Subclinical Infection of Macaques and Baboons with A Baboon Simarterivirus


 ,
,  and
and 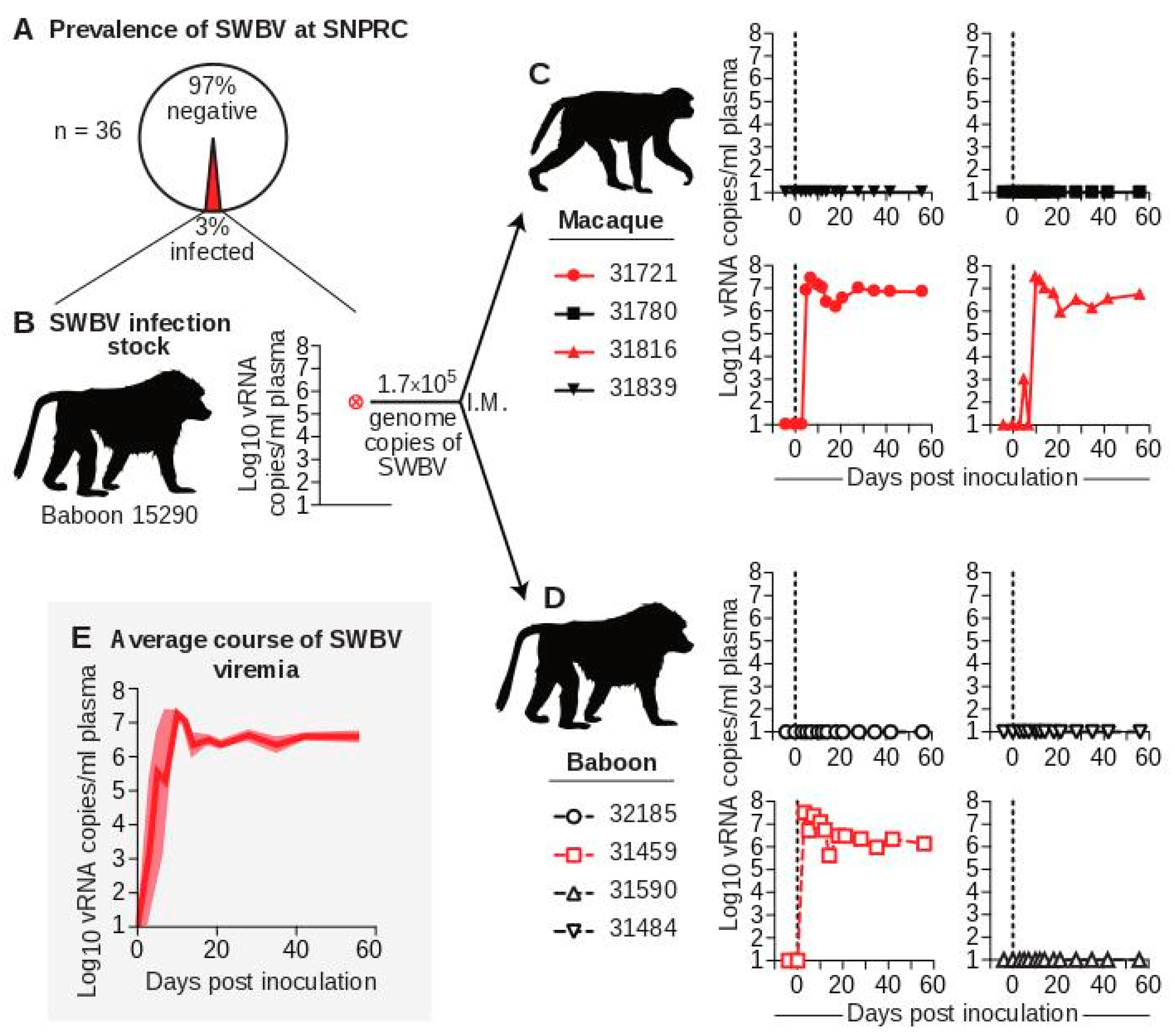{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Care and Ethical Use of Animals
2.3. Clinical Scoring
2.4. Virus Inoculations
2.5. Deep Sequencing
2.6. Sequence Read Mapping and Variant Calling
2.7. Viral Loads
2.8. Peptide Array
2.9. Flow Cytometry
2.10. Statistical Analysis
2.11. Data Accessibility
3. Results
3.1. Prevalence of SWBV Infection at SNPRC and Selection of SWBV-Naïve Baboons
3.2. The Course of SWBV Infection in Baboons and Macaques
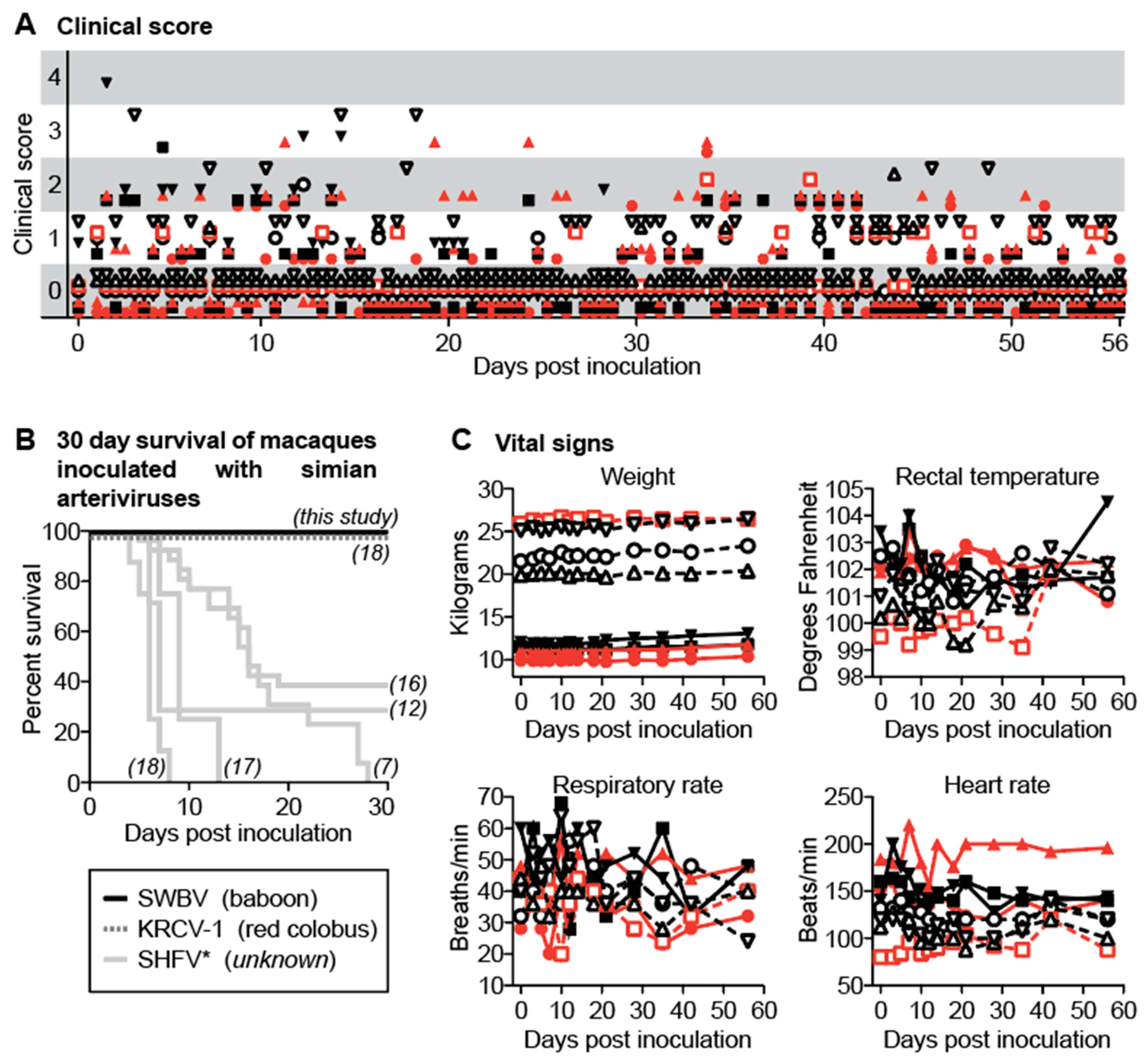
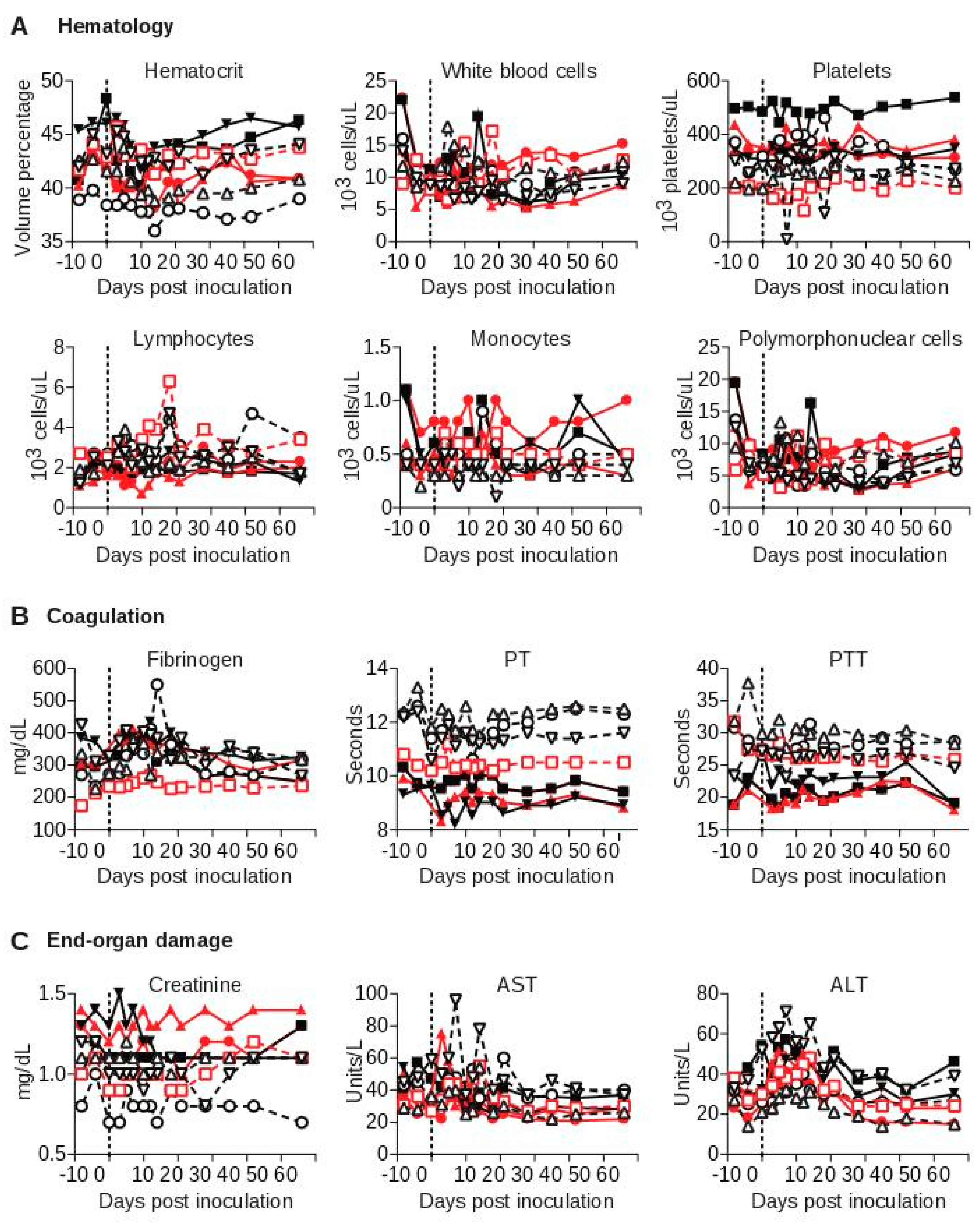
3.3. SWBV Does not Cause Overt Disease in Baboons or Macaques
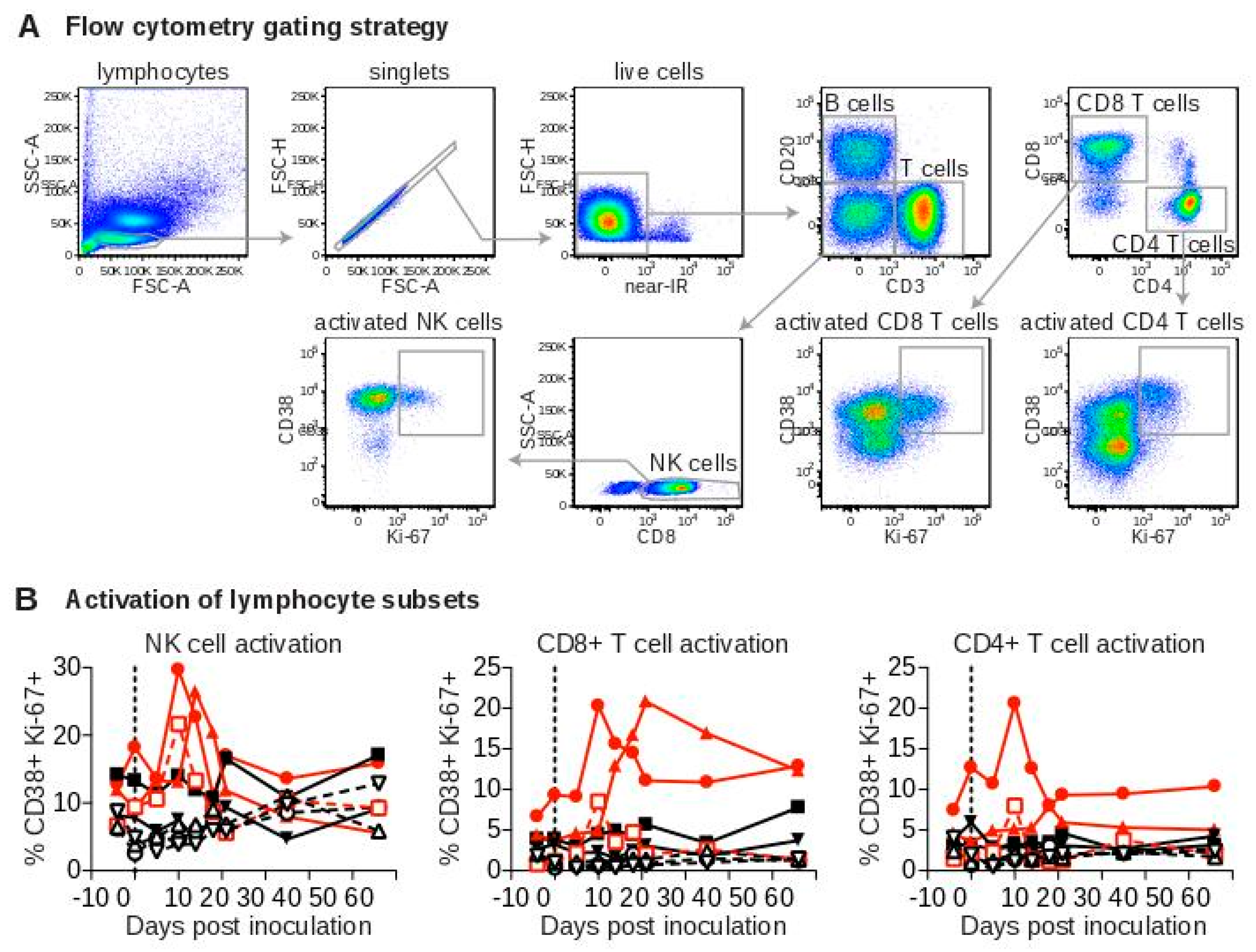
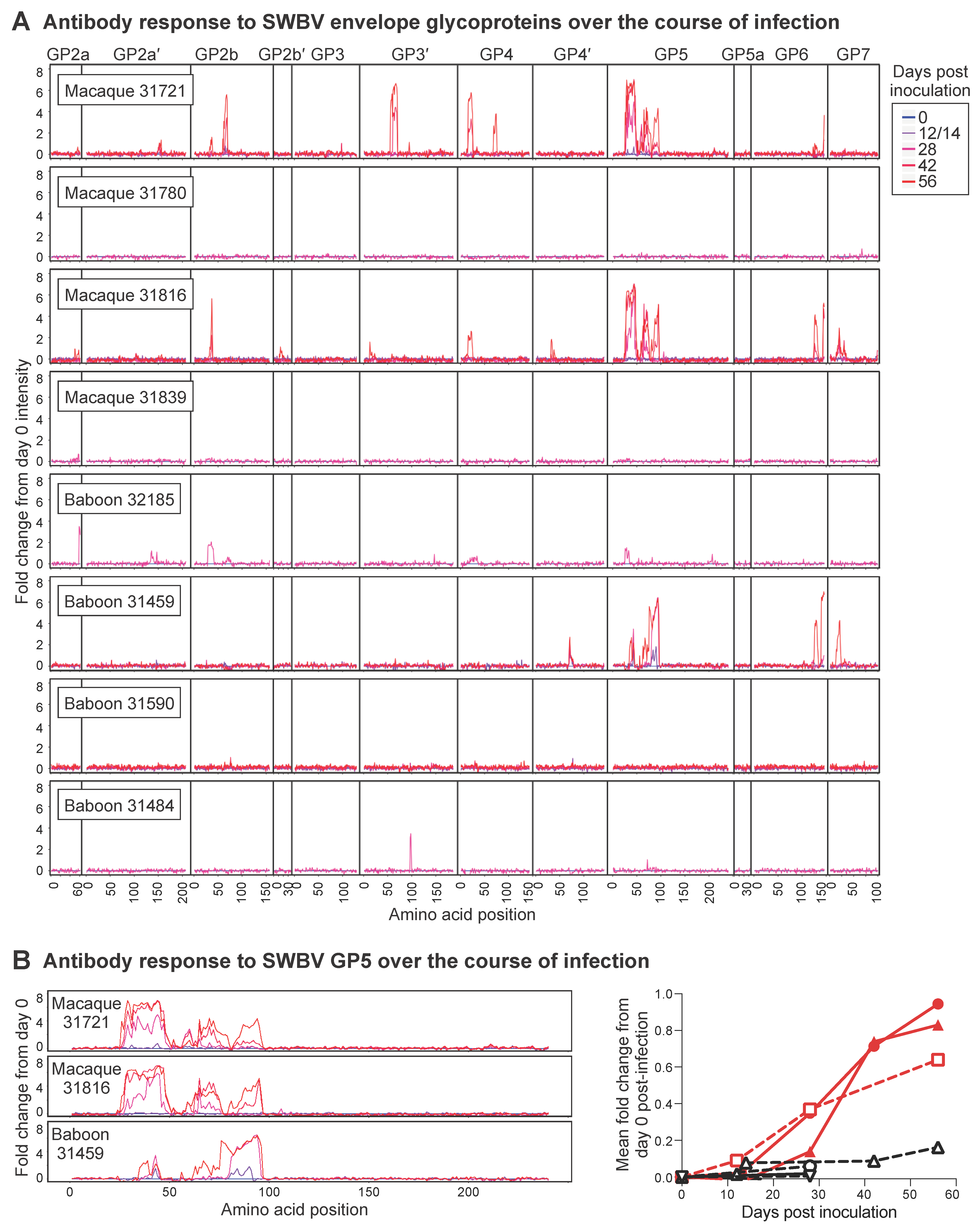
3.4. Arterivirus-Specific Adaptive Immune Responses in SWBV-Infected Macaques and Baboons
