High-Throughput Sequencing Reveals Cyclamen persicum Mill. as a Natural Host for Fig Mosaic Virus


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Source Material
2.2. Extraction of Double-Stranded RNAs, High-Throughput Sequencing, and Sequence Bioinformatics Analysis
2.3. Extraction of Total Nucleic Acids and PCR Detection of Cyclamen-Infecting Viruses and Phytoplasmas
2.4. Sequence Analyses
2.5. Grafting and Mechanical Transmission of FMV
3. Results and Discussion
3.1. HTS Data Analyses and Identification of FMV
3.2. RT-PCR Detection and Genetic Variability of FMV from Cyclamen Plants
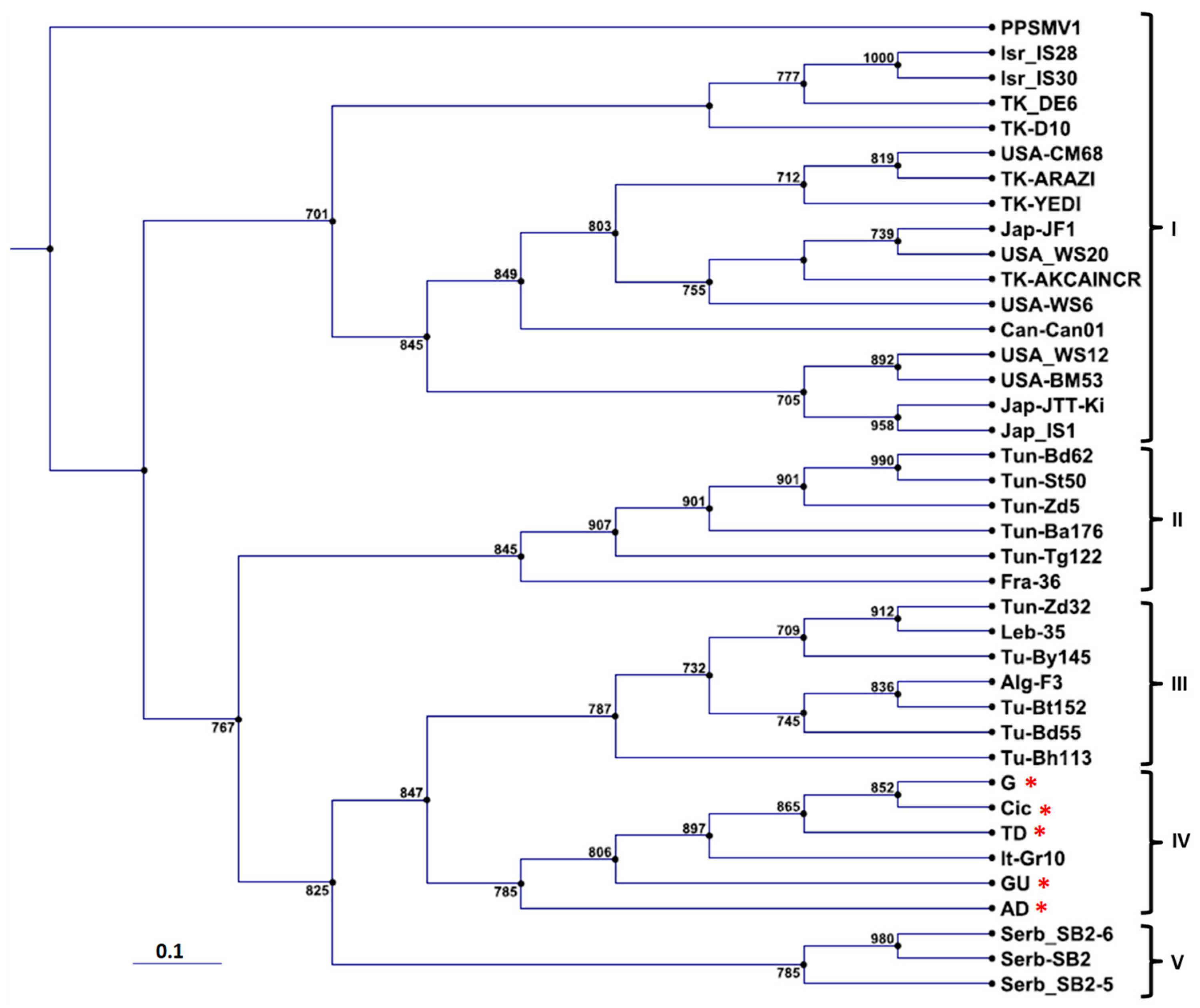
3.3. Phylogenetic Affinities of Cyclamen FMV Isolates
3.4. Etiology of FMV with Mosaic Disease on Cyclamen
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Elbeaino, T.; Digiaro, M.; Alabdullah, A.K.; De Stradis, A.; Minafra, A.; Mielke, N.; Castellano, M.A.; Martelli, G.P. A multipartite negative-sense single-stranded RNA virus is the putative agent of fig mosaic disease. J. Gen. Virol. 2009, 90, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Mielke-Ehret, N.; Muehlbach, H.P.; Martelli, G.P. ICTV Report Consortium. ICTV Virus Taxonomy Profile: Fimoviridae. J. Gen. Virol. 2018, 99, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Martelli, G.P. RNA-5 and -6, two additional negative-sense RNA segments associated with Fig mosaic virus. J. Plant Pathol. 2012, 94, 421–425. [Google Scholar]
- Elbeaino, T.; Digiaro, M.; Martelli, G.P. Complete nucleotides sequence of four viral RNAs segments of fig mosaic virus. Arch. Virol. 2009, 154, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Slykhuis, J.T. Viruses and mites. In Viruses and Invertebrates; Gibbs, A.J., Ed.; North Holland Publishing Co: Amsterdam, The Netherlands, 1973; pp. 391–405. [Google Scholar]
- Gentit, P.; Foissac, X.; Svanella-Dumas, L.; Peypelut, M.; Candresse, T. Characterization of two different Apricot latent virus variants associated with peach asteroid spot and peach sooty riHTSpot diseases. Arch. Virol. 2001, 146, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Candresse, T.; Faure, C.; Theil, S.; Marais, A. First report of Nectarine stem pitting-associated virus infecting Prunus mume in Japan. Plant Dis. 2017, 101, 393. [Google Scholar] [CrossRef]
