrez, L.; Acosta-Sanchez, A.; Petrus, I.; Wang, L.; Ma, L.; De Waele, L.; Iwasaki, Y.; Gillijns, V.; Wilson, J.M.; et al. Efficacy and safety of adeno-associated viral vectors based on serotype 8 and 9 vs. lentiviral vectors for hemophilia B gene therapy. J. Thromb. Haemost. 2007, 5, 16–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nienhuis, A.W.; Nathwani, A.C.; Davidoff, A.M. Gene Therapy for Hemophilia. Mol. Ther. 2017, 25, 1163–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chir, B.; Chowdary, P.; Riddell, A.; Sc, B.; Pie, A.J.; Davidoff, A.M. Adenovirus-associated virus vector-mediated gene transfer in hemophilia B. N. Engl. J. Med. 2012, 365, 2357–2365. [Google Scholar] [CrossRef]
- Nathwani, A.C. Long-Term Safety and Efficacy of Factor IX Gene Therapy in Hemophilia B. N. Engl. J. Med. 2015, 374, 1994–2004. [Google Scholar] [CrossRef] [PubMed]
- Lebherz, C.; Gao, G.; Louboutin, J.-P.; Millar, J.; Rader, D.; Wilson, J.M. Gene therapy with novel adeno-associated virus vectors substantially diminishes atherosclerosis in a murine model of familial hypercholesterolemia. J. Gene Med. 2004, 6, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhu, T.; Qiao, C.; Zhou, L.; Wang, B.; Zhang, J.; Chen, C.; Li, J.; Xiao, X. Adeno-associated virus serotype 8 efficiently delivers genes to muscle and heart. Nat. Biotechnol. 2005, 23, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Kaludov, N.; Brown, K.E.; Walters, R.W.; Zabner, J.; Chiorini, J.A. Adeno-associated virus serotype 4 (AAV4) and AAV5 both require sialic acid binding for hemagglutination and efficient transduction but differ in sialic acid linkage specificity. J. Virol. 2001, 75, 6884–6893. [Google Scholar] [CrossRef] [PubMed]
- Summerford, C.; Samulski, R.J. Membrane-associated heparan sulfate proteoglycan is a receptor for adeno-associated virus type 2 virions. J. Virol. 1998, 72, 1438–1445. [Google Scholar] [PubMed]
- Shen, S.; Bryant, K.D.; Brown, S.M.; Randell, S.H.; Asokan, A. Terminal N-linked galactose is the primary receptor for adeno-associated virus. J. Biol. Chem. 2011, 286, 13532–13540. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.; Zhou, Y.; Torian, U.; Nguyen, H.; Claire, M.S.; Yu, C.; Purcell, R.H.; Emerson, S.U.; Irol, J.V. Mutations within Potential Glycosylation Sites in the Capsid Protein of Hepatitis E Virus Prevent the Formation of Infectious Virus Particles. J. Virol. 