Fowl Adenovirus 4 (FAdV-4)-Based Infectious Clone for Vaccine Vector Development and Viral Gene Function Studies

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Polymerase Chain Reaction Amplification
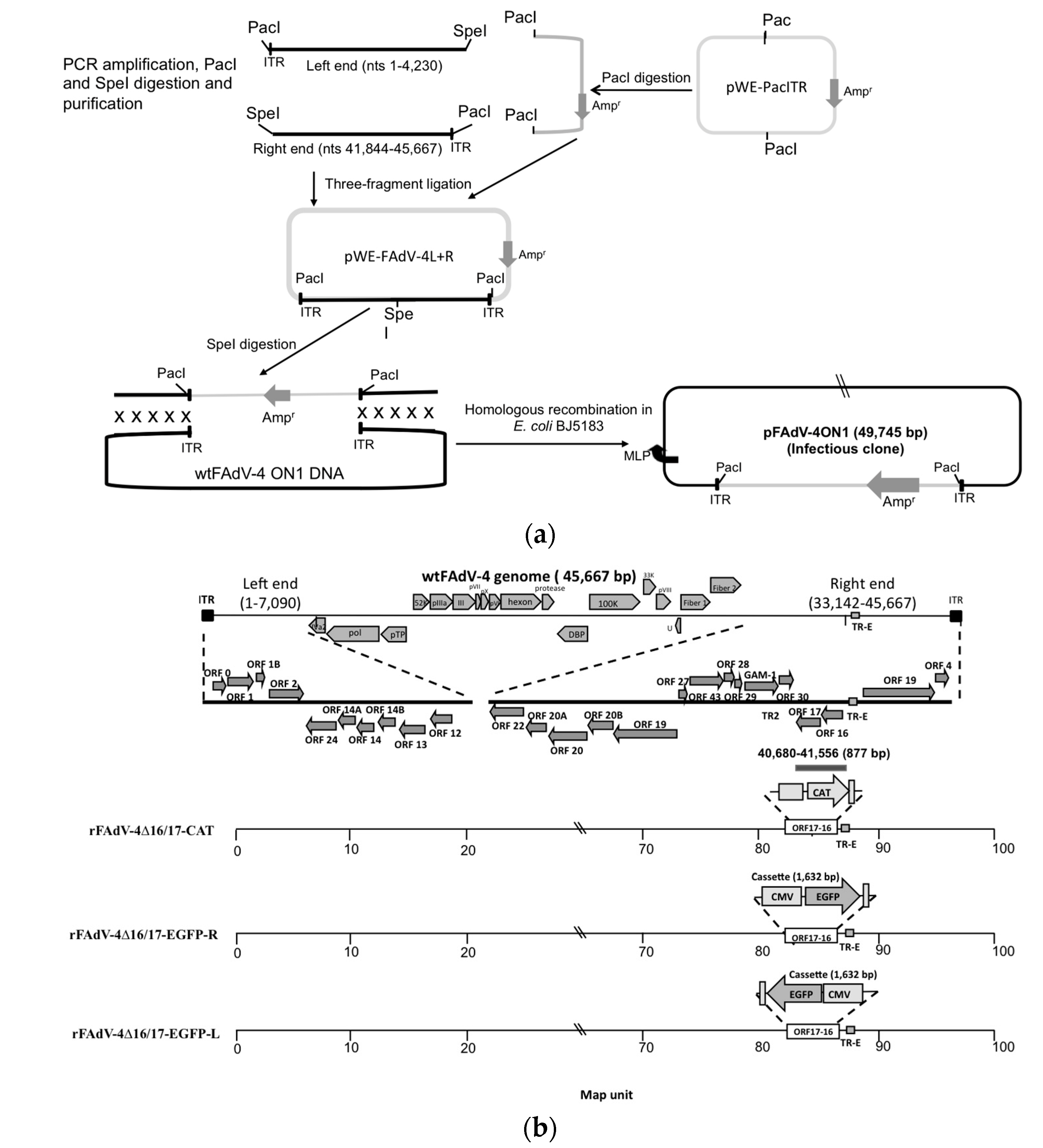
2.3. Construction of FAdV-4 ON1 Infectious Clone
2.4. Construction of ORFs 16- and 17-Deleted Mutant/Recombinant Viruses
2.5. One-Step Growth Curves
3. Results
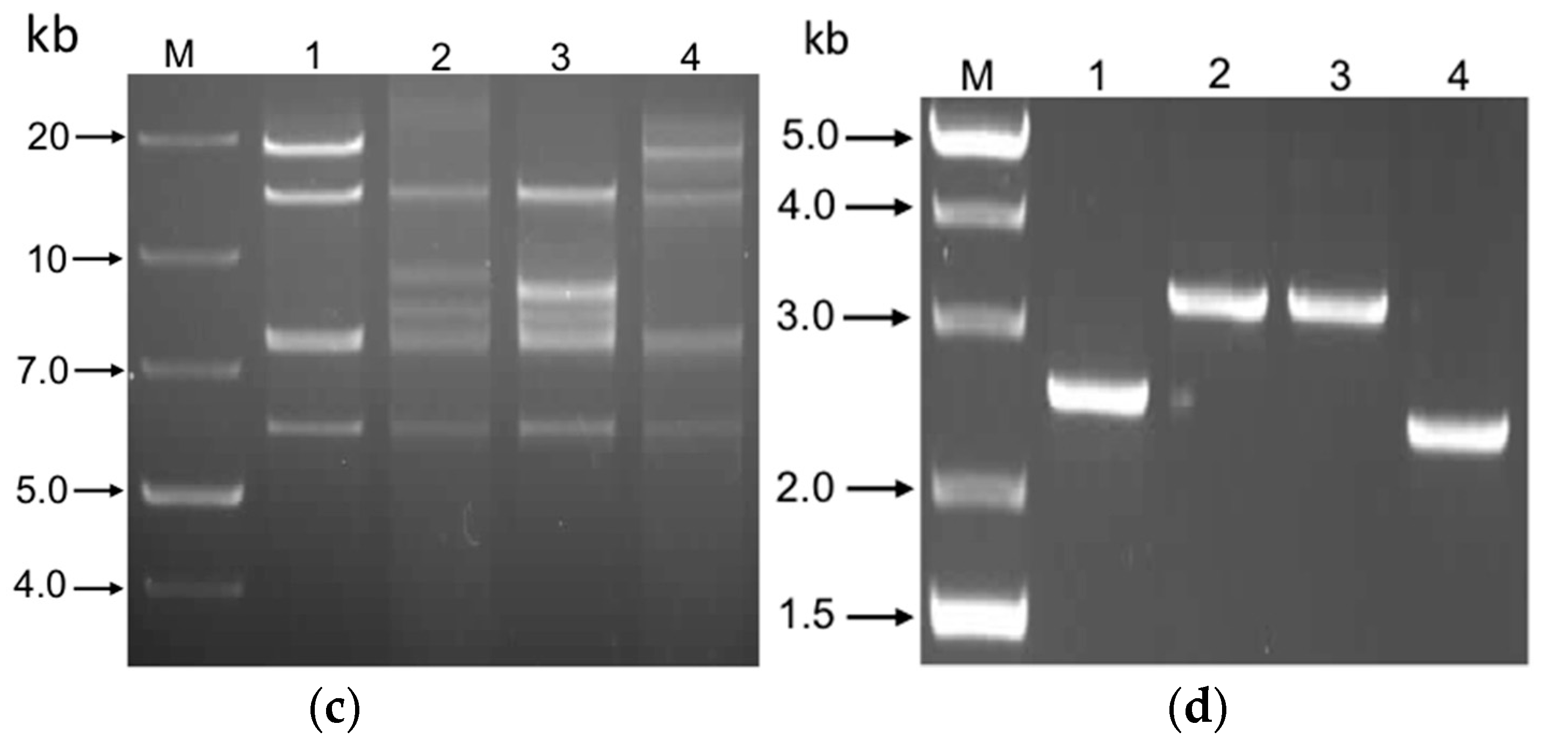
3.1. Generation of FAdV-4 Infectious Clone (pFAdV-4 ON1) and ORF16–17-Deleted Mutant/Recombinant Viruses
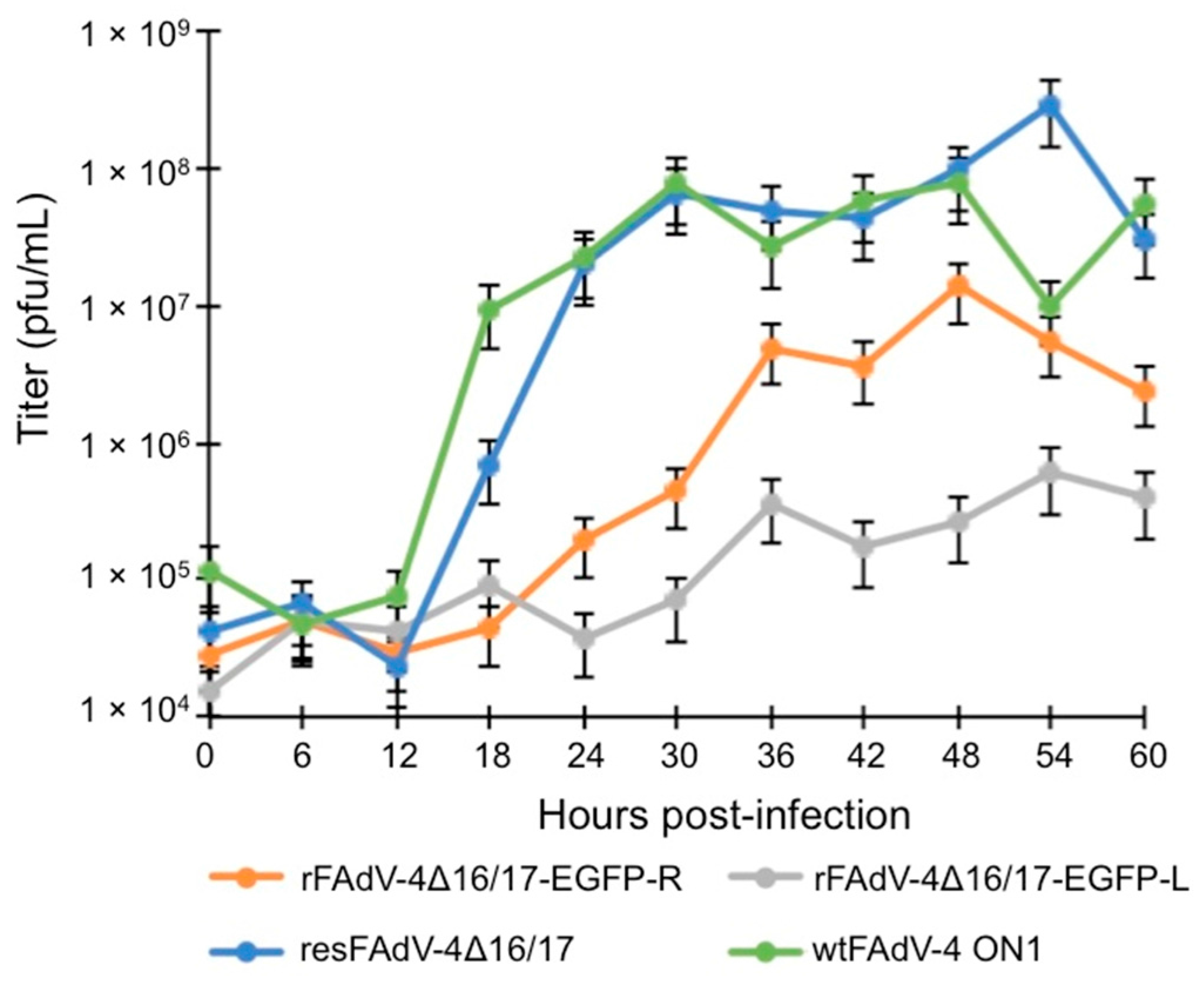
3.2. Viral Growth Kinetics
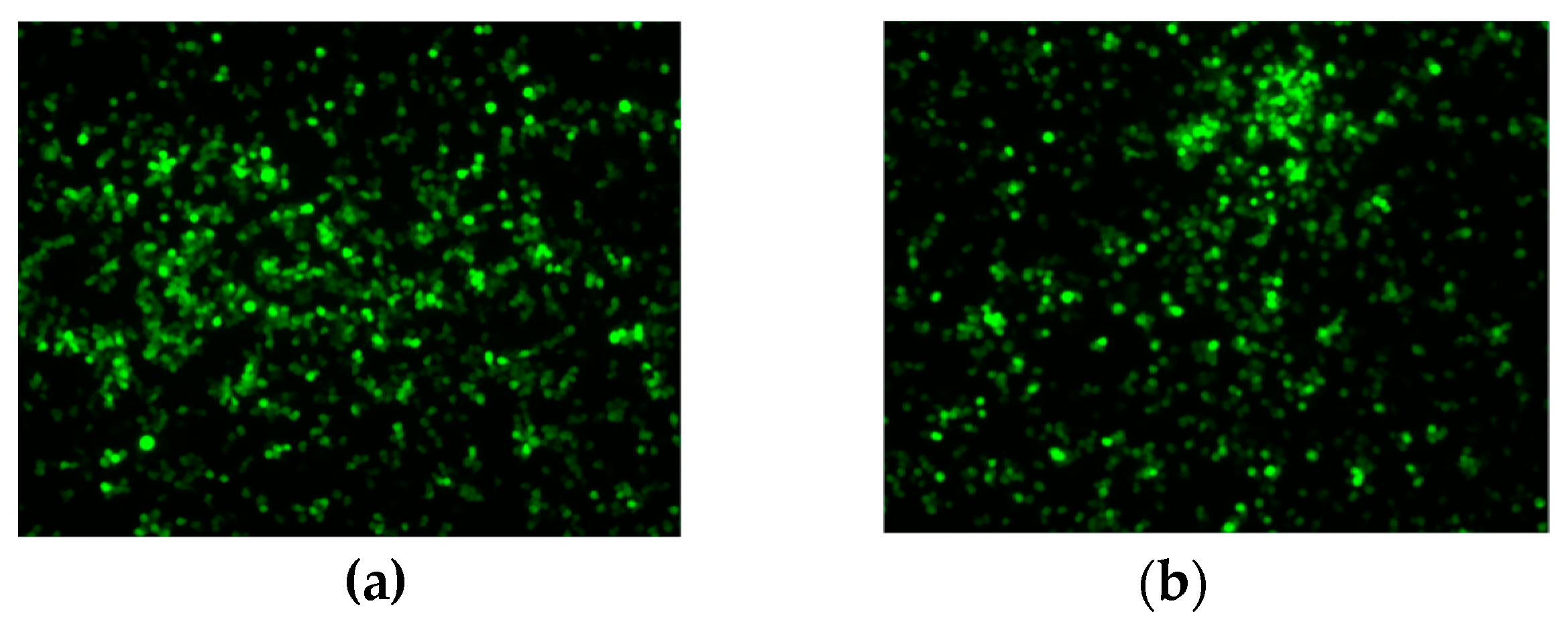
3.3. Transgene Expression Upon Infection with Recombinant Viruses
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rahul, S.; Kataria, J.M.; Senthilkumar, N.; Dhama, K.; Sylvester, S.A.; Uma, R. Association of fowl adenovirus serotype 12 with hydropericardium syndrome of poultry in India. Acta Virol. 2005, 49, 139–143. [Google Scholar] [PubMed]
- Zhang, T.; Jin, Q.; Ding, P.; Wang, Y.; Chai, Y.; Li, Y.; Liu, X.; Luo, J.; Zhang, G. Molecular epidemiology of hydropericardium syndrome outbreak-associated serotype 4 fowl adenovirus isolates in central China. Virol. J. 2016, 13, 14–16. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhong, Q.; Zhao, Y.; Hu, Y.-X.; Zhang, G.-Z. Pathogenicity and Complete Genome Characterization of Fowl Adenoviruses Isolated from Chickens Associated with Inclusion Body Hepatitis and Hydropericardium Syndrome in China. PLoS ONE 2015, 10, e0133073. [Google Scholar] [CrossRef] [PubMed]
- Cheema, A.H.; Ahmad, J.; Afzal, M. An adenovirus infection of poultry in Pakistan. Rev. Sci. Tech. Off. Int. Epiz 1989, 8, 789–795. [Google Scholar] [CrossRef]
