Pathobiological and Genomic Characterization of a Cold-Adapted Infectious Bronchitis Virus (BP-caKII)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus, Egg and Chicken
2.2. Primers, RNA Extraction, Real-Time and Conventional RT-PCR, and Genome Analysis
2.3. Cold Adaptation of the Virus
2.4. Virus Titration, Embryo Pathogenicity and Embryo Mortality Rate by Virus
2.5. Growth Kinetics
2.6. Tissue Tropism and Persistence Test
2.7. Generation of a Premature Reproductive Tract Pathogenicity Model Using DES (Diethylstilbestrol)
2.8. Statistical Analysis
2.9. Ethical Statement
3. Results
3.1. Establishment and Growth Kinetics of a Cold-Adapted IBV, BP-caKII
3.2. Embryonic Pathogenicity of BP-caKII
3.3. Tissue Tropism and Persistence of BP-caKII
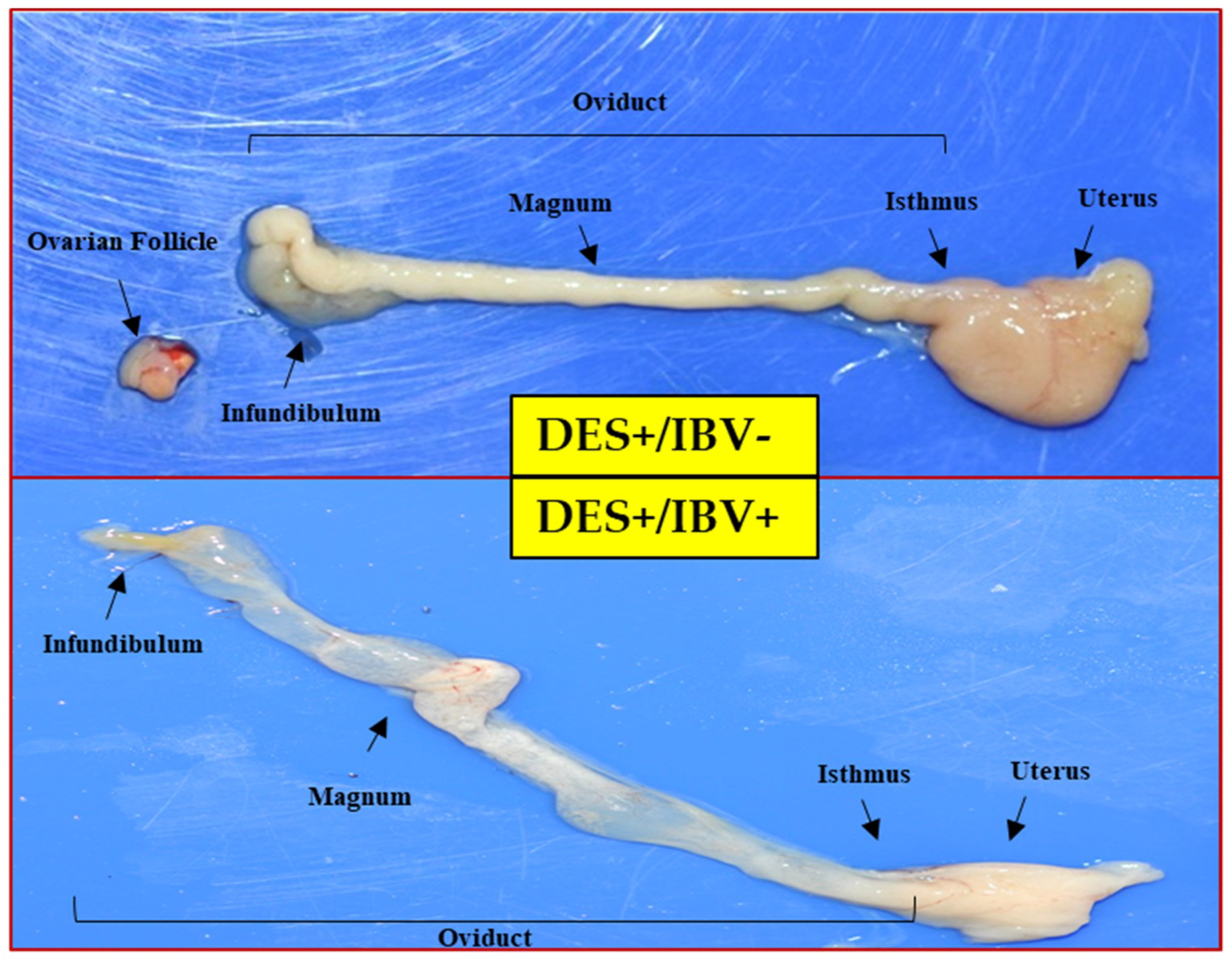
3.4. Pathogenicity of BP-caKII in the Premature Reproductive Tract Pathogenicity Model
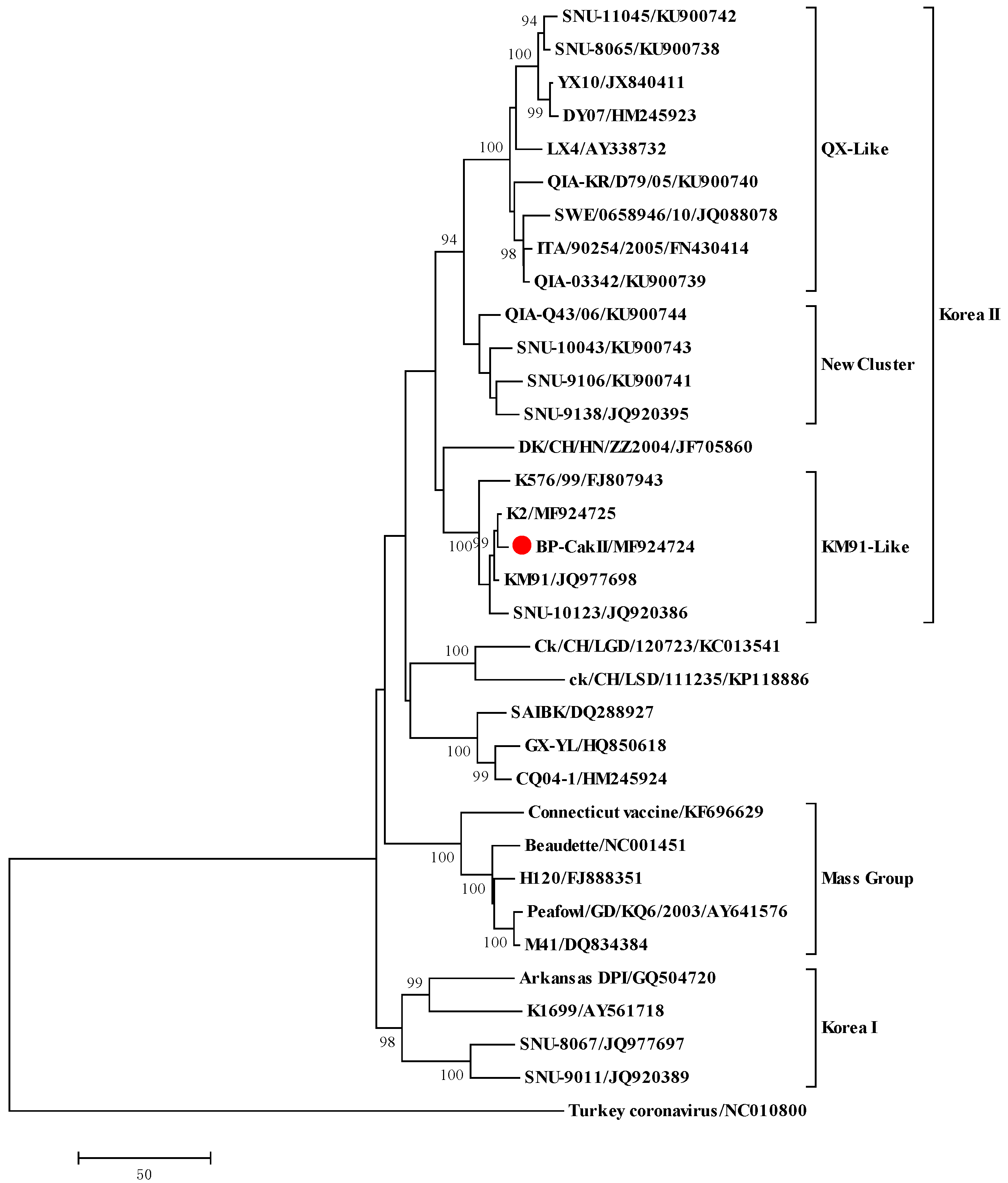
3.5. Genome Sequence Analysis of BP-caKII and K2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.C.; Ambali, A.G. Re-excretion of an enterotropic infectious bronchitis virus by hens at point of lay after experimental infection at day old. Vet. Rec. 1987, 120, 617–618. [Google Scholar] [CrossRef] [PubMed]
- Naqi, S.; Gay, K.; Patalla, P.; Mondal, S.; Liu, R. Establishment of persistent avian infectious bronchitis virus infection in antibody-free and antibody-positive chickens. Avian Dis. 2003, 47, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Armesto, M.; Cavanagh, D.; Britton, P. The replicase gene of avian coronavirus infectious bronchitis virus is a determinant of pathogenicity. PLoS ONE 2009, 4, e7384. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.G.; Shen, H.; Wang, J.; Tay, F.P.; Liu, D.X. Proteolytic processing of polyproteins 1a and 1ab between non-structural proteins 10 and 11/12 of Coronavirus infectious bronchitis virus is dispensable for viral replication in cultured cells. Virology 2008, 379, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.M.; Kwon, H.J.; Kim, I.H.; Mo, M.L.; Kim, J.H. Comparative genomics of Korean infectious bronchitis viruses (IBVs) and an animal model to evaluate pathogenicity of IBVs to the reproductive organs. Viruses 2012, 4, 2670–2683. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.K.; Jeon, W.J.; Lee, Y.J.; Jeong, O.M.; Choi, J.G.; Kwon, J.H.; Choi, K.S. Genetic diversity of avian infectious bronchitis virus isolates in Korea between 2003 and 2006. Avian Dis. 2008, 52, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Youn, H.N.; Kwon, J.S.; Lee, Y.J.