Feline APOBEC3s, Barriers to Cross-Species Transmission of FIV?


 ,
, 
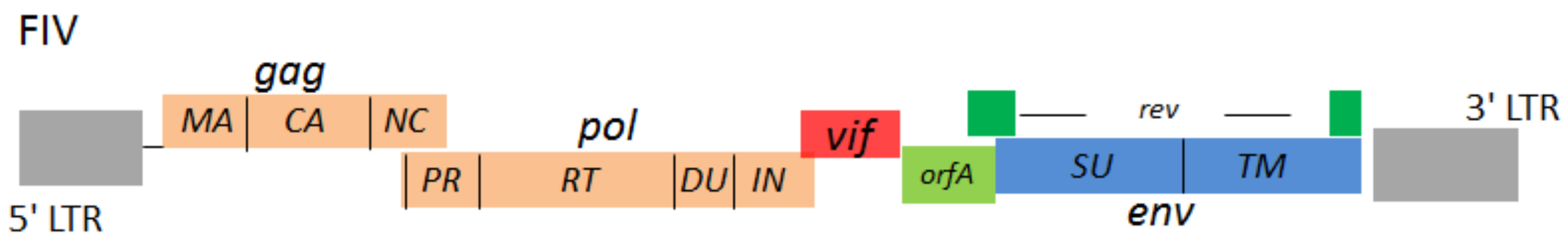{kind=link}
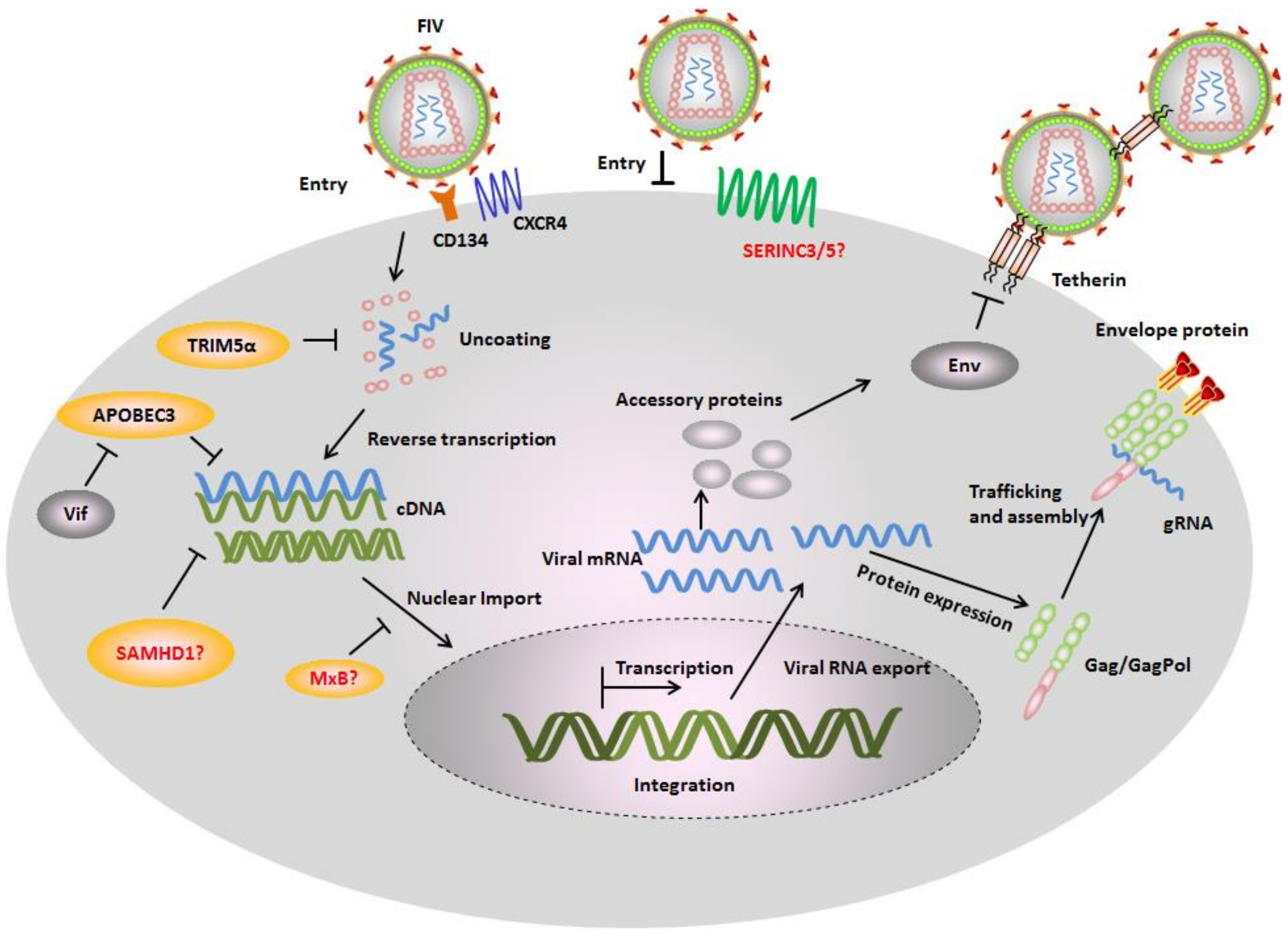{kind=link}
{kind=link}
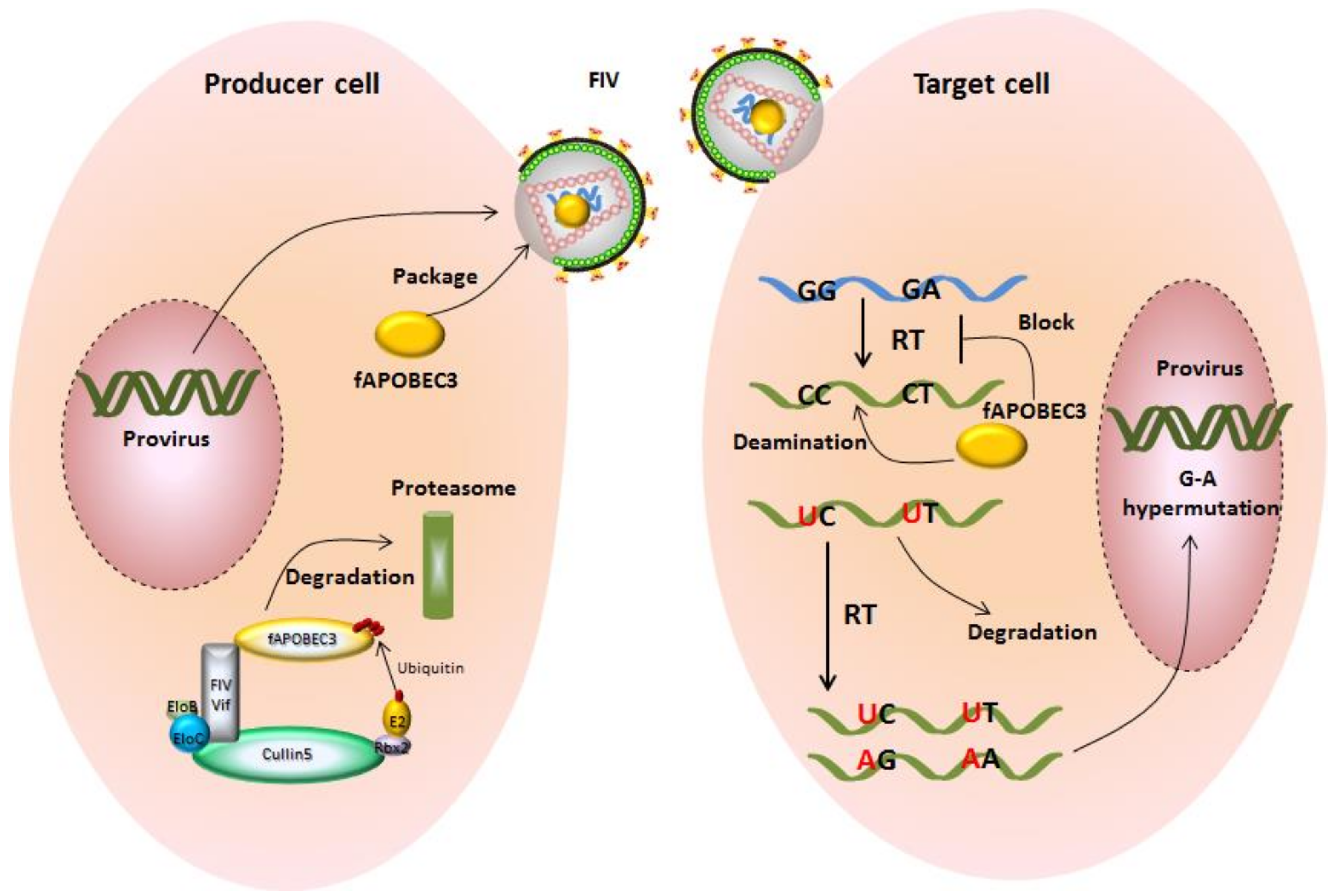{kind=link}
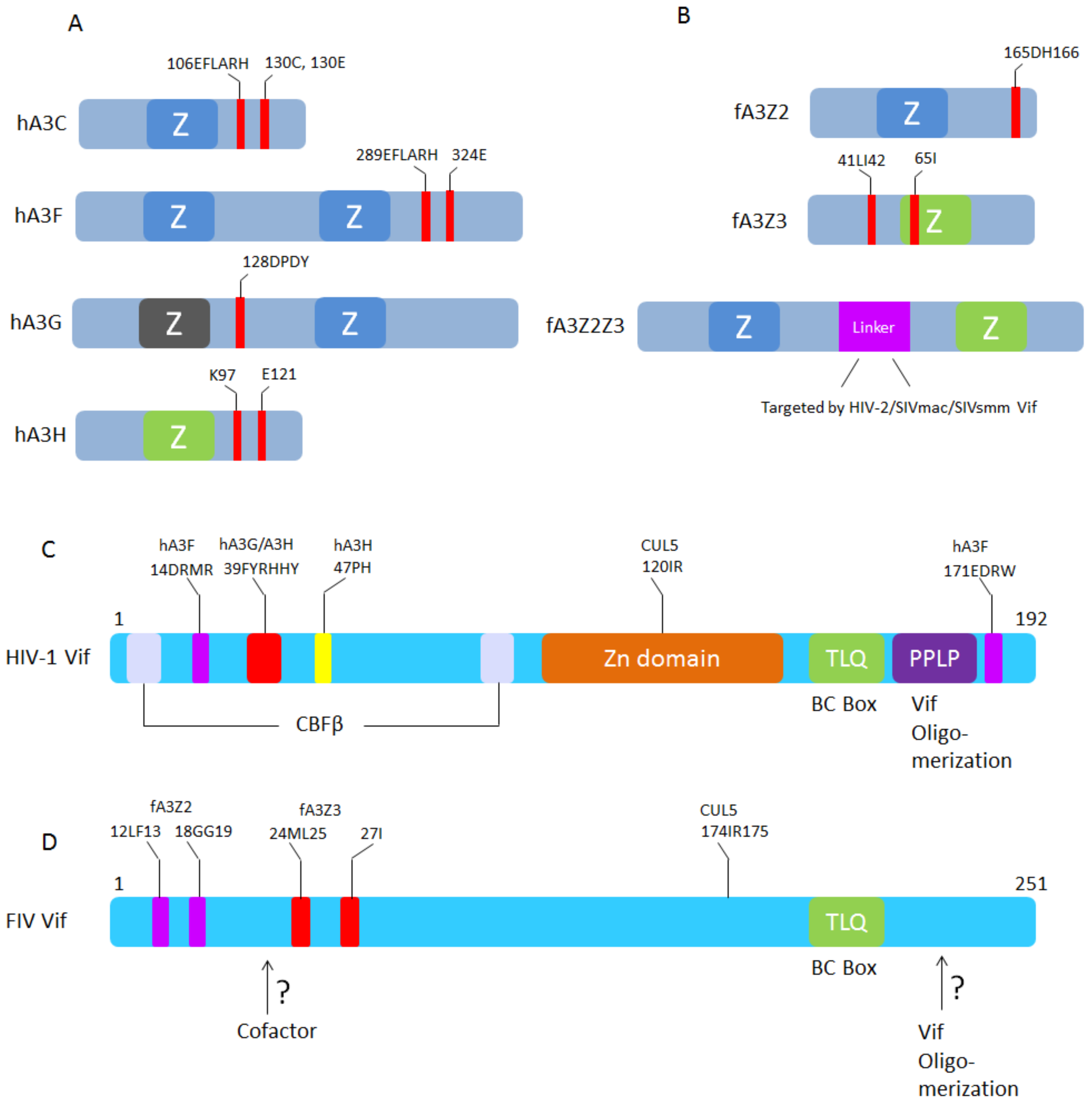{kind=link}
Abstract
:1. Introduction
2. FIV Vif and Feline APOBEC3
3. Feline Foamy Virus and Feline Leukemia Virus
4. FIV Cross-Species Transmission
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pedersen, N.C.; Ho, E.W.; Brown, M.L.; Yamamoto, J.K. Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science 1987, 235, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Yamamoto, J.K.; Ishida, T.; Hansen, H. Feline immunodeficiency virus infection. Vet. Immunol. Immunopathol. 1989, 21, 111–129. [Google Scholar] [CrossRef]
- Hayward, J.J.; Rodrigo, A.G. Molecular epidemiology of feline immunodeficiency virus in the domestic cat (Felis catus). Vet. Immunol. Immunopathol. 2010, 134, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Shimojima, M.; Miyazawa, T.; Ikeda, Y.; McMonagle, E.L.; Haining, H.; Akashi, H.; Takeuchi, Y.; Hosie, M.J.; Willett, B.J. Use of CD134 as a primary receptor by the feline immunodeficiency virus. Science 2004, 303, 1192–1195. [Google Scholar] [CrossRef] [PubMed]
- De Parseval, A.; Chatterji, U.; Sun, P.; Elder, J.H. Feline immunodeficiency virus targets activated CD4+ T cells by using CD134 as a binding receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 13044–13049. [Google Scholar] [CrossRef] [PubMed]
- Willett, B.J.; McMonagle, E.L.; Ridha, S.; Hosie, M.J. Differential utilization of CD134 as a functional receptor by diverse strains of feline immunodeficiency virus. J. Virol. 2006, 80, 3386–3394. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, J.; Griffiths, J.; Tews, I.; Cragg, M.S. Ox40: Structure and function—What questions remain? Mol. Immunol. 2017, 83, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Troyer, J.L.; Vandewoude, S.; Pecon-Slattery, J.; McIntosh, C.; Franklin, S.; Antunes, A.; Johnson, W.; O’Brien, S.J. FIV cross-species transmission: An evolutionary prospective. Vet. Immunol. Immunopathol. 2008, 123, 159–166. [Google Scholar] [CrossRef] [PubMed]
- VandeWoude, S.; Troyer, J.; Poss, M. Restrictions to cross-species transmission of lentiviral infection gleaned from studies of FIV. Vet. Immunol. Immunopathol. 2010, 134, 25–32. [Google Scholar] [CrossRef] [PubMed]
- VandeWoude, S.; Apetrei, C. Going wild: Lessons from naturally occurring T-lymphotropic lentiviruses. Clin. Microbiol. Rev. 2006, 19, 728–762. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.A.; O’Brien, S.J. Coadaptation and immunodeficiency virus: Lessons from the felidae. Curr. Opin. Genet. Dev. 1995, 5, 739–745. [Google Scholar] [CrossRef]
- O’Brien, S.J.; Troyer, J.L.; Brown, M.A.; Johnson, W.E.; Antunes, A.; Roelke, M.E.; Pecon-Slattery, J. Emerging viruses in the Felidae: Shifting paradigms. Viruses 2012, 4, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Pecon-Slattery, J.; Troyer, J.L.; Johnson, W.E.; O’Brien, S.J. Evolution of feline immunodeficiency virus in felidae: Implications for human health and wildlife ecology. Vet. Immunol. Immunopathol. 2008, 123, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Troyer, J.L.; Pecon-Slattery, J.; Roelke, M.E.; Johnson, W.; VandeWoude, S.; Vazquez-Salat, N.; Brown, M.; Frank, L.; Woodroffe, R.; Winterbach, C.; et al. Seroprevalence and genomic divergence of circulating strains of feline immunodeficiency virus among felidae and hyaenidae species. J. Virol. 2005, 79, 8282–8294. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Goto, Y.; Yoneda, K.; Endo, Y.; Mizuno, T.; Hamachi, M.; Maruyama, H.; Kinoshita, H.; Koga, S.; Komori, M.; et al. Interspecies transmission of feline immunodeficiency virus from the domestic cat to the tsushima cat (Felis bengalensis euptilura) in the wild. J. Virol. 1999, 73, 7916–7921. [Google Scholar] [PubMed]
- Lee, J.; Malmberg, J.L.; Wood, B.A.; Hladky, S.; Troyer, R.; Roelke, M.; Cunningham, M.; McBride, R.; Vickers, W.; Boyce, W.; et al. Feline immunodeficiency virus cross-species transmission: Implications for emergence of new lentiviral infections. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.A.; Brown, E.W.; Culver, M.; Johnson, W.E.; Pecon-Slattery, J.; Brousset, D.; O’Brien, S.J. Genetic and phylogenetic divergence of feline immunodeficiency virus in the puma (Puma concolor). J. Virol. 1996, 70, 6682–6693. [Google Scholar] [PubMed]