3.5. Intra-Host Evolution of SWBV over the Course of Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Davies, T.J.; Pedersen, A.B. Phylogeny and geography predict pathogen community similarity in wild primates and humans. Proc. Biol. Sci. R. Soc. 2008, 275, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, N.; Nunn, C.L. Identifying future zoonotic disease threats: Where are the gaps in our understanding of primate infectious diseases? Evol. Med. Public Health 2013, 2013, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Calvignac Spencer, S.; Leendertz, S.A.J.; Gillespie, T.R.; Leendertz, F.H. Wild great apes as sentinels and sources of infectious disease. Clin. Microbiol. Infect. 2012, 18, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, E.; Pauly, M.; Gillespie, T.R.; Akoua-Koffi, C.; Hohmann, G.; Fruth, B.; Karhemere, S.; Madinda, N.F.; Mugisha, L.; Muyembe, J.-J. Multiple cross-species transmission events of human adenoviruses (HAdV) during hominine evolution. Mol. Biol. Evol. 2015, 8, 2072–2084. [Google Scholar] [CrossRef] [PubMed]
- Bailey, A.L.; Lauck, M.; Sibley, S.D.; Friedrich, T.C.; Kuhn, J.H.; Freimer, N.B.; Jasinska, A.J.; Phillips-Conroy, J.E.; Jolly, C.J.; Marx, P.A.; et al. Zoonotic Potential of Simian Arteriviruses. J. Virol. 2015, 90, 630–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, A.L.; Lauck, M.; Ghai, R.R.; Nelson, C.W.; Heimbruch, K.; Hughes, A.L.; Goldberg, T.L.; Kuhn, J.H.; Jasinska, A.J.; Freimer, N.B. Arteriviruses, pegiviruses, and lentiviruses are common among wild African monkeys. J. Virol. 2016, 15, 6724–6737. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.E.; Allen, A.M.; Tauraso, N.M.; Shelokov, A. Simian hemorrhagic fever. I. Clinical and epizootiologic aspects of an outbreak among quarantined monkeys. Am. J. Trop. Med. Hyg. 1968, 17, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.M.; Palmer, A.E.; Tauraso, N.M.; Shelokov, A. Simian hemorrhagic fever. II. Studies in pathology. Am. J. Trop. Med. Hyg. 1968, 17, 413–421. [Google Scholar] [CrossRef]