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Characterization of a novel live attenuated infectious bronchitis virus vaccine candidate derived from a Korean nephropathogenic strain. Vaccine 2010, 28, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Belshe, R.B.; Mendelman, P.M.; Treanor, J.; King, J.; Gruber, W.C.; Piedra, P.; Bernstein, D.I.; Hayden, F.G.; Kotloff, K.; Zangwill, K.; et al. The efficacy of live attenuated, cold-adapted, trivalent, intranasal influenzavirus vaccine in children. N. Engl. J. Med. 1998, 338, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Maassab, H.F.; Bryant, M.L. The development of live attenuated cold-adapted influenza virus vaccine for humans. Rev. Med. Virol. 1999, 9, 237–244. [Google Scholar] [CrossRef]
- Gelb, J., Jr.; Lunt, R.L.; Metz, A.L.; Fries, P.A. Attenuation of avian infectious bronchitis virus by cold-adaptation. Avian Dis. 1991, 35, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Liu, D.X. Characterization of temperature-sensitive (ts) mutants of Coronavirus infectious bronchitis virus (IBV). Adv. Exp. Med. Biol. 2001, 494, 557–562. [Google Scholar] [PubMed]
- Kim, J.H. Serological Differentiation, Pathogenicity and Immunogenicity of Avian Infectious Bronchitis Viruses Isolated in Korea; Seoul National University: Seoul, Korea, 1995. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Alexander, D.J.; Allan, W.H. Newcastle disease virus pathotypes. Avian Pathol. 1974, 3, 269–278. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US). Methods for the Examination of Poultry Biologics; National Academies: Washington, DC, USA, 1963. [Google Scholar]
- Orsi, M.; Doretto Júnior, L.; Reischak, D.; Silva, L.D.; Spilki, F.; Buzinaro, M.; Arns, C. Newcastle disease virus vaccine strains: Immunogenicity is not influenced by ICPI. Braz. J. Poult. Sci. 2009, 11, 129–133. [Google Scholar] [CrossRef]