- Lee, J.S.; Bevins, S.N.; Serieys, L.E.; Vickers, W.; Logan, K.A.; Aldredge, M.; Boydston, E.E.; Lyren, L.M.; McBride, R.; Roelke-Parker, M.; et al. Evolution of puma lentivirus in bobcats (Lynx rufus) and mountain lions (Puma concolor) in North America. J. Virol. 2014, 88, 7727–7737. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Münk, C. Cellular restriction factors of feline immunodeficiency virus. Viruses 2011, 3, 1986–2005. [Google Scholar] [CrossRef] [PubMed]
- Doyle, T.; Goujon, C.; Malim, M.H. HIV-1 and interferons: Who’s interfering with whom? Nat. Rev. Microbiol. 2015, 13, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.; Chande, A.; Ziglio, S.; De Sanctis, V.; Bertorelli, R.; Goh, S.L.; McCauley, S.M.; Nowosielska, A.; Antonarakis, S.E.; Luban, J.; et al. HIV-1 NEF promotes infection by excluding SERINC5 from virion incorporation. Nature 2015, 526, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Usami, Y.; Wu, Y.; Gottlinger, H.G. SERINC3 and SERINC5 restrict HIV-1 infectivity and are counteracted by NEF. Nature 2015, 526, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, E.E.; Miyoshi, H.; Nagai, Y.; Shioda, T. A specific region of 37 amino acid residues in the spry (b30.2) domain of African green monkey TRIM5Α determines species-specific restriction of simian immunodeficiency virus sivmac infection. J. Virol. 2005, 79, 8870–8877. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Kuebler, P.J.; Heymann, J.J.; Sheehy, M.; Ortiz, G.M.; Ogg, G.; Barbour, J.D.; Lenz, J.; Steinfeld, A.D.; Nixon, D.F. Detection of T lymphocytes specific for human endogenous retrovirus K (HERV-K) in patients with seminoma. AIDS Res. Hum. Retrovir. 2006, 22, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; Stremlau, M.; Yuan, W.; Song, B.; Perron, M.; Sodroski, J. Functional replacement of the ring, B-box 2, and coiled-coil domains of tripartite motif 5α (TRIM5Α) by heterologous trim domains. J. Virol. 2006, 80, 6198–6206. [Google Scholar] [CrossRef] [PubMed]
- Saenz, D.T.; Teo, W.; Olsen, J.C.; Poeschla, E.M. Restriction of feline immunodeficiency virus by REF1, LV1, and primate TRIM5Α proteins. J. Virol. 2005, 79, 15175–15188. [Google Scholar] [CrossRef] [PubMed]
- Ozato, K.; Shin, D.M.; Chang, T.H.; Morse, H.C., 3rd. Trim family proteins and their emerging roles in innate immunity. Nat. Rev. Immunol. 2008, 8, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Mandell, M.A.; Kimura, T.; Jain, A.; Johansen, T.; Deretic, V. TRIM proteins regulate autophagy: TRIM5 is a selective autophagy receptor mediating HIV-1 restriction. Autophagy 2014, 10, 2387–2388. [Google Scholar] [CrossRef] [PubMed]
- Mandell, M.A.; Jain, A.; Arko-Mensah, J.; Chauhan, S.; Kimura, T.; Dinkins, C.; Silvestri, G.; Munch, J.; Kirchhoff, F.; Simonsen, A.; et al. Trim proteins regulate autophagy and can target autophagic substrates by direct recognition. Dev. Cell 2014, 30, 394–409. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.S.; Dudley, J.P. Apobecs and virus restriction. Virology 2015, 479–480, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, D.; Zhu, M.; Shen, Y.; Wei, W.; Ying, S.; Korner, H.; Li, J. Roles of SAMHD1 in antiviral defense, autoimmunity and cancer. Rev. Med. Virol. 2017, 27. [Google Scholar] [CrossRef] [PubMed]
- Kane, M.; Yadav, S.S.; Bitzegeio, J.; Kutluay, S.B.; Zang, T.; Wilson, S.J.; Schoggins, J.W.; Rice, C.M.; Yamashita, M.; Hatziioannou, T.; et al. MX2 is an interferon-induced inhibitor of HIV-1 infection. Nature 2013, 502, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Goujon, C.; Moncorge, O.; Bauby, H.; Doyle, T.; Ward, C.C.; Schaller, T.; Hue, S.; Barclay, W.S.; Schulz, R.; Malim, M.H. Human MX2 is an interferon-induced post-entry inhibitor of HIV-1 infection. Nature 2013, 502, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Neil, S.J. The antiviral activities of tetherin. Curr. Top. Microbiol. Immunol. 2013, 371, 67–104. [Google Scholar] [PubMed]
- McEwan, W.A.; Schaller, T.; Ylinen, L.M.; Hosie, M.J.; Towers, G.J.; Willett, B.J. Truncation of TRIM5 in the feliformia explains the absence of retroviral restriction in cells of the domestic cat. J. Virol. 2009, 83, 8270–8275. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, I.; Macintyre, A.; McMonagle, E.; Price, A.J.; James, L.C.; McEwan, W.A.; Hosie, M.J.; Willett, B.J. Potent lentiviral restriction by a synthetic feline TRIM5 cyclophilin a fusion. J. Virol. 2010, 84, 8980–8985. [Google Scholar] [CrossRef] [PubMed]