- Mielke, N.; Muehlbach, H.P. A novel, multipartite, negative-strand RNA virus is associated with the riHTSpot disease of European mountain ash (Sorbus aucuparia L.). J. Gen. Virol. 2007, 88, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Foissac, X.; Svanella-Dumas, L.; Dulucq, M.J.; Candresse, T.; Gentit, P. Polyvalent detection of fruit tree tricho, capillo and foveavirus by nested RT-PCR using degenerated and inosine containing primers (DOP RT-PCR). Acta Horticul. 2001, 550, 37–44. [Google Scholar] [CrossRef]
- Mumford, R.A.; Barker, I.; Wood, K.R. An improved method for the detection of tospoviruses using the polymerase chain reaction. J. Virol. Methods 1996, 57, 109–115. [Google Scholar] [CrossRef]
- Mumford, R.A.; Barker, I.; Wood, K.R. The detection of Tomato spotted wilt virus using the polymerase chain reaction. J. Virol. Methods 1994, 46, 303–311. [Google Scholar] [CrossRef]
- Adkins, S.; Rosskopf, E.N. Key West nightshade, a new experimental host for plant viruses. Plant Dis. 2002, 86, 1310–1314. [Google Scholar] [CrossRef]
- Grieco, F.; Alkowni, R.; Saponari, M.; Savino, V.; Martelli, G.P. Molecular detection of olive viruses. EPPO Bull. 2000, 30, 469–473. [Google Scholar] [CrossRef]
- Peiman, M.; Xie, C. Sensitive detection of potato viruses, PVX, PLRV and PVS, by RT-PCR in potato leaf and tuber. Australas. Plant Dis. Notes 2006, 1, 41–46. [Google Scholar] [CrossRef]
- Riga, E.; Larsen, R.; Eastwell, K.; Guerra, N.; Guerra, L.; Crosslin, J.M. Rapid Detection of Tobacco rattle tobravirus in viruliferous paratrichodorus allius from greenhouse and field specimens. J. Nematol. 2009, 41, 60–63. [Google Scholar]
- Verma, N.; Kumar, K.; Kulshrestha, S.; Garg, I.D.; Raikhy, G.; Hallan, V.; Ram, R.; Zaidi, A.A. Detection and Molecular Characterization of a Tomato Aspermy Virus Isolate Infecting Chrysanthemums in India. Acta Hortic. 2006, 722, 41–54. [Google Scholar] [CrossRef]
- Davis, R.E.; Lee, L.M. Cluster-specific polymerase chain reaction amplification of 16s rDNA sequences for detection and identification of mycoplasmaslike organisms. Phytopathology 1993, 83, 100–1011. [Google Scholar] [CrossRef]
- Lee, I.M.; Gundersen, D.E.; Hammond, R.W.; Davis, R.E. Use of mycoplasma-like organism (MLO) group-specific oligonucleotide primers for nested-PCR assays to detect mixed- MLO infections in a single host plant. Phytopathology 1994, 84, 559–566. [Google Scholar] [CrossRef]