- Mo, M.L.; Hong, S.M.; Kwon, H.J.; Kim, I.H.; Song, C.S.; Kim, J.H. Genetic diversity of spike, 3a, 3b and e genes of infectious bronchitis viruses and emergence of new recombinants in Korea. Viruses 2013, 5, 550–567. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Kwon, H.J.; Choi, K.S.; Kim, J.H. Comparative genomics of QX-like infectious bronchitis virus in Korea. Arch. Virol. 2017, 162, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Bijlenga, G.; Cook, J.K.; Gelb, J., Jr.; de Wit, J.J. Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: A review. Avian Pathol. 2004, 33, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Xue, Y.; Wang, J.; Chen, W.; Chen, F.; Bi, Y.; Xie, Q. Development and efficacy of a novel live-attenuated QX-like nephropathogenic infectious bronchitis virus vaccine in China. Vaccine 2015, 33, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.H.; Kim, M.S.; Jang, J.H.; Lee, D.H.; Park, J.K.; Youn, H.N.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poult. Sci. 2012, 91, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Khanh, N.P.; Tan, S.W.; Yeap, S.K.; Lee, H.J.; Choi, K.S.; Hair-Bejo, M.; Bich, T.N.; Omar, A.R. Comparative pathogenicity of Malaysian QX-like and variant infectious bronchitis virus strains in chickens at different age of exposure to the viruses. J. Comp. Pathol. 2018, 161, 43–54. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, J.A.; Newbold, R.R.; Bullock, B.C. Long-term effects on the female mouse genital tract associated with prenatal exposure to diethylstilbestrol. Cancer Res. 1980, 40, 3988–3999. [Google Scholar] [PubMed]
- Hao, W.; Wojdyla, J.A.; Zhao, R.; Han, R.; Das, R.; Zlatev, I.; Manoharan, M.; Wang, M.; Cui, S. Crystal structure of Middle East respiratory syndrome coronavirus helicase. PLoS Pathog. 2017, 13, e1006474. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, K.A.; Thiel, V.; Dobbe, J.C.; van der Meer, Y.; Snijder, E.J.; Ziebuhr, J. Multiple enzymatic activities associated with severe acute respiratory syndrome coronavirus helicase. J. Virol. 2004, 78, 5619–5632. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, H.; Pan, J.; Xiang, N.; Tien, P.; Ahola, T.; Guo, D. Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase. Proc. Natl. Acad. Sci. USA 2009, 106, 3484–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minskaia, E.; Hertzig, T.; Gorbalenya, A.E.; Campanacci, V.; Cambillau, C.; Canard, B.; Ziebuhr, J. Discovery of an RNA virus 3'->5' exoribonuclease that is critically involved in coronavirus RNA synthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 5108–5113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snijder, E.J.; Decroly, E.; Ziebuhr, J. The Nonstructural Proteins Directing Coronavirus RNA Synthesis and Processing. Adv. Virus Res. 2016, 96, 59–126. [Google Scholar] [PubMed]