- Asthana, M.; Chandra, R.; Kumar, R. Hydropericardium syndrome: Current state and future developments. Arch. Virol. 2013, 158, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Pallister, J.; Wright, P.J.; Sheppard, M. A single gene encoding the fiber is responsible for variations in virulence in the fowl adenoviruses. J. Virol. 1996, 70, 5115–5122. [Google Scholar] [PubMed]
- Pan, Q.; Yang, Y.; Shi, Z.; Liu, L.; Gao, Y.; Qi, X.; Liu, C.; Zhang, Y.; Cui, H.; Wang, X. Different dynamic distribution in chickens and ducks of the hypervirulent, novel genotype fowl adenovirus serotype 4 recently emerged in China. Front. Microbiol. 2017, 8, 1005. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.D.; Nagy, É. Coding potential and transcript analysis of fowl adenovirus 4: Insight into upstream ORFs as common sequence features in adenoviral transcripts. J. Gen. Virol. 2011, 92, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Grgić, H.; Poljak, Z.; Sharif, S.; Nagy, É. Pathogenicity and cytokine gene expression pattern of a serotype 4 fowl adenovirus isolate. PLoS ONE 2013, 8, e77601. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wan, W.; Gao, D.; Li, Y.; Yang, X.; Liu, H.; Yao, H.; Chen, L.; Wang, C.; Zhao, J. Genetic characterization of novel fowl aviadenovirus 4 isolates from outbreaks of hepatitis-hydropericardium syndrome in broiler chickens in China. Emerg. Microbes Infect. 2016, 5, e117. [Google Scholar] [CrossRef] [PubMed]
- Vera-Hernández, P.F.; Morales-Garzón, A.; Cortés-Espinosa, D.V.; Galiote-Flores, A.; García-Barrera, L.J.; Rodríguez-Galindo, E.T.; Toscano-Contreras, A.; Lucio-Decanini, E.; Absalón, A.E. Clinicopathological characterization and genomic sequence differences observed in a highly virulent fowl Aviadenovirus serotype 4. Avian Pathol. 