Development and Validation of a Microtiter Plate-Based Assay for Determination of Bacteriophage Host Range and Virulence


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Bacteriophage Strains and Culture Conditions
2.3. Phage Host Range Agar Overlay Spot Assay
2.4. Methodology Development for the Microtiter Plate Host Range Assay
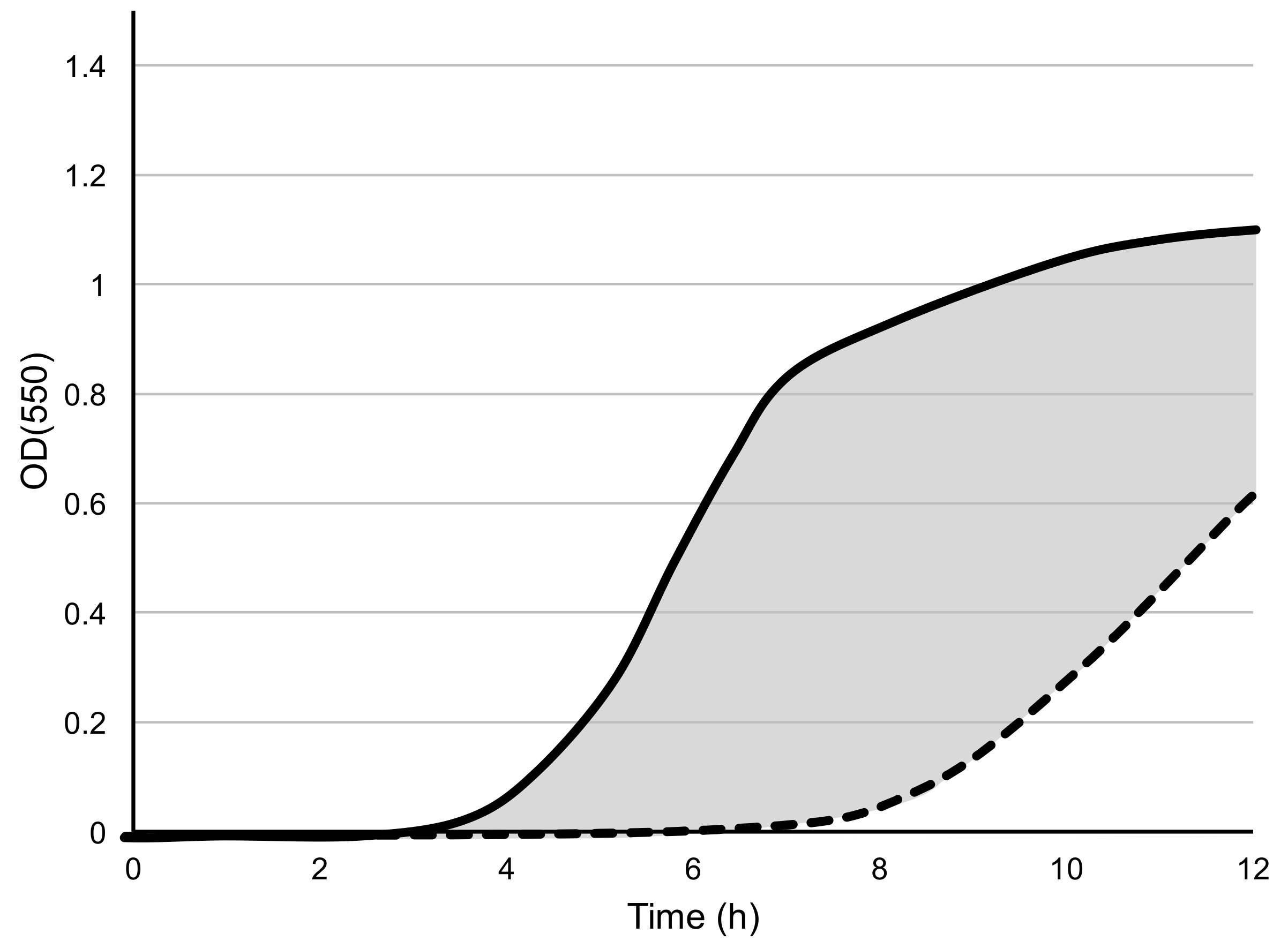
2.5. Analysis of Microtiter Plate Host Range Assay Data
2.6. Statistical Analysis