- Decroly, E.; Imbert, I.; Coutard, B.; Bouvet, M.; Selisko, B.; Alvarez, K.; Gorbalenya, A.E.; Snijder, E.J.; Canard, B. Coronavirus nonstructural protein 16 is a cap-0 binding enzyme possessing (nucleoside-2′O)-methyltransferase activity. J. Virol. 2008, 82, 8071–8084. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Hackbart, M.; Mettelman, R.C.; O'Brien, A.; Mielech, A.M.; Yi, G.; Kao, C.C.; Baker, S.C. Coronavirus nonstructural protein 15 mediates evasion of dsRNA sensors and limits apoptosis in macrophages. Proc. Natl. Acad. Sci. USA 2017, 114, E4251–E4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.E.; Jackwood, M.W.; McKinley, E.T.; Thor, S.W.; Hilt, D.A.; Acevedol, N.D.; Williams, S.M.; Kissinger, J.C.; Paterson, A.H.; Robertson, J.S.; et al. Changes in nonstructural protein 3 are associated with attenuation in avian coronavirus infectious bronchitis virus. Virus Genes 2012, 44, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Kusov, Y.; Hilgenfeld, R. Nsp3 of coronaviruses: Structures and functions of a large multi-domain protein. Antiviral Res. 2018, 149, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Zheng, Y.; Yang, Y.; Liu, C.; Geng, Q.; Luo, C.; Zhang, W.; Li, F. Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathog. 2018, 14, e1007009. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.J.; Kitame, F.; Kendal, A.P.; Maassab, H.F.; Naeve, C. Identification of sequence changes in the cold-adapted, live attenuated influenza vaccine strain, A/Ann Arbor/6/60 (H2N2). Virology 1988, 167, 554–567. [Google Scholar] [CrossRef]
- Hurst, K.R.; Koetzner, C.A.; Masters, P.S. Characterization of a critical interaction between the coronavirus nucleocapsid protein and nonstructural protein 3 of the viral replicase-transcriptase complex. J. Virol. 2013, 87, 9159–9172. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, H.; Fan, H.; Bowman, B.R.; Ooi, A.; Jayaram, J.; Collisson, E.W.; Lescar, J.; Prasad, B.V. X-ray structures of the N- and C-terminal domains of a coronavirus nucleocapsid protein: Implications for nucleocapsid formation. J. Virol. 2006, 80, 6612–6620. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Chang, C.K.; Chang, Y.W.; Sue, S.C.; Bai, H.I.; Riang, L.; Hsiao, C.D.; Huang, T.H. Structure of the SARS coronavirus nucleocapsid protein RNA-binding dimerization domain suggests a mechanism for helical packaging of viral RNA. J. Mol. Biol. 2007, 368, 1075–1086. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer | Amplicon (bp) | Forward (5′ to 3′) | Reverse (5′ to 3′) | Usage |