2008, 82, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Cauet, G.; Strub, J.; Leize, E.; Wagner, E.; Dorsselaer, A.; Lusky, M. Identification of the Glycosylation Site of the Adenovirus Type 5 Fiber Protein. Biochemistry 2005, 44, 5453–5460. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Li, B.; Mah, C.S.; Govindasamy, L.; Agbandje-McKenna, M.; Cooper, M.; Herzog, R.W.; Zolotukhin, I.; Warrington, K.H.; Weigel-Van Aken, K.A.; et al. Next generation of adeno-associated virus 2 vectors: Point mutations in tyrosines lead to high-efficiency transduction at lower doses. Proc. Natl. Acad. Sci. USA. 2008, 105, 7827–7832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieger, J.C.; Choi, V.W.; Samulski, R.J. Production and characterization of adeno-associated viral vectors. Nat. Protoc. 2006, 1, 1412–1428. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Olsen, J.V.; Mann, M. Nature In-gel Digestion Protocol. Ref. Nat. Protoc. 2007, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, L.; Nass, S.; O’Riordan, C.; Pastor, E.; Zhang, X.K. Direct Liquid Chromatography/Mass Spectrometry Analysis for Complete Characterization of Recombinant Adeno-Associated Virus Capsid Proteins. Hum. Gene Ther. Methods 2017, 28, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Anumula, K.R. Advances in fluorescence derivatization methods for high-performance liquid chromatographic analysis of glycoprotein carbohydrates. Anal. Biochem. 2006, 1, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Geyer, H.; Geyer, R. Strategies for analysis of glycoprotein glycosylation. Biochim. Biophys. Acta-Proteins Proteom. 2006, 1764, 1853–1869. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Zhao, X.; Han, H.; Tong, W.; Zhang, Q.; Qin, P.; Chang, C.; Peng, B.; Ying, W.; Qian, X. N-linked glycoproteome profiling of human serum using tandem enrichment and multiple fraction concatenation. Electrophoresis 2013, 34, 2440–2450. [Google Scholar] [CrossRef] [PubMed]
- Gashash, E.A.; Aloor, A.; Li, D.; Zhu, H.; Xu, X.Q.; Xiao, C.; Zhang, J.; Parameswaran, A.; Song, J.; Ma, C.; Xiao, W.; Wang, P.G. An Insight into Glyco-Microheterogeneity of Plasma von Willebrand Factor by Mass Spectrometry. J. Proteome Res. 2017, 16, 3348–3362. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Li, D.Q.; Fu, Y.; Wang, H.P.; Zhang, J.F.; Yuan, Z.F.; Sun, R.X.; Zeng, R.; He, S.M.; Gao, W. pFind 2.0: A software package for peptide and protein identification via tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 2985–2991. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Lindskog, I.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Hochstrasser, D.F.; Appel, R.D. Detailed peptide characterization using PEPTIDEMASS—A World-Wide-Web-accessible tool. Electrophoresis 1997, 18, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.; Fung, K.; Wang, H.; Yen, C.; Cios, K. Identification of contaminants in proteomics mass spectrometry data. In Proceedings of the 2003 IEEE Bioinformatics Conference on Computational Systems Bioinformatics (CSB2003), Stanford, CA, USA, 11–14 August 2003; pp. 409–410. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, J.S. Protein identification using MS/MS data. J. Proteom. 2011, 74, 1842–1851. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Nilsson, C.L.; Hare, J.T.; Emmett, M.R.; Korostelev, A.; Ongley, H.; Marshall, A.G.; Chapman, M.S. Characterization of the Capsid Protein Glycosylation of Adeno-Associated Virus Type 2 by High-Resolution Mass Spectrometry. J. Virol. 2006, 80, 6171–6176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Horowitz, E.D.; Troupes, A.N.; Brown, S.M.; Pulicherla, N.; Samulski, R.J.; Agbandje-McKenna, M.; Asokan, A. Engraftment of a galactose receptor footprint onto adeno-associated viral capsids improves transduction efficiency. J. Biol. Chem. 2013, 288, 28814–28823. [Google Scholar] [CrossRef] [PubMed]
- Denard, J.; Beley, C.; Kotin, R.; Lai-Kuen, R.; Blot, S.; Leh, H.; Asokan, A.; Samulski, R.J.; Moullier, P.; Voit, T.; Garcia, L.; Svinartchouk, F. Human Galectin 3 Binding Protein Interacts with Recombinant Adeno-Associated Virus Type 6. J. Virol. 2012, 86, 6620–6631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afione, S.; DiMattia, M.A.; Halder, S.; Di Pasquale, G.; Agbandje-McKenna, M.; Chiorini, J.A. Identification and Mutagenesis of the Adeno-Associated Virus 5 Sialic Acid Binding Region. J. Virol. 2015, 89, 1660–1672. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
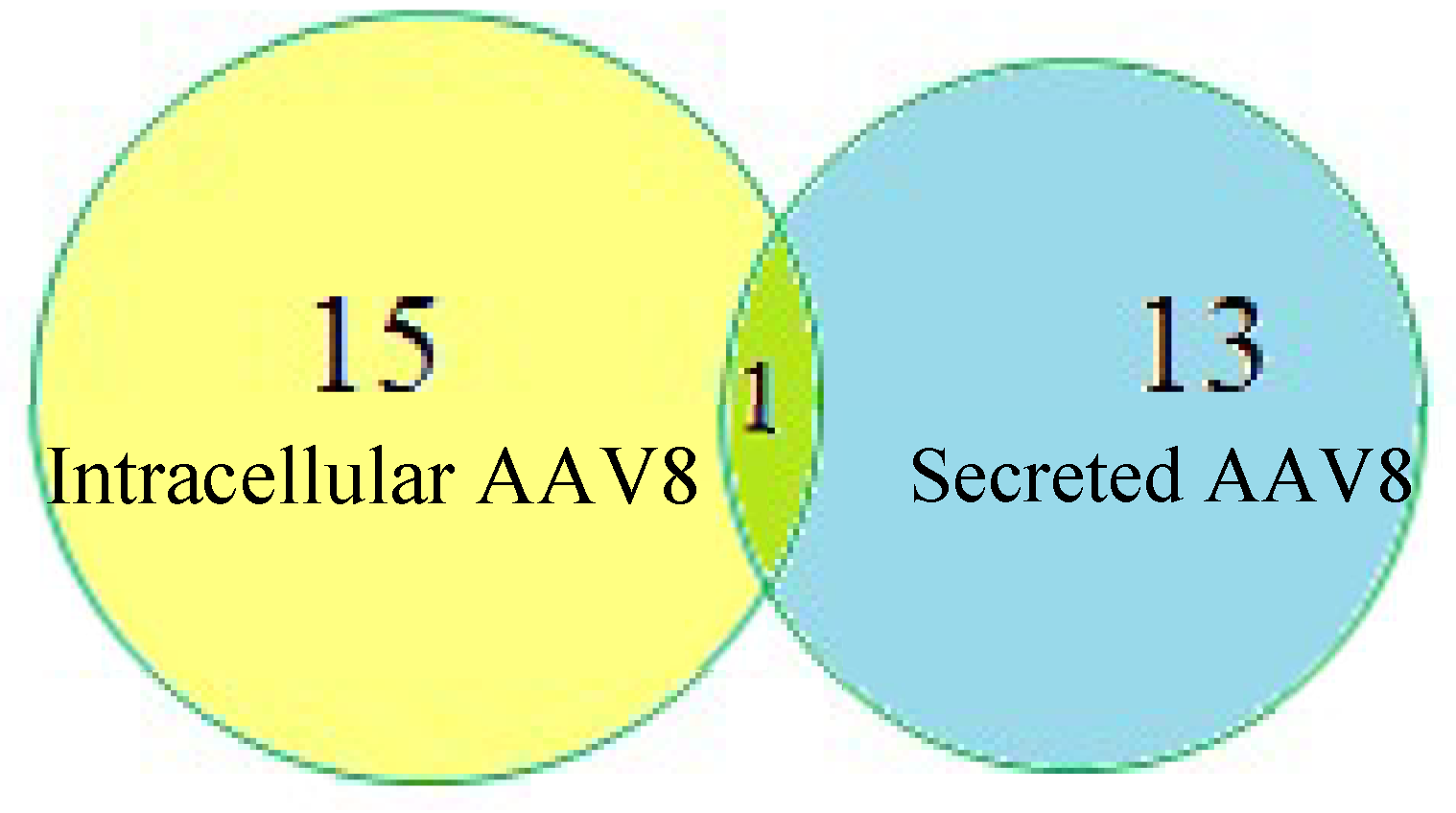{kind=link}
| UniProt Accession No. | Protein Identity | Coverage% | |
|---|---|---|---|
| Intracellular | Secreted | ||
| P0DP23 | Calmodulin 1 (Human) | 38.93 | ND |
| O60637-3 | Tetraspanin-3 (Human) | 34.39 | ND |
| P06748-3 | Nucleophosmin (Human) | 34.36 | ND |