3. Results and Discussion
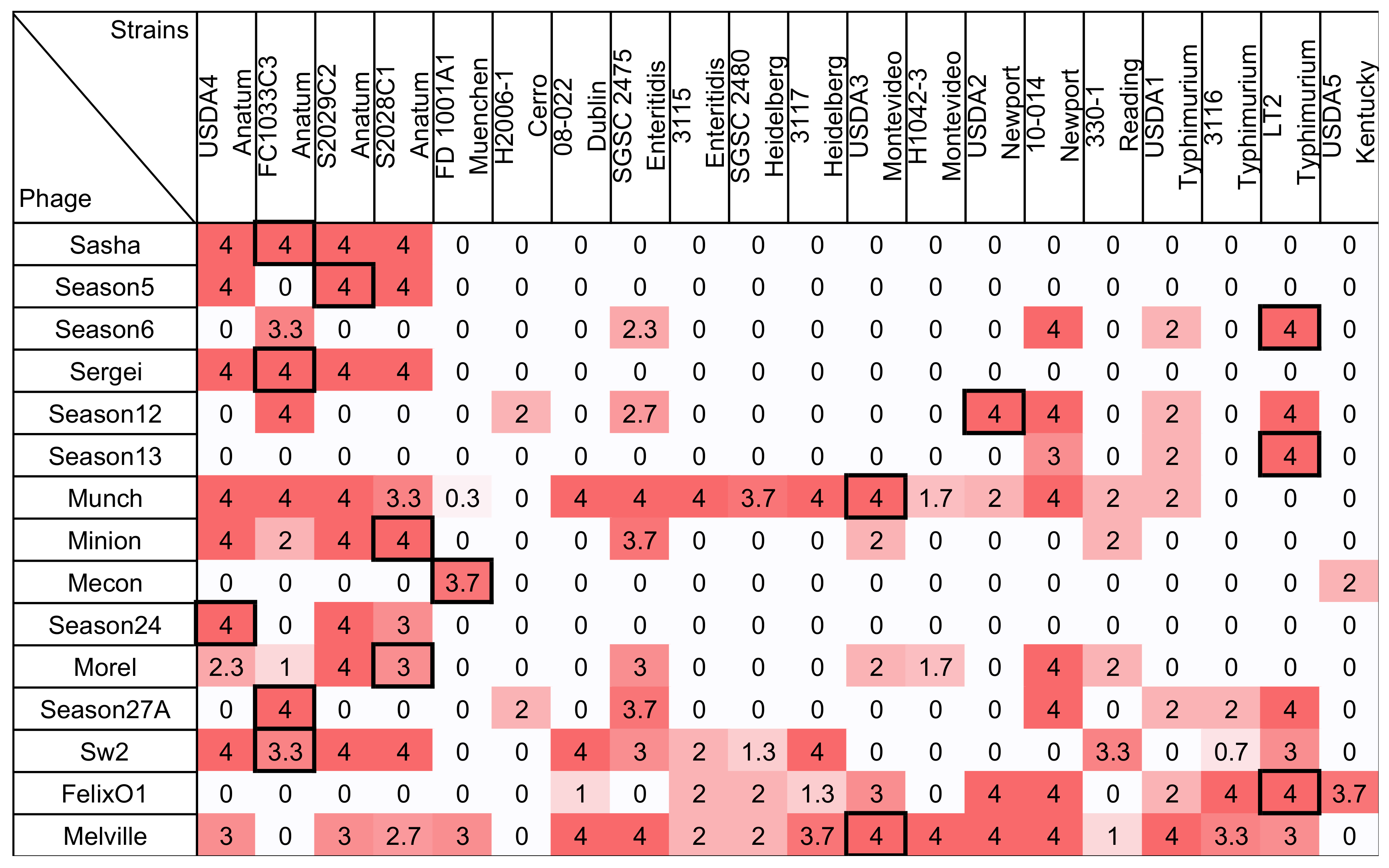
3.1. Measurement of Phage Host Range by Traditional Spot Assay
3.2. Determination of Microtiter Assay Parameters
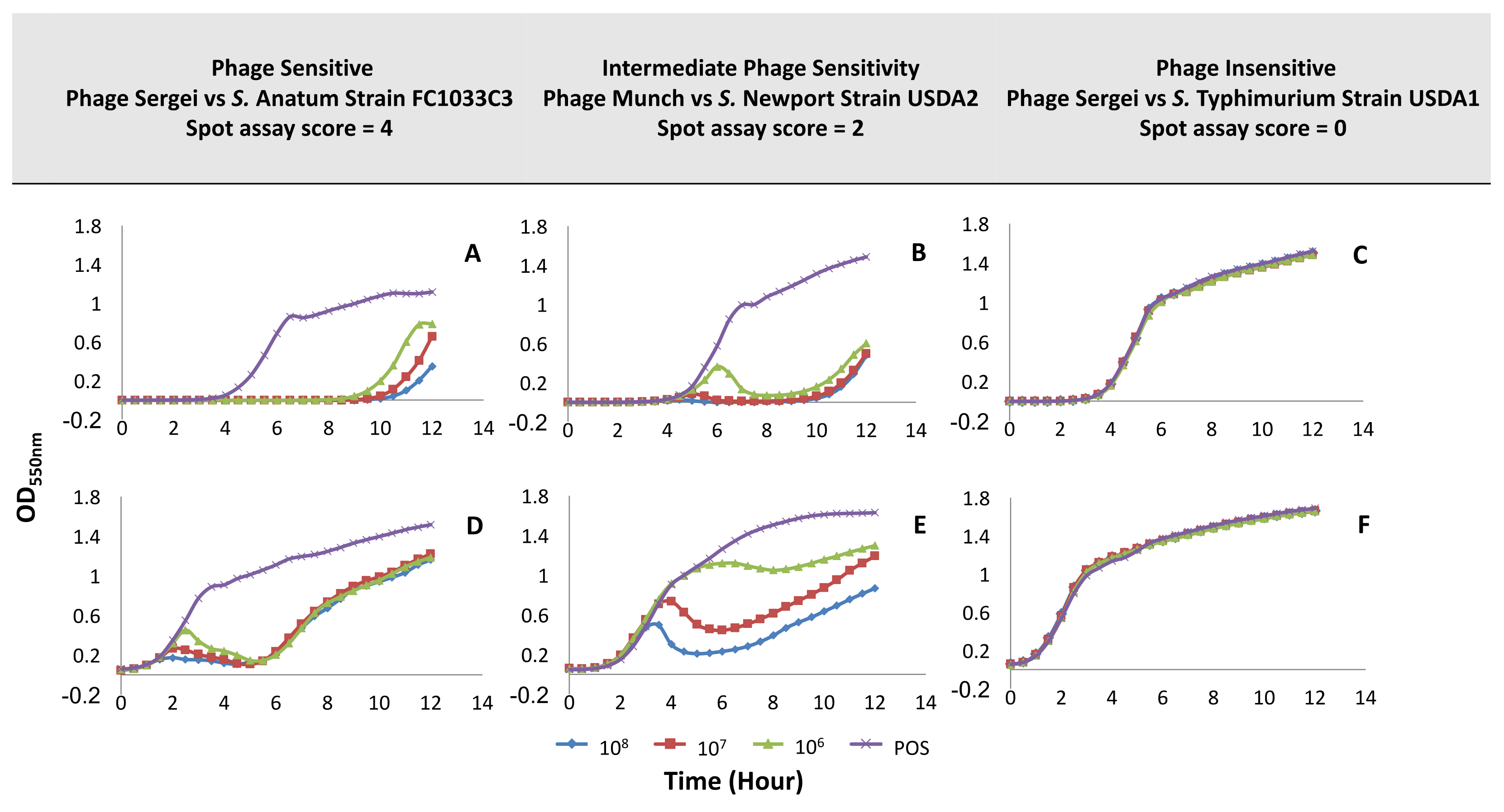