- Busnadiego, I.; Kane, M.; Rihn, S.J.; Preugschas, H.F.; Hughes, J.; Blanco-Melo, D.; Strouvelle, V.P.; Zang, T.M.; Willett, B.J.; Boutell, C.; et al. Host and viral determinants of MX2 antiretroviral activity. J. Virol. 2014, 88, 7738–7752. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Young, J.M.; Emerman, M.; Malik, H.S. Evolutionary analyses suggest a function of MXB immunity proteins beyond lentivirus restriction. PLoS Pathog. 2015, 11, e1005304. [Google Scholar] [CrossRef] [PubMed]
- Asadian, P.; Finnie, G.; Bienzle, D. The expression profile of sterile α motif and histidine-aspartate domain-containing protein 1 (SAMHD1) in feline tissues. Vet. Immunol. Immunopathol. 2018, 195, 7–18. [Google Scholar] [CrossRef] [PubMed]
- White, T.E.; Brandariz-Nunez, A.; Valle-Casuso, J.C.; Amie, S.; Nguyen, L.; Kim, B.; Brojatsch, J.; Diaz-Griffero, F. Contribution of SAM and HD domains to retroviral restriction mediated by human SAMHD1. Virology 2013, 436, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.H.; Guevara, R.B.; Marcano, A.C.; Saenz, D.T.; Fadel, H.J.; Rogstad, D.K.; Poeschla, E.M. Feline immunodeficiency virus envelope glycoproteins antagonize tetherin through a distinctive mechanism that requires virion incorporation. J. Virol. 2014, 88, 3255–3272. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, I.; McMonagle, E.L.; Petit, S.J.; Vijayakrishnan, S.; Logan, N.; Chan, C.N.; Towers, G.J.; Hosie, M.J.; Willett, B.J. Feline tetherin efficiently restricts release of feline immunodeficiency virus but not spreading of infection. J. Virol. 2011, 85, 5840–5852. [Google Scholar] [CrossRef] [PubMed]
- Celestino, M.; Calistri, A.; Del Vecchio, C.; Salata, C.; Chiuppesi, F.; Pistello, M.; Borsetti, A.; Palu, G.; Parolin, C. Feline tetherin is characterized by a short N-terminal region and is counteracted by the feline immunodeficiency virus envelope glycoprotein. J. Virol. 2012, 86, 6688–6700. [Google Scholar] [CrossRef] [PubMed]
- Münk, C.; Beck, T.; Zielonka, J.; Hotz-Wagenblatt, A.; Chareza, S.; Battenberg, M.; Thielebein, J.; Cichutek, K.; Bravo, I.G.; O’Brien, S.J.; et al. Functions, structure, and read-through alternative splicing of feline APOBEC3 genes. Genome Biol. 2008, 9, R48. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Marino, D.; Hofmann, H.; Yuhki, N.; Löchelt, M.; Münk, C. VIF of feline immunodeficiency virus from domestic cats protects against APOBEC3 restriction factors from many felids. J. Virol. 2010, 84, 7312–7324. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Zhang, Z.; Cano Ortiz, L.; Franco, A.C.; Häussinger, D.; Münk, C. Feline immunodeficiency virus vif N-terminal residues selectively counteract feline APOBEC3s. J. Virol. 2016, 90, 10545–10557. [Google Scholar] [CrossRef] [PubMed]
- Gabuzda, D.H.; Lawrence, K.; Langhoff, E.; Terwilliger, E.; Dorfman, T.; Haseltine, W.A.; Sodroski, J. Role of VIF in replication of human immunodeficiency virus type 1 in CD4+ T lymphocytes. J. Virol. 1992, 66, 6489–6495. [Google Scholar] [PubMed]
- Gabuzda, D.H.; Li, H.; Lawrence, K.; Vasir, B.S.; Crawford, K.; Langhoff, E. Essential role of VIF in establishing productive HIV-1 infection in peripheral blood T lymphocytes and monocyte/macrophages. J. Acquir. Immune Defic. Syndr. 1994, 7, 908–915. [Google Scholar] [PubMed]
- Gibbs, J.S.; Regier, D.A.; Desrosiers, R.C. Construction and in vitro properties of HIV-1 mutants with deletions in “nonessential” genes. AIDS Res. Hum. Retrovir. 1994, 10, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Strebel, K.; Daugherty, D.; Clouse, K.; Cohen, D.; Folks, T.; Martin, M.A. The HIV ‘a’ (SOR) gene product is essential for virus infectivity. Nature 1987, 328, 728–730. [Google Scholar] [CrossRef] [PubMed]
- Michaels, F.H.; Hattori, N.; Gallo, R.C.; Franchini, G. The human immunodeficiency virus type 1 (HIV-1) VIF protein is located in the cytoplasm of infected cells and its effect on viral replication is equivalent in HIV-2. AIDS Res. Hum. Retrovir. 1993, 9, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Shibata, R.; Adachi, A.; Sakai, H.; Ishimoto, A.; Miura, T.; Hayami, M. Mutational analysis of simian immunodeficiency virus from African green monkeys and human immunodeficiency virus type 2. J. Med. Primatol. 1990, 19, 217–225. [Google Scholar] [PubMed]