2016, 45, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Liu, L.; Wang, Y.; Zhang, Y.; Qi, X.; Liu, C.; Gao, Y.; Wang, X.; Cui, H. The first whole genome sequence and pathogenicity characterization of a fowl adenovirus 4 isolated from ducks associated with inclusion body hepatitis and hydropericardium syndrome. Avian Pathol. 2017, 46, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Liang, G.; Zhang, J.; Wang, W.; Song, N.; Wang, P.; Zheng, W.; Xie, Q.; Shao, H.; Wan, Z.; et al. Outbreaks of serotype 4 fowl adenovirus with novel genotype, China. Emerg. Microbes Infect. 2016, 5, e50. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, M.; Werner, W.; Tsatas, E.; McCoy, R.; Prowse, S.; Johnson, M. Fowl adenovirus recombinant expressing VP2 of infectious bursal disease virus induces protective immunity against bursal disease. Arch. Virol. 1998, 143, 915–930. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; Pooley, C.; Ignjatovic, J.; Tyack, S.G. A recombinant fowl adenovirus expressing the S1 gene of infectious bronchitis virus protects against challenge with infectious bronchitis virus. Vaccine 2003, 21, 2730–2736. [Google Scholar] [CrossRef]
- Michou, A.I.; Lehrmann, H.; Saltik, M.; Cotten, M. Mutational analysis of the avian adenovirus CELO, which provides a basis for gene delivery vectors. J. Virol. 1999, 73, 1399–1410. [Google Scholar] [PubMed]
- Ojkic, D.; Nagy, É. The long repeat region is dispensable for fowl adenovirus replication in vitro. Virology 2001, 283, 197–206. [Google Scholar] [CrossRef] [PubMed]
- François, A.; Eterradossi, N.; Delmas, B.; Payet, V.; Langlois, P. Construction of avian adenovirus CELO recombinants in cosmids. J. Virol. 2001, 75, 5288–5301. [Google Scholar] [CrossRef] [PubMed]