|---|---|---|---|---|
| Nsp 3 | 158 | CACCTACAAGTTAACACCTG | GTAGGAATGCGAAGAGACTTAC | Real-Time RT-PCR |
| GS1 | 1487 | ACTTAAGTGTGATATAAATATATATC | GTTTGGTCAAGCAGTGTTAGG | Sequencing |
| GS2 | 1580 | GTCAATTGTTGTCCTAGCAGC | CACATGTAGCTGGTTTAAC | |
| GS3 | 755 | GAATTCATGGAGACTTGCTCTTC | GTCTTGTATAAGAGCCAACAC | |
| GS4 | 471 | TTCTGATGTTCCTAGAGAAG | CGCCATCTACAAGAACATTC | |
| GS5 | 598 | CCGCTCTGTTGTTGTAAAAC | GGGCAATTTGAATATTGCGTC | |
| GS6 | 1629 | CTGGACTGGTTTGTTCAAAC | AAGACAATGGTCGCATAAGC | |
| GS7 | 1059 | GGAAGCATTGAAATGTGAAC | GGAATGTACCAAGGTTTTCGC | |
| GS8 | 760 | TCTCACTGCCTAAGTGGTTG | CCTTCTGTATATGCAGTAAG | |
| GS9 | 551 | CACATACCATCTTATGCTG | CATGCTACATTATCACCAC | |
| GS10 | 543 | CTTACAGTCTAAAGGGCATG | ATCAGGATCACATCCACTAGC | |
| GS11 | 501 | AGCGAGCCTTTGATGTATG | GGTTTCCGAACTCAATAGC | |
| GS12 | 1713 | CACTGCATGTTCTCATGCAGC | ACAACGCGTCATTATAGCATC | |
| GS13 | 2159 | CAATCCGGAATTGGAACAG | CTTAGCCTTAGTAATGCGAG | |
| GS14 | 1441 | GCTACCTAACACACTAAACAC | GCACTTTGGTAGTAGTACAC | |
| GS15 | 616 | TGGTCCTGTCTGTGATAAC | CGTAAGAATAGCACTCTGC | |
| GS16 | 363 | GGTAGTGGAAGACATGTTC | CCACCATTTTGACAACTCGTC |
| Virus | Titer (EID50/0.1 mL) | Hours Post-Infection | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 40 | 48 | 64 | 72 | 80 | 88 | 96 | 104 | 112 | 120 | 136 | 144 | Mortality | ALD a | MDT b | ||
| BP-caKII | 1 × 104.0 | - | 1 | - | - | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 100% | 1 × 104.0 | 106.4 ±29.7 * (hours) |
| 1 × 103.0 | 1 | - | - | 1 | 1 | 1 | 1 | 2 | - | 1 | - | - | 80% | |||
| 1 × 102.0 | - | - | - | 1 | - | - | 1 | - | - | - | - | - | 20% | |||
| K2 | 1 × 104.0 | 2 | 1 | 2 | 3 | 2 | - | - | - | - | - | - | - | 100% | 1 × 103.0 | 63.2 ±15.3 (hours) |
| 1 × 103.0 | 1 | - | - | 1 | 2 | 2 | 2 | - | - | 2 | - | 100% | ||||
| 1 × 102.0 | - | - | - | 2 | 1 | - | - | - | - | - | 30% | |||||
| Positive Rate of Virus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trachea | Kidney | Cecal Tonsils | ||||||||||
| 1 a | 2 | 3 | 4 | 1 | 2 | 3 | 4 | 1 | 2 | 3 | 4 | |
| Cont | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 | 0/5 |
| BP-caKII | 3/5 | 4/5 | 1/5 | 0/5 | 0/5 | 0/5 | 3/5 | 0/5 | 0/5 | 0/5 | 0/5 | 3/5 |