- Lockridge, K.M.; Himathongkham, S.; Sawai, E.T.; Chienand, M.; Sparger, E.E. The feline immunodeficiency virus VIF gene is required for productive infection of feline peripheral blood mononuclear cells and monocyte-derived macrophages. Virology 1999, 261, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Leutenegger, C.M.; Stefano Cole, K.; Pedersen, N.C.; Sparger, E.E. A feline immunodeficiency virus VIF-deletion mutant remains attenuated upon infection of newborn kittens. J. Gen. Virol. 2007, 88, 2793–2799. [Google Scholar] [CrossRef] [PubMed]
- Paul, T.A.; Casey, J.W.; Avery, R.J.; Sutton, C.A. Expression of feline immunodeficiency virus VIF is associated with reduced viral mutation rates without restoration of replication of VIF mutant viruses. Virology 2007, 361, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.N.; Holmes, R.K.; Sheehy, A.M.; Malim, M.H. APOBEC-mediated editing of viral RNA. Science 2004, 305, 645. [Google Scholar] [CrossRef] [PubMed]
- LaRue, R.S.; Andresdottir, V.; Blanchard, Y.; Conticello, S.G.; Derse, D.; Emerman, M.; Greene, W.C.; Jonsson, S.R.; Landau, N.R.; Löchelt, M.; et al. Guidelines for naming nonprimate APOBEC3 genes and proteins. J. Virol. 2009, 83, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.; Bloch, N.; Landau, N.R. Intrinsic host restrictions to HIV-1 and mechanisms of viral escape. Nat. Immunol. 2015, 16, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Münk, C.; Willemsen, A.; Bravo, I.G. An ancient history of gene duplications, fusions and losses in the evolution of APOBEC3 mutators in mammals. BMC Evol. Biol. 2012, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Huthoff, H.; Malim, M.H. Identification of amino acid residues in APOBEC3g required for regulation by human immunodeficiency virus type 1 VIF and virion encapsidation. J. Virol. 2007, 81, 3807–3815. [Google Scholar] [CrossRef] [PubMed]
- Apolonia, L.; Schulz, R.; Curk, T.; Rocha, P.; Swanson, C.M.; Schaller, T.; Ule, J.; Malim, M.H. Promiscuous rna binding ensures effective encapsidation of APOBEC3 proteins by HIV-1. PLoS Pathog. 2015, 11, e1004609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafer, A.; Bogerd, H.P.; Cullen, B.R. Specific packaging of APOBEC3g into HIV-1 virions is mediated by the nucleocapsid domain of the gag polyprotein precursor. Virology 2004, 328, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Alce, T.M.; Popik, W. APOBEC3g is incorporated into virus-like particles by a direct interaction with HIV-1 gag nucleocapsid protein. J. Biol. Chem. 2004, 279, 34083–34086. [Google Scholar] [CrossRef] [PubMed]
- Zennou, V.; Perez-Caballero, D.; Gottlinger, H.; Bieniasz, P.D. APOBEC3g incorporation into human immunodeficiency virus type 1 particles. J. Virol. 2004, 78, 12058–12061. [Google Scholar] [CrossRef] [PubMed]
- Svarovskaia, E.S.; Xu, H.; Mbisa, J.L.; Barr, R.; Gorelick, R.J.; Ono, A.; Freed, E.O.; Hu, W.S.; Pathak, V.K. Human apolipoprotein b mRNA-editing enzyme-catalytic polypeptide-like 3g (APOBEC3g) is incorporated into HIV-1 virions through interactions with viral and nonviral RNAs. J. Biol. Chem. 2004, 279, 35822–35828. [Google Scholar] [CrossRef] [PubMed]
- Mangeat, B.; Turelli, P.; Caron, G.; Friedli, M.; Perrin, L.; Trono, D. Broad antiretroviral defence by human APOBEC3g through lethal editing of nascent reverse transcripts. Nature 2003, 424, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Refsland, E.W.; Hultquist, J.F.; Harris, R.S. Endogenous origins of HIV-1 G-to-A hypermutation and restriction in the nonpermissive T cell line CEM2n. PLoS Pathog. 2012, 8, e1002800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, B.; Pomerantz, R.J.; Zhang, C.; Arunachalam, S.C.; Gao, L. The cytidine deaminase CEM15 induces hypermutation in newly synthesized HIV-1 DNA. Nature 2003, 424, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Mariani, R.; Chen, D.; Schröfelbauer, B.; Navarro, F.; König, R.; Bollman, B.; Münk, C.; Nymark-McMahon, H.; Landau, N.R. Species-specific exclusion of APOBEC3g from HIV-1 virions by VIF. Cell 2003, 114, 21–31. [Google Scholar] [CrossRef]