- Corredor, J.C.; Nagy, É. The non-essential left end region of the fowl adenovirus 9 genome is suitable for foreign gene insertion/replacement. Virus Res. 2010, 149, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Qin, X.; Krell, P.; Lu, R.; Sharif, S.; Nagy, É. Characterization and functional studies of fowl adenovirus 9 dUTPase. Virology 2016, 497, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Griffin, B.; de Jong, J.; Krell, P.J.; Nagy, É. Rapid generation of fowl adenovirus 9 vectors. J. Virol. Methods 2015, 223, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Corredor, J.C.; Garceac, A.; Krell, P.J.; Nagy, É. Sequence comparison of the right end of fowl adenovirus genomes. Virus Genes 2008, 36, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Ballmann, M.Z.; Kosiol, C.; Harrach, B.; Schlotterer, C.; Hess, M. Whole-genome sequences of two turkey adenovirus types reveal the existence of two unknown lineages that merit the establishment of novel species within the genus Aviadenovirus. J. Gen. Virol. 2014, 95, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Kaján, G.L.; Davison, A.J.; Palya, V.; Harrach, B.; Benko, M. Genome sequence of a waterfowl aviadenovirus, goose adenovirus 4. J. Gen. Virol. 2012, 93, 2457–2465. [Google Scholar] [CrossRef] [PubMed]
- Washietl, S.; Eisenhaber, F. Reannotation of the CELO genome characterizes a set of previously unassigned open reading frames and points to novel modes of host interaction in avian adenoviruses. BMC Bioinformatics 2003, 4, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, Y.; Krell, P.J.; Nagy, É. Generation and characterization of a fowl adenovirus 9 dual-site expression vector. J. Biotechnol. 2018, 266, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.S.; Huber, P.; Cao, J.; Krell, P.J.; Nagy, É. Growth characteristics of fowl adenovirus type 8 in a chicken hepatoma cell line. J. Virol. Methods 1998, 74, 9–14. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Cloning and Transformation with Plasmid Vectors. In Molecular Cloning: A Laboratory Manual; Inglis, J., Boyle, A., Gann, A., Eds.; Cold Spring Harbor laboratory Press: Cold Spring Harbor, NY, USA, 2012; Volume 1, pp. 157–258. [Google Scholar]