| K2 | 5/5 | 3/5 | 2/5 | 0/5 | 0/5 | 2/5 | 3/5 | 0/5 | 0/5 | 1/5 | 2/5 | 1/5 |
| KM91 | 4/5 | 5/5 | 4/5 | 0/5 | 5/5 * | 5/5 * | 4/5 | 0/5 | 4/5 * | 4/5 ** | 5/5 ** | 5/5 *** |
| Group | Frequency of Pathological Lesions | ||||
|---|---|---|---|---|---|
| Cyst Score a | Caseous Material in the Oviduct | Total Lesion Frequency (%) | |||
| + | ++ | +++ | |||
| DES.C | 0/7 | 0/7 | 0/7 | 0/7 | 0/7 (0) |
| BP-caKII | 0/11 | 0/11 | 0/11 | 0/11 | 0/11 (0) |
| K2 | 0/9 | 1/9 | 0/9 | 0/9 | 1/9 (11) |
| Km91 | 0/8 | 1/8 | 1/8 | 3/8 | 5/8 (64) b |
| Gene | Length of KM91/K2/BP-caKII | K2 | BP-caKII | ||
|---|---|---|---|---|---|
| Identity (%, nt/aa) | Missense Mutations | Identity (%, nt/aa) | Missense Mutations | ||
| Genome | 27,629 (38.12)/27,626 (38.19)/27,623 (38.13) | 99.2% | 99.1% | ||
| 5′-UTR | 99.6% | G270A (nt) | 99.4% | T4G, G7A, G270A | |
| 1a | 3952 | 99.8%/99.7% | Table 6 | 99.8%/99.6% | Table 6 |
| 1ab | 6630/6329 | 99.0%/99.0% | Table 6 | 98.9%/99.2% | Table 6 |
| S | 1163/1162/1162 | 99.6%/99.5% | S1:L2S, V21del, F23N, H24N, R118M, D406G, K488E S2: F697L, S883F | 99.4%/98.6% | S1: L2S, V21del, F23N, H24N, V66A, R118M, F273L, S365I, K488E, V500L, S2: F697L, I847F, S879T, S883F, S1006F |
| 3a | 48 | 100%/100% | 100%/100% | ||
| 3b | 62 | 100%/100% | 100%/100% | ||
| E | 109 | 99.6%/100% | 99.6%/99% | L27F | |
| M | 226 | 100%/100% | 99.7%/99.5% | L80F | |
| 4b(ORF X) | 94 | 99.6%/98.9% | H21R | 99.6%/98.9% | H21R |
| 4c | 56 | 100%/100% | 99.4%/98.2% | I36V | |
| 5a | 65 | 99.4%/98.4% | F7C | 99.4%/98.4% | F7C |
| 5b | 82 | 99.5%/98.7% | Q66R | 99.5%/98.7% | Q66R |
| N | 409 | 99.5%/99.7% | S109R | 99%/98.5% | NTD: S109R, K136T LKR: I204V CTD: V317L, T343S IDR: N407Y |
| 6b | 73 | 98.6%/97.2% | Y29H | 99%/98.6% | V500G |
| 3′-UTR | 27,324/27,327 | 98.5% | A89T, A189G, T206C, G275A | 98.5% | A189G, G193A, A246G, G270A |
| NSP (Size) | Putative Function b | Protease a | K2 | BP-caKII | ||
|---|---|---|---|---|---|---|
| Identity | Missense Mutation | Identity | Missense Mutation | |||
| 2 (673) | Unknown | PLP | 99.5% | A30V, K320T, H478Y | 99.5% | A30V, K320T, H478Y |
| 3 (1593/1592) | Papain-like viral protease | PLP | 99.8% | T189I(PL1), D1510A(YD) | 99.6% | T189I (PL1), P321L (PL1), L978F (BSM), V1167del (3Ecto) V1168I (3Ecto), D1510A(YD) |
| 4 (514) | Unknown | PLP/3CLpro | 99.6% | W16R, T50A | 99.4% | T50A, R417C, Q514H |