- Yang, B.; Chen, K.; Zhang, C.; Huang, S.; Zhang, H. Virion-associated uracil DNA glycosylase-2 and apurinic/apyrimidinic endonuclease are involved in the degradation of APOBEC3g-edited nascent HIV-1 DNA. J. Biol. Chem. 2007, 282, 11667–11675. [Google Scholar] [CrossRef] [PubMed]
- Gillick, K.; Pollpeter, D.; Phalora, P.; Kim, E.Y.; Wolinsky, S.M.; Malim, M.H. Suppression of HIV-1 infection by APOBEC3 proteins in primary human CD4(+) T cells is associated with inhibition of processive reverse transcription as well as excessive cytidine deamination. J. Virol. 2013, 87, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Iwatani, Y.; Chan, D.S.; Wang, F.; Maynard, K.S.; Sugiura, W.; Gronenborn, A.M.; Rouzina, I.; Williams, M.C.; Musier-Forsyth, K.; Levin, J.G. Deaminase-independent inhibition of HIV-1 reverse transcription by APOBEC3g. Nucleic Acids Res. 2007, 35, 7096–7108. [Google Scholar] [CrossRef] [PubMed]
- Holmes, R.K.; Koning, F.A.; Bishop, K.N.; Malim, M.H. APOBEC3f can inhibit the accumulation of HIV-1 reverse transcription products in the absence of hypermutation. Comparisons with APOBEC3g. J. Biol. Chem. 2007, 282, 2587–2595. [Google Scholar] [CrossRef] [PubMed]
- Mbisa, J.L.; Barr, R.; Thomas, J.A.; Vandegraaff, N.; Dorweiler, I.J.; Svarovskaia, E.S.; Brown, W.L.; Mansky, L.M.; Gorelick, R.J.; Harris, R.S.; et al. Human immunodeficiency virus type 1 CDNAS produced in the presence of APOBEC3g exhibit defects in plus-strand DNA transfer and integration. J. Virol. 2007, 81, 7099–7110. [Google Scholar] [CrossRef] [PubMed]
- Mbisa, J.L.; Bu, W.; Pathak, V.K. APOBEC3f and APOBEC3g inhibit HIV-1 DNA integration by different mechanisms. J. Virol. 2010, 84, 5250–5259. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ao, Z.; Chen, L.; Kobinger, G.; Peng, J.; Yao, X. The cellular antiviral protein APOBEC3g interacts with HIV-1 reverse transcriptase and inhibits its function during viral replication. J. Virol. 2012, 86, 3777–3786. [Google Scholar] [CrossRef] [PubMed]
- Pollpeter, D.; Parsons, M.; Sobala, A.E.; Coxhead, S.; Lang, R.D.; Bruns, A.M.; Papaioannou, S.; McDonnell, J.M.; Apolonia, L.; Chowdhury, J.A.; et al. Deep sequencing of HIV-1 reverse transcripts reveals the multifaceted antiviral functions of APOBEC3g. Nat. Microbiol. 2017, 3, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Troyer, R.M.; Thompson, J.; Elder, J.H.; VandeWoude, S. Accessory genes confer a high replication rate to virulent feline immunodeficiency virus. J. Virol. 2013, 87, 7940–7951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gu, Q.; Jaguva Vasudevan, A.A.; Hain, A.; Kloke, B.P.; Hasheminasab, S.; Mulnaes, D.; Sato, K.; Cichutek, K.; Häussinger, D.; et al. Determinants of FIV and HIV VIF sensitivity of feline APOBEC3 restriction factors. Retrovirology 2016, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Löchelt, M.; Romen, F.; Bastone, P.; Muckenfuss, H.; Kirchner, N.; Kim, Y.B.; Truyen, U.; Rosler, U.; Battenberg, M.; Saib, A.; et al. The antiretroviral activity of APOBEC3 is inhibited by the foamy virus accessory bet protein. Proc. Natl. Acad. Sci. USA 2005, 102, 7982–7987. [Google Scholar] [CrossRef] [PubMed]
- Münk, C.; Zielonka, J.; Constabel, H.; Kloke, B.P.; Rengstl, B.; Battenberg, M.; Bonci, F.; Pistello, M.; Löchelt, M.; Cichutek, K. Multiple restrictions of human immunodeficiency virus type 1 in feline cells. J. Virol. 2007, 81, 7048–7060. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.A.; Hu, C.; Saenz, D.T.; Fadel, H.J.; Sims, O.; Peretz, M.; Poeschla, E.M. Productive replication of vif-chimeric HIV-1 in feline cells. J. Virol. 2010, 84, 7378–7395. [Google Scholar] [CrossRef] [PubMed]
- Larue, R.S.; Lengyel, J.; Jonsson, S.R.; Andresdottir, V.; Harris, R.S. Lentiviral VIF degrades the APOBEC3z3/APOBEC3h protein of its mammalian host and is capable of cross-species activity. J. Virol. 2010, 84, 8193–8201. [Google Scholar] [CrossRef] [PubMed]