- Corredor, J.C.; Pei, Y.; Nagy, É. Fowl Adenovirus-Based Vaccine Platform. In Methods in Molecular Biology; Ferran, M.C., Skuse, G.R., Eds.; Springer: Rochester, NY, USA, 2017; Volume 1581, pp. 29–54. [Google Scholar]
- Quick, L.N.; Shah, A.; Wilson, J.W. A series of vectors with alternative antibiotic resistance markers for use in Lambda Red recombination. J. Microbiol. Biotechnol. 2010, 20, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Francois, A.; Chevalier, C.; Delmas, B.; Eterradossi, N.; Toquin, D.; Rivallan, G.; Langlois, P. Avian adenovirus CELO recombinants expressing VP2 of infectious bursal disease virus induce protection against bursal disease in chickens. Vaccine 2004, 22, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; Pooley, C.; Lowenthal, J.W. Delivery of avian cytokines by adenovirus vectors. Dev. Comp. Immunol. 2000, 24, 343–354. [Google Scholar] [CrossRef]
- Deng, L.; Sharif, S.; Nagy, É. Oral inoculation of chickens with a candidate fowl adenovirus 9 vector. Clin. Vaccine Immunol. 2013, 20, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Schonewille, E.; Jaspers, R.; Paul, G.; Hess, M. Specific-pathogen-free chickens vaccinated with a live FAdV-4 vaccine are fully protected against a severe challenge even in the absence of neutralizing antibodies. Avian Dis. 2010, 54, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Lim, T.H.; Lee, D.H.; Youn, H.N.; Yuk, S.S.; Kim, B.Y.; Choi, S.W.; Jung, C.H.; Han, J.H.; Song, C.S. An inactivated oil-emulsion fowl Adenovirus serotype 4 vaccine provides broad cross-protection against various serotypes of fowl Adenovirus. Vaccine 2014, 32, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Schachner, A.; Marek, A.; Jaskulska, B.; Bilic, I.; Hess, M. Recombinant