- Tauraso, N.M.; Shelokov, A.; Palmer, A.E.; Allen, A.M. Simian hemorrhagic fever. III. Isolation and characterization of a viral agent. Am. J. Trop. Med. Hyg. 1968, 17, 422–431. [Google Scholar] [CrossRef]
- Lapin, B.A.; Pekerman, S.M.; Iakovleva, L.A.; Dzhikidze, E.K.; Shevtsova, Z.V.; Kuksova, M.I.; Dan’ko, L.V.; Krylova, R.I.; Akbroit, E.I.; Agrba, V.Z. Hemorrhagic fever in monkeys. Vopr. Virusol. 1967, 12, 168–173. [Google Scholar]
- Espana, C. Review of some outbreaks of viral disease in captive nonhuman primates. Lab. Anim. Sci. 1971, 21, 1023–1031. [Google Scholar] [PubMed]
- Abildgaard, C.; Harrison, J.; Espana, C.; Spangler, W.; Gribble, D. Simian hemorrhagic fever: Studies of coagulation and pathology. Am. J. Trop. Med. Hyg. 1975, 24, 537–544. [Google Scholar] [CrossRef] [PubMed]
- London, W.T. Epizootiology, transmission and approach to prevention of fatal simian haemorrhagic fever in rhesus monkeys. Nature 1977, 268, 344–345. [Google Scholar] [CrossRef] [PubMed]