- Chatterji, U.; Grant, C.K.; Elder, J.H. Feline immunodeficiency virus VIF localizes to the nucleus. J. Virol. 2000, 74, 2533–2540. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, W.; Lv, M.; Zuo, T.; Kong, W.; Yu, X. Identification of a cullin5-elonginb-elonginc E3 complex in degradation of feline immunodeficiency virus vif-mediated feline APOBEC3 proteins. J. Virol. 2011, 85, 12482–12491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, J.; Evans, S.L.; Yu, Y.; Yu, X.F. T-cell differentiation factor CBF-β regulates HIV-1 VIF-mediated evasion of host restriction. Nature 2012, 481, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Jager, S.; Kim, D.Y.; Hultquist, J.F.; Shindo, K.; LaRue, R.S.; Kwon, E.; Li, M.; Anderson, B.D.; Yen, L.; Stanley, D.; et al. VIF hijacks CBF-β to degrade APOBEC3g and promote HIV-1 infection. Nature 2012, 481, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.R.; Stanley, D.J.; Hultquist, J.F.; Johnson, J.R.; Mietrach, N.; Binning, J.M.; Jonsson, S.R.; Barelier, S.; Newton, B.W.; Johnson, T.L.; et al. Lineage-specific viral hijacking of non-canonical e3 ubiquitin ligase cofactors in the evolution of vif anti-APOBEC3 activity. Cell Rep. 2015, 11, 1236–1250. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Zhu, D.; Wang, C.; Su, C.; Ma, J.; Ma, J.; Wang, X. Core-binding factor subunit β is not required for non-primate lentiviral VIF-mediated APOBEC3 degradation. J. Virol. 2014, 88, 12112–12122. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liang, W.; Hua, D.; Zhou, X.; Du, J.; Evans, S.L.; Gao, Q.; Wang, H.; Viqueira, R.; Wei, W.; et al. Evolutionarily conserved requirement for core binding factor β in the assembly of the human immunodeficiency virus/simian immunodeficiency virus VIF-cullin 5-ring e3 ubiquitin ligase. J. Virol. 2014, 88, 3320–3328. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, R.; Takeuchi, J.S.; Yamada, E.; Nakano, Y.; Ren, F.; Tanaka, H.; Münk, C.; Harris, R.S.; Miyazawa, T.; Koyanagi, Y.; et al. VIF determines the requirement for CBF-β in APOBEC3 degradation. J. Gen. Virol. 2015, 96, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.S.; Anderson, B.D. Evolutionary paradigms from ancient and ongoing conflicts between the lentiviral VIF protein and mammalian APOBEC3 enzymes. PLoS Pathog. 2016, 12, e1005958. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, R.; Takeuchi, J.S.; Yamada, E.; Nakano, Y.; Misawa, N.; Kimura, Y.; Ren, F.; Miyazawa, T.; Koyanagi, Y.; Sato, K. Feline immunodeficiency virus evolutionarily acquires two proteins, VIF and protease, capable of antagonizing feline APOBEC3. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Pathak, V.K. Identification of specific determinants of human APOBEC3f, APOBEC3c, and APOBEC3de and african green monkey APOBEC3f that interact with HIV-1 VIF. J. Virol. 2010, 84, 12599–12608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gu, Q.; Jaguva Vasudevan, A.A.; Jeyaraj, M.; Schmidt, S.; Zielonka, J.; Perkovic, M.; Heckel, J.O.; Cichutek, K.; Häussinger, D.; et al. Vif proteins from diverse human immunodeficiency virus/simian immunodeficiency virus lineages have distinct binding sites in A3C. J. Virol. 2016, 90, 10193–10208. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Ode, H.; Nakashima, M.; Imahashi, M.; Naganawa, Y.; Kurosawa, T.; Yokomaku, Y.; Yamane, T.; Watanabe, N.; Suzuki, A.; et al. The APOBEC3c crystal structure and the interface for HIV-1 VIF binding. Nat. Struct. Mol. Biol. 2012, 19, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Tsuzuki, S.; Awazu, H.; Hamano, A.; Okada, A.; Ode, H.; Maejima, M.; Hachiya, A.; Yokomaku, Y.; Watanabe, N.; et al. Mapping region of human restriction factor APOBEC3h critical for interaction with HIV-1 VIF. J. Mol. Biol. 2017, 429, 1262–1276. [Google Scholar] [CrossRef] [PubMed]
- Salter, J.D.; Morales, G.A.; Smith, H.C. Structural insights for HIV-1 therapeutic strategies targeting VIF. Trends Biochem. Sci. 2014, 39, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, R.; Izumi, T.; Yamada, E.; Nakano, Y.; Misawa, N.; Ren, F.; Carpenter, M.A.; Ikeda, T.; Münk, C.; Harris, R.S.; et al. A naturally occurring domestic cat APOBEC3 variant confers resistance to feline immunodeficiency virus infection. J. Virol. 2016, 90, 474–485. [Google Scholar] [CrossRef] [PubMed]
- De Castro, F.L.; Junqueira, D.M.; de Medeiros, R.M.; da Silva, T.R.; Costenaro, J.G.; Knak, M.B.; de Matos Almeida, S.E.; Campos, F.S.; Roehe, P.M.; Franco, A.C. Analysis of single-nucleotide polymorphisms in the APOBEC3h gene of domestic cats (Felis catus) and their association with the susceptibility to feline immunodeficiency virus and feline leukemia virus infections. Infect. Genet. Evol. 2014, 27, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.; Emerman, M. Polymorphism in human APOBEC3h affects a phenotype dominant for subcellular localization and antiviral activity. J. Virol. 2011, 85, 8197–8207. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Abudu, A.; Son, S.; Dang, Y.; Venta, P.J.; Zheng, Y.H. Analysis of human APOBEC3h haplotypes and anti-human immunodeficiency virus type 1 activity. J. Virol. 2011, 85, 3142–3152. [Google Scholar] [CrossRef] [PubMed]