FAdV-4 fiber-2 protein protects chickens against hepatitis-hydropericardium syndrome (HHS). Vaccine 2014, 32, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.S.; Ashraf, A.; Khan, M.I.; Rahman, M.; Habib, M.; Chughtai, M.I.; Qureshi, J.A. Fowl adenovirus: history, emergence, biology and development of a vaccine against hydropericardium syndrome. Arch. Virol. 2017, 162, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Han, H.; Li, J.; Wong, L. DNAFSMiner: A web-based software toolbox to recognize two types of functional sites in DNA sequences. Bioinformatics 2005, 21, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Glotzer, J.B.; Saltik, M.; Chiocca, S.; Michou, A.I.; Moseley, P.; Cotten, M. Activation of heat-shock response by an adenovirus is essential for virus replication. Nature 2000, 407, 207–211. [Google Scholar] [PubMed]
- Fraser, N.W.; Nevins, J.R.; Ziff, E.; Darnell, J.E. The major late adenovirus type-2 transcription unit: Termination is downstream from the last poly(A) site. J. Mol. Biol. 1979, 129, 643–656. [Google Scholar] [CrossRef]
- Ginn, S.L.; Fleming, J.; Rowe, P.B.; Alexander, I.E. Promoter interference mediated by the U3 region in early-generation HIV-1-derived lentivirus vectors can influence detection of transgene expression in a cell-type and species-specific manner. Hum. Gene Ther. 2003, 14, 1127–1137. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5’-3’) | Location | Purpose |
|---|---|---|---|
| FAdV4R-EndPacI-F | agtcTTAATTAAcatcatcttatataaccgcgtcttttgacac | 1–31 | Generate a 4.2-kb fragment (left end genomic region) with PacI and SpeI restriction sites |
| FAdV4R-EndSpeI–R | agtcACTAGTcttacctcggatgaactatgccactg | 4205–4230 | |
| FAdV4L-EndSpeI-F | cacaaggtacatgaatcACTAGTaatggtc | 41,844–41,873 | Generate a 3.8-kb fragment (right end genomic region) with PacI and SpeI restriction sites |