- Renquist, D. Outbreak of simian hemorrhagic fever. J. Med. Primatol. 1990, 19, 77–79. [Google Scholar] [PubMed]
- Dalgard, D.W.; Hardy, R.J.; Pearson, S.L.; Pucak, G.J.; Quander, R.V.; Zack, P.M.; Peters, C.J.; Jahrling, P.B. Combined simian hemorrhagic fever and Ebola virus infection in cynomolgus monkeys. Lab. Anim. Sci. 1992, 42, 152–157. [Google Scholar] [PubMed]
- Johnson, R.F.; Dodd, L.E.; Yellayi, S.; Gu, W.; Cann, J.A.; Jett, C.; Bernbaum, J.G.; Ragland, D.R.; St Claire, M.; Byrum, R.; et al. Simian hemorrhagic fever virus infection of rhesus macaques as a model of viral hemorrhagic fever: Clinical characterization and risk factors for severe disease. Virology 2011, 421, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Vatter, H.A.; Donaldson, E.F.; Huynh, J.; Rawlings, S.; Manoharan, M.; Legasse, A.; Planer, S.; Dickerson, M.F.; Lewis, A.D.; Colgin, L.M.; et al. A simian hemorrhagic fever virus isolate from persistently infected baboons efficiently induces hemorrhagic fever disease in Japanese macaques. Virology 2015, 474, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Wahl-Jensen, V.; Johnson, J.C.; Lauck, M.; Weinfurter, J.T.; Moncla, L.H.; Weiler, A.M.; Charlier, O.; Rojas, O.; Byrum, R.; Ragland, D.R.; et al. Divergent Simian Arteriviruses Cause Simian Hemorrhagic Fever of Differing Severities in Macaques. MBio 2016, 7, e02009-15. [Google Scholar] [CrossRef]
- Lauck, M.; Alkhovsky, S.V.; Bào, Y.M.; Bailey, A.L.; Shevtsova, Z.V.; Shchetinin, A.M.; Vishnevskaya, T.V.; Lackemeyer, M.G.; Postnikova, E.; Mazur, S.; et al. Historical Outbreaks of Simian Hemorrhagic Fever in Captive Macaques Were Caused by Distinct Arteriviruses. J. Virol. 2015, 15, 8082–8087. [Google Scholar] [CrossRef]