- Harari, A.; Ooms, M.; Mulder, L.C.; Simon, V. Polymorphisms and splice variants influence the antiretroviral activity of human APOBEC3h. J. Virol. 2009, 83, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.; Wu, L.I.; Emerman, M. The range of human APOBEC3h sensitivity to lentiviral VIF proteins. J. Virol. 2010, 84, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Binka, M.; Ooms, M.; Steward, M.; Simon, V. The activity spectrum of vif from multiple HIV-1 subtypes against APOBEC3g, APOBEC3f, and APOBEC3h. J. Virol. 2012, 86, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Ooms, M.; Brayton, B.; Letko, M.; Maio, S.M.; Pilcher, C.D.; Hecht, F.M.; Barbour, J.D.; Simon, V. HIV-1 VIF adaptation to human APOBEC3h haplotypes. Cell Host Microbe 2013, 14, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Refsland, E.W.; Hultquist, J.F.; Luengas, E.M.; Ikeda, T.; Shaban, N.M.; Law, E.K.; Brown, W.L.; Reilly, C.; Emerman, M.; Harris, R.S. Natural polymorphisms in human APOBEC3h and HIV-1 VIF combine in primary t lymphocytes to affect viral g-to-a mutation levels and infectivity. PLoS Genet. 2014, 10, e1004761. [Google Scholar] [CrossRef] [PubMed]
- OhAinle, M.; Kerns, J.A.; Malik, H.S.; Emerman, M. Adaptive evolution and antiviral activity of the conserved mammalian cytidine deaminase APOBEC3h. J. Virol. 2006, 80, 3853–3862. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Siew, L.M.; Wang, X.; Han, Y.; Lampen, R.; Zheng, Y.H. Human cytidine deaminase APOBEC3h restricts HIV-1 replication. J. Biol. Chem. 2008, 283, 11606–11614. [Google Scholar] [CrossRef] [PubMed]
- OhAinle, M.; Kerns, J.A.; Li, M.M.; Malik, H.S.; Emerman, M. Antiretroelement activity of APOBEC3h was lost twice in recent human evolution. Cell Host Microbe 2008, 4, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Naruse, T.K.; Sakurai, D.; Ohtani, H.; Sharma, G.; Sharma, S.K.; Vajpayee, M.; Mehra, N.K.; Kaur, G.; Kimura, A. APOBEC3h polymorphisms and susceptibility to HIV-1 infection in an Indian population. J. Hum. Genet. 2016, 61, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, D.; Iwatani, Y.; Ohtani, H.; Naruse, T.K.; Terunuma, H.; Sugiura, W.; Kimura, A. APOBEC3h polymorphisms associated with the susceptibility to HIV-1 infection and aids progression in Japanese. Immunogenetics 2015, 67, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gu, Q.; de Manuel Montero, M.; Bravo, I.G.; Marques-Bonet, T.; Häussinger, D.; Münk, C. Stably expressed APOBEC3h forms a barrier for cross-species transmission of simian immunodeficiency virus of chimpanzee to humans. PLoS Pathog. 2017, 13, e1006746. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Zhang, Z.; Gertzen, C.G.W.; Häussinger, D.; Gohlke, H.; Münk, C. Identification of a conserved interface of HIV-1 and FIV Vifs with cullin 5. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Chareza, S.; Slavkovic Lukic, D.; Liu, Y.; Rathe, A.M.; Munk, C.; Zabogli, E.; Pistello, M.; Lochelt, M. Molecular and functional interactions of cat APOBEC3 and feline foamy and immunodeficiency virus proteins: Different ways to counteract host-encoded restriction. Virology 2012, 424, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Lukic, D.S.; Hotz-Wagenblatt, A.; Lei, J.; Rathe, A.M.; Muhle, M.; Denner, J.; Munk, C.; Lochelt, M. Identification of the feline foamy virus bet domain essential for APOBEC3 counteraction. Retrovirology 2013, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Herchenroder, O.; Renne, R.; Loncar, D.; Cobb, E.K.; Murthy, K.K.; Schneider, J.; Mergia, A.; Luciw, P.A. Isolation, cloning, and sequencing of simian foamy viruses from chimpanzees (SFVCPZ): High homology to human foamy virus (HFV). Virology 1994, 201, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.A.; Wiegand, H.L.; Moore, M.D.; Schafer, A.; McClure, M.O.; Cullen, B.R. Foamy virus bet proteins function as novel inhibitors of the APOBEC3 family of innate antiretroviral defense factors. J. Virol. 2005, 79, 8724–8731. [Google Scholar] [CrossRef] [PubMed]