- Gundersen, D.E.; Lee, I.M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol. Mediterr. 1996, 35, 144–151. [Google Scholar]
- Ahrens, U.; Seemüller, E. Detection of DNA of plant pathogenic mycoplasma-like organisms by a polymerase chain reaction that amplifies a sequence of the 16S rRNA gene. Phytopathology 1992, 82, 828–832. [Google Scholar] [CrossRef]
- El Air, M.; Mahfoudhi, N.; Dhouibi, M.H.; Digiaro, M.; Elbeaino, T. Genetic variability of RNA-1, RNA-2 And RNA-3 of Fig mosaic virus isolates from Tunisia. J. Plant Pathol. 2016, 98, 295–299. [Google Scholar]



{kind=link}
{kind=link}
| FMV-Cic | 5′NCR % | CR % | 3′NCR % | Isolate (Accession Number) | |
|---|---|---|---|---|---|
| nt | nt | aa | nt | Order of Isolates: JS1, SB1, Gr10, CAN01 | |
| RNA-1 | 84–100 | 88–97 | 93–96 | 87–99 | (AB697826, AB697827, AM941711, HQ703343) |
| RNA-2 | 98–100 | 91–97 | 95–98 | 95–99 | (AB697828, AB697829, FM864225, HQ703344) |
| RNA-3 | 93–99 | 96–98 | 98–99 | 90–96 | (AB697843, AB697785, FM991954, HQ703345) |
| RNA-4 | 91–100 | 95–98 | 98–99 | 89–98 | (AB697857, AB697865, FM992851, HQ703346) |
| RNA-5 | 84–100 | 84–97 | 82–96 | 84–95 | (AB697871, AB697879, HE803826, n.a) |
| RNA-6 | 91–100 | 92–98 | 88–99 | 87–95 | (AB697885, AB697893, HE803827, n.a) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbeaino, T.; Marais, A.; Faure, C.; Trioano, E.; Candresse, T.; Parrella, G. High-Throughput Sequencing Reveals Cyclamen persicum Mill. as a Natural Host for Fig Mosaic Virus. Viruses 2018, 10, 684. https://doi.org/10.3390/v10120684
Elbeaino T, Marais A, Faure C, Trioano E, Candresse T, Parrella G. High-Throughput Sequencing Reveals Cyclamen persicum Mill. as a Natural Host for Fig Mosaic Virus. Viruses. 2018; 10(12):684. https://doi.org/10.3390/v10120684
Chicago/Turabian StyleElbeaino, Toufic, Armelle Marais, Chantal Faure, Elisa Trioano, Thierry Candresse, and Giuseppe Parrella. 2018. "High-Throughput Sequencing Reveals Cyclamen persicum Mill. as a Natural Host for Fig Mosaic Virus" Viruses 10, no. 12: 684. https://doi.org/10.3390/v10120684
APA StyleElbeaino, T., Marais, A., Faure, C., Trioano, E., Candresse, T., & Parrella, G. (2018). High-Throughput Sequencing Reveals Cyclamen persicum Mill. as a Natural Host for Fig Mosaic Virus. Viruses, 10(12), 684. https://doi.org/10.3390/v10120684