- Bailey, A.L.; Lauck, M.; Sibley, S.D.; Pecotte, J.; Rice, K.; Weny, G.; Tumukunde, A.; Hyeroba, D.; Greene, J.; Correll, M.; et al. Two novel simian arteriviruses in captive and wild baboons (Papio spp.). J. Virol. 2014, 88, 13231–13239. [Google Scholar] [CrossRef]
- BioProject. SWBV Sequencing from Macaques and Baboons at SNPRC. Accession number: PRJNA483240. Available online: https://www.ncbi.nlm.nih.gov/bioproject/483240 (accessed on 7 December 2018).
- Joint Genome Institute. BBMap Guide. Available online: https://jgi.doe.gov/data-and-tools/bbtools/bb-tools-user-guide/bbmap-guide/ (accessed on 7 December 2018).
- SnpEff Genomic Variant Annotations and Functional Effect Prediction Toolbox. Available online: http://snpeff.sourceforge.net/ (accessed on 7 December 2018).
- University of Wisconsin-Madison AIDS Vaccine Research Laboratory. Non-pathogenic Infection of Macaques and Baboons with a Baboon Arterivirus. Available online: https://dholk.primate.wisc.edu/project/dho/public/manuscripts/201808-SNPRC-SWBV/begin.view? (accessed on 7 December 2018).