| 5 (307) | Coronavirus endopeptidase C30 | 3CLpro | 99.6% | T171P | 100% | |
| 6 (293) | Hydrophobic domain | 3CLpro | 100% | 100% | ||
| 7 (83) | nsp7 superfamily | 3CLpro | 98.7% | D43Y | 97.5% | S4I, D43Y |
| 8 (210) | nsp8 superfamily | 3CLpro | 99.5% | T155I | 99.5% | T155I |
| 9 (111) | nsp9 superfamily | 3CLpro | 100% | 100% | ||
| 10 (145) | nsp10 superfamily, RNA synthesis | 3CLpro | 100% | 100% | ||
| 11 (21) | RNA-dependent RNA polymerase | 3CLpro | 100% | 100% | ||
| 12 (918) | RNA-dependent RNA polymerase | 3CLpro | 99.7% | S567N, E832D | 99.4% | P214L, S567N, R626C, P812L, E832D |
| 13 (600) | Viral RNA helicase | 3CLpro | 99.1% | S51L, RecA2 domain: I474V, P555Q, S559N, G594D | 99.0% | S51L, RecA2 domain: I474V, T512I, P555Q, S559N, G594D |
| 14 (521) | nsp11 superfamily; exoribonucelase | 3CLpro | 98.4% | T207S, L252Q, N256D, D287G, V455I, S505T, Q507N, N516S | 98.6% | L252Q, N256D, D287G, V455I, S505T, Q507N, N516S |
| 15 (338) | Nidoviral uridylate-specific endoribonucelase | 3CLpro | 97.3% | I22M, I38V, V133I, N173S, D202E, V249A, I265M, I297S, K308R | 97.0% | I22M, I38V, V133I, N173S, D202E, V249A, I265M, I297S, K308R, S313L |
| 16 (275) | 23S rRNA methylase | 3CLpro | 97.8% | F110L, V143I, K160R, V173L, L209I, K270Q | 97.8% | F110L, V143I, K160R, V173L, L209I, K270Q |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.-M.; An, S.-H.; Lee, C.-Y.; Song, C.-S.; Choi, K.-S.; Kim, J.-H.; Kwon, H.-J. Pathobiological and Genomic Characterization of a Cold-Adapted Infectious Bronchitis Virus (BP-caKII). Viruses 2018, 10, 652. https://doi.org/10.3390/v10110652
Hong S-M, An S-H, Lee C-Y, Song C-S, Choi K-S, Kim J-H, Kwon H-J. Pathobiological and Genomic Characterization of a Cold-Adapted Infectious Bronchitis Virus (BP-caKII). Viruses. 2018; 10(11):652. https://doi.org/10.3390/v10110652
Chicago/Turabian StyleHong, Seung-Min, Se-Hee An, Chung-Young Lee, Chang-Seon Song, Kang-Seuk Choi, Jae-Hong Kim, and Hyuk-Joon Kwon. 2018. "Pathobiological and Genomic Characterization of a Cold-Adapted Infectious Bronchitis Virus (BP-caKII)" Viruses 10, no. 11: 652. https://doi.org/10.3390/v10110652
APA StyleHong, S. -M., An, S. -H., Lee, C. -Y., Song, C. -S., Choi, K. -S., Kim, J. -H., & Kwon, H. -J. (2018). Pathobiological and Genomic Characterization of a Cold-Adapted Infectious Bronchitis Virus (BP-caKII). Viruses, 10(11), 652. https://doi.org/10.3390/v10110652