| FAdV4L-EndPacI-R | agtcTTAATTAAcatcatcttatataAccgcgtcttttgacacacttac | 45,631–45,667 | |
| FAdV-4ORF17CAT-F | catgacacagagggaggagactgcgagtaatcacctttaattattaacagctATTTAAATgtgtaggctggagctgcttc | 40,629–40,680 | Introduce CAT-gene expression cassette with upstream and downstream homologous arms to ORFs17 and 16, respectively |
| FAdV-4ORF16CAT-R | gagcaggaaaatctgcagagcactcttttggcggtcccgtgtgcggtgggtaATTTAAATcatatgaatatcctccttagttc | 41,557–41,608 | |
| FAdV-4Ver-F | cgactcctcctctttgtgggc | 40,085–40,105 | Verification |
| FAdV-4Ver-R | gcggcatctcctagaatgagg | 42,518–42,538 | Verification |
| EGFPcaSwaI-F | agctgcATTTAAATgtattaccgccatgcattag | 4717–3 | EGFP cassette amplification |
| EGFPcaSwaI-R | agctgcATTTAAATccacaactagaatgcagtg | 1597–1615 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pei, Y.; Corredor, J.C.; Griffin, B.D.; Krell, P.J.; Nagy, É. Fowl Adenovirus 4 (FAdV-4)-Based Infectious Clone for Vaccine Vector Development and Viral Gene Function Studies. Viruses 2018, 10, 97. https://doi.org/10.3390/v10020097
Pei Y, Corredor JC, Griffin BD, Krell PJ, Nagy É. Fowl Adenovirus 4 (FAdV-4)-Based Infectious Clone for Vaccine Vector Development and Viral Gene Function Studies. Viruses. 2018; 10(2):97. https://doi.org/10.3390/v10020097
Chicago/Turabian StylePei, Yanlong, Juan C. Corredor, Bryan D. Griffin, Peter J. Krell, and Éva Nagy. 2018. "Fowl Adenovirus 4 (FAdV-4)-Based Infectious Clone for Vaccine Vector Development and Viral Gene Function Studies" Viruses 10, no. 2: 97. https://doi.org/10.3390/v10020097
APA StylePei, Y., Corredor, J. C., Griffin, B. D., Krell, P. J., & Nagy, É. (2018). Fowl Adenovirus 4 (FAdV-4)-Based Infectious Clone for Vaccine Vector Development and Viral Gene Function Studies. Viruses, 10(2), 97. https://doi.org/10.3390/v10020097