| Q92542-2 | Nicastrin (Human) | 33.53 | ND |
| Q9BY67-2 | Cell adhesion molecule 1 (Human) | 33.33 | ND |
| P13473 | Lysosome-associated membrane glycoprotein 2 (Human) | 32.44 | ND |
| Q5ZPR3-3 | Isoform 3 of CD276 (Human) | 29.61 | ND |
| P11279 | Lysosome-associated membrane glycoprotein 1 (Human) | 29.5 | ND |
| Q14108 | Lysosome membrane protein 2 (Human) | 28.24 | ND |
| Q13740 | CD166 (Human) | 27.1 | ND |
| Q6PCB8-2 | Embigin (Human) | 25.27 | ND |
| Q08380 | Galectin-3-binding protein (Human) | 23.93 | 36.75 |
| P60900 | Proteasome subunit alpha type-6 (Human) | 22.76 | ND |
| P05556-2 | Isoform 2 of Integrin beta-1 (Human) | 22.18 | ND |
| Q13162 | Peroxiredoxin-4 (Human) | 21.03 | ND |
| P12268 | Inosine-5′-monophosphate dehydrogenase 2 (Human) | ND | 19.65 |
| P68363-2 | Tubulin alpha-1B chain | ND | 41.49 |
| P01834 | Ig kappa chain C region (Human) | ND | 35.85 |
| P02751-5 | Isoform 5 of Fibronectin (Human) | ND | 33.7 |
| P01619 | Ig kappa chain V-III region (Human) | ND | 31.19 |
| P23142-4 | Isoform C of Fibulin-1 | ND | 30.75 |
| P23142 | Fibulin-1 | ND | 30.3 |
| Q16222-2 | Isoform AGX1 of UDP-N-acetyl hexosamine pyro phosphorylase | ND | 29.31 |
| P07437 | Tubulin beta chain | ND | 26.35 |
| P0CG48 | Polyubiquitin-C | ND | 23.65 |
| Q04837 | Single-stranded DNA-binding protein, mitochondrial | ND | 22.3 |
| P35556 | Fibrillin-2 (Human) | ND | 20.81 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aloor, A.; Zhang, J.; Gashash, E.A.; Parameswaran, A.; Chrzanowski, M.; Ma, C.; Diao, Y.; Wang, P.G.; Xiao, W. Site-Specific N-Glycosylation on the AAV8 Capsid Protein. Viruses 2018, 10, 644. https://doi.org/10.3390/v10110644
Aloor A, Zhang J, Gashash EA, Parameswaran A, Chrzanowski M, Ma C, Diao Y, Wang PG, Xiao W. Site-Specific N-Glycosylation on the AAV8 Capsid Protein. Viruses. 2018; 10(11):644. https://doi.org/10.3390/v10110644
Chicago/Turabian StyleAloor, Arya, Junping Zhang, Ebtesam A. Gashash, Aishwarya Parameswaran, Matthew Chrzanowski, Cheng Ma, Yong Diao, Peng George Wang, and Weidong Xiao. 2018. "Site-Specific N-Glycosylation on the AAV8 Capsid Protein" Viruses 10, no. 11: 644. https://doi.org/10.3390/v10110644
APA StyleAloor, A., Zhang, J., Gashash, E. A., Parameswaran, A., Chrzanowski, M., Ma, C., Diao, Y., Wang, P. G., & Xiao, W. (2018). Site-Specific N-Glycosylation on the AAV8 Capsid Protein. Viruses, 10(11), 644. https://doi.org/10.3390/v10110644