- Jaguva Vasudevan, A.A.; Perkovic, M.; Bulliard, Y.; Cichutek, K.; Trono, D.; Häussinger, D.; Münk, C. Prototype foamy virus bet impairs the dimerization and cytosolic solubility of human APOBEC3g. J. Virol. 2013, 87, 9030–9040. [Google Scholar] [CrossRef] [PubMed]
- Perkovic, M.; Schmidt, S.; Marino, D.; Russell, R.A.; Stauch, B.; Hofmann, H.; Kopietz, F.; Kloke, B.P.; Zielonka, J.; Strover, H.; et al. Species-specific inhibition of APOBEC3c by the prototype foamy virus protein bet. J. Biol. Chem. 2009, 284, 5819–5826. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.W.; Brown, M.A.; Shindle, D.B.; Terrell, S.P.; Hayes, K.A.; Ferree, B.C.; McBride, R.T.; Blankenship, E.L.; Jansen, D.; Citino, S.B.; et al. Epizootiology and management of feline leukemia virus in the florida puma. J. Wildl. Dis. 2008, 44, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Terry, A.; Kilbey, A.; Naseer, A.; Levy, L.S.; Ahmad, S.; Watts, C.; Mackay, N.; Cameron, E.; Wilson, S.; Neil, J.C. Barriers to infection of human cells by feline leukemia virus: Insights into resistance to zoonosis. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, J.K.; Sparger, E.; Ho, E.W.; Andersen, P.R.; O’Connor, T.P.; Mandell, C.P.; Lowenstine, L.; Munn, R.; Pedersen, N.C. Pathogenesis of experimentally induced feline immunodeficiency virus infection in cats. Am. J. Vet. Res. 1988, 49, 1246–1258. [Google Scholar] [PubMed]
- Ueland, K.; Nesse, L.L. No evidence of vertical transmission of naturally acquired feline immunodeficiency virus infection. Vet. Immunol. Immunopathol. 1992, 33, 301–308. [Google Scholar] [CrossRef]
- Medeiros Sde, O.; Martins, A.N.; Dias, C.G.; Tanuri, A.; Brindeiro Rde, M. Natural transmission of feline immunodeficiency virus from infected queen to kitten. Virol. J. 2012, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- VandeWoude, S.; O’Brien, S.J.; Langelier, K.; Hardy, W.D.; Slattery, J.P.; Zuckerman, E.E.; Hoover, E.A. Growth of lion and puma lentiviruses in domestic cat cells and comparisons with FIV. Virology 1997, 233, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Troyer, J.L.; Roelke, M.E.; Jespersen, J.M.; Baggett, N.; Buckley-Beason, V.; MacNulty, D.; Craft, M.; Packer, C.; Pecon-Slattery, J.; O’Brien, S.J. FIV diversity: FIV PLE subtype composition may influence disease outcome in african lions. Vet. Immunol. Immunopathol. 2011, 143, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Franklin, S.P.; Troyer, J.L.; Terwee, J.A.; Lyren, L.M.; Boyce, W.M.; Riley, S.P.; Roelke, M.E.; Crooks, K.R.; Vandewoude, S. Frequent transmission of immunodeficiency viruses among bobcats and pumas. J. Virol. 2007, 81, 10961–10969. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, R.; Nakano, Y.; Yamada, E.; Izumi, T.; Misawa, N.; Koyanagi, Y.; Sato, K. Species-specific differences in the ability of feline lentiviral vif to degrade feline APOBEC3proteins. Microbiol. Immunol. 2016, 60, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Poss, M.; Ross, H.A.; Painter, S.L.; Holley, D.C.; Terwee, J.A.; Vandewoude, S.; Rodrigo, A. Feline lentivirus evolution in cross-species infection reveals extensive G-to-A mutation and selection on key residues in the viral polymerase. J. Virol. 2006, 80, 2728–2737. [Google Scholar] [CrossRef] [PubMed]
- Yamada, E.; Yoshikawa, R.; Nakano, Y.; Misawa, N.; Kobayashi, T.; Ren, F.; Izumi, T.; Miyazawa, T.; Koyanagi, Y.; Sato, K. A naturally occurring bovine APOBEC3 confers resistance to bovine lentiviruses: Implication for the co-evolution of bovids and their lentiviruses. Sci. Rep. 2016, 6, 33988. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Gu, Q.; Marino, D.; Lee, K.-L.; Kong, I.-K.; Häussinger, D.; Münk, C. Feline APOBEC3s, Barriers to Cross-Species Transmission of FIV? Viruses 2018, 10, 186. https://doi.org/10.3390/v10040186
Zhang Z, Gu Q, Marino D, Lee K-L, Kong I-K, Häussinger D, Münk C. Feline APOBEC3s, Barriers to Cross-Species Transmission of FIV? Viruses. 2018; 10(4):186. https://doi.org/10.3390/v10040186
Chicago/Turabian StyleZhang, Zeli, Qinyong Gu, Daniela Marino, Kyeong-Lim Lee, Il-Keun Kong, Dieter Häussinger, and Carsten Münk. 2018. "Feline APOBEC3s, Barriers to Cross-Species Transmission of FIV?" Viruses 10, no. 4: 186. https://doi.org/10.3390/v10040186
APA StyleZhang, Z., Gu, Q., Marino, D., Lee, K.-L., Kong, I.-K., Häussinger, D., & Münk, C. (2018). Feline APOBEC3s, Barriers to Cross-Species Transmission of FIV? Viruses, 10(4), 186. https://doi.org/10.3390/v10040186