- Dokland, T. The structural biology of PRRSV. Virus Res. 2010, 154, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Gravell, M.; Palmer, A.E.; Rodriguez, M.; London, W.T.; Hamilton, R.S. Method to detect asymptomatic carriers of simian hemorrhagic fever virus. Lab. Anim. Sci. 1980, 30, 988–991. [Google Scholar] [PubMed]
- Bailey, A.L.; Lauck, M.; Weiler, A.; Sibley, S.D.; Dinis, J.M.; Bergman, Z.; Nelson, C.W.; Correll, M.; Gleicher, M.; Hyeroba, D.; et al. High genetic diversity and adaptive potential of two simian hemorrhagic fever viruses in a wild primate population. PLoS ONE 2014, 9, e90714. [Google Scholar] [CrossRef] [PubMed]
- Lauck, M.; Switzer, W.M.; Sibley, S.D.; Hyeroba, D.; Tumukunde, A.; Weny, G.; Taylor, B.; Shankar, A.; Ting, N.; Chapman, C.A.; et al. Discovery and full genome characterization of two highly divergent simian immunodeficiency viruses infecting black-and-white colobus monkeys (Colobus guereza) in Kibale National Park, Uganda. Retrovirology 2013, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Lauck, M.; Sibley, S.D.; Hyeroba, D.; Tumukunde, A.; Weny, G.; Chapman, C.A.; Ting, N.; Switzer, W.M.; Kuhn, J.H.; Friedrich, T.C.; et al. Exceptional simian hemorrhagic Fever virus diversity in a wild african primate community. J. Virol. 2013, 87, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Gravell, M.; London, W.T.; Rodriguez, M.; Palmer, A.E.; Hamilton, R.S.; Curfman, B.L. Studies on simian hemorrhagic fever virus infection of patas monkeys - I. Serology. In The Comparative Pathology of Zoo Animals (the Symposia of the National Zoological Park); Montali, R.J., Migaki, G., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1980; pp. 167–170. [Google Scholar]
- Gravell, M.; London, W.T.; Leon, M.E.; Palmer, A.E.; Hamilton, R.S. Differences among isolates of simian hemorrhagic fever (SHF) virus. Proc. Soc. Exp. Biol. Med. (N. Y.) 1986, 181, 112–119. [Google Scholar] [CrossRef]
- Gravell, M.; London, W.T.; Leon, M.; Palmer, A.E.; Hamilton, R.S. Elimination of persistent simian hemorrhagic fever (SHF) virus infection in patas monkeys. Proc. Soc. Exp. Biol. Med. (N. Y.) 1986, 181, 219–225. [Google Scholar] [CrossRef]
- Balasuriya, U.B.; Patton, J.F.; Rossitto, P.V.; Timoney, P.J.; McCollum, W.H.; MacLachlan, N.J. Neutralization determinants of laboratory strains and field isolates of equine arteritis virus: Identification of four neutralization sites in the amino-terminal ectodomain of the G(L) envelope glycoprotein. Virology 1997, 232, 114–128. [Google Scholar] [CrossRef]
- Plagemann, P.G. Complexity of the single linear neutralization epitope of the mouse arterivirus lactate dehydrogenase-elevating virus. Virology 2001, 290, 11–20. [Google Scholar] [CrossRef]
- Plagemann, P.G.W.; Rowland, R.R.R.; Faaberg, K.S. The primary neutralization epitope of porcine respiratory and reproductive syndrome virus strain VR-2332 is located in the middle of the GP5 ectodomain. Arch. Virol. 2002, 147, 2327–2347. [Google Scholar] [CrossRef]
- Plagemann, P.G.W. The primary GP5 neutralization epitope of North American isolates of porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2004, 102, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain. J. Virol. 2002, 76, 4241–4250. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buechler, C.R.; Semler, M.; Baker, D.A.; Newman, C.; Cornish, J.P.; Chavez, D.; Guerra, B.; Lanford, R.; Brasky, K.; Kuhn, J.H.; et al. Subclinical Infection of Macaques and Baboons with A Baboon Simarterivirus. Viruses 2018, 10, 701. https://doi.org/10.3390/v10120701
Buechler CR, Semler M, Baker DA, Newman C, Cornish JP, Chavez D, Guerra B, Lanford R, Brasky K, Kuhn JH, et al. Subclinical Infection of Macaques and Baboons with A Baboon Simarterivirus. Viruses. 2018; 10(12):701. https://doi.org/10.3390/v10120701
Chicago/Turabian StyleBuechler, Connor R., Matthew Semler, David A. Baker, Christina Newman, Joseph P. Cornish, Deborah Chavez, Bernadette Guerra, Robert Lanford, Kathy Brasky, Jens H. Kuhn, and et al. 2018. "Subclinical Infection of Macaques and Baboons with A Baboon Simarterivirus" Viruses 10, no. 12: 701. https://doi.org/10.3390/v10120701
APA StyleBuechler, C. R., Semler, M., Baker, D. A., Newman, C., Cornish, J. P., Chavez, D., Guerra, B., Lanford, R., Brasky, K., Kuhn, J. H., Johnson, R. F., O’Connor, D. H., & Bailey, A. L. (2018). Subclinical Infection of Macaques and Baboons with A Baboon Simarterivirus. Viruses, 10(12), 701. https://doi.org/10.3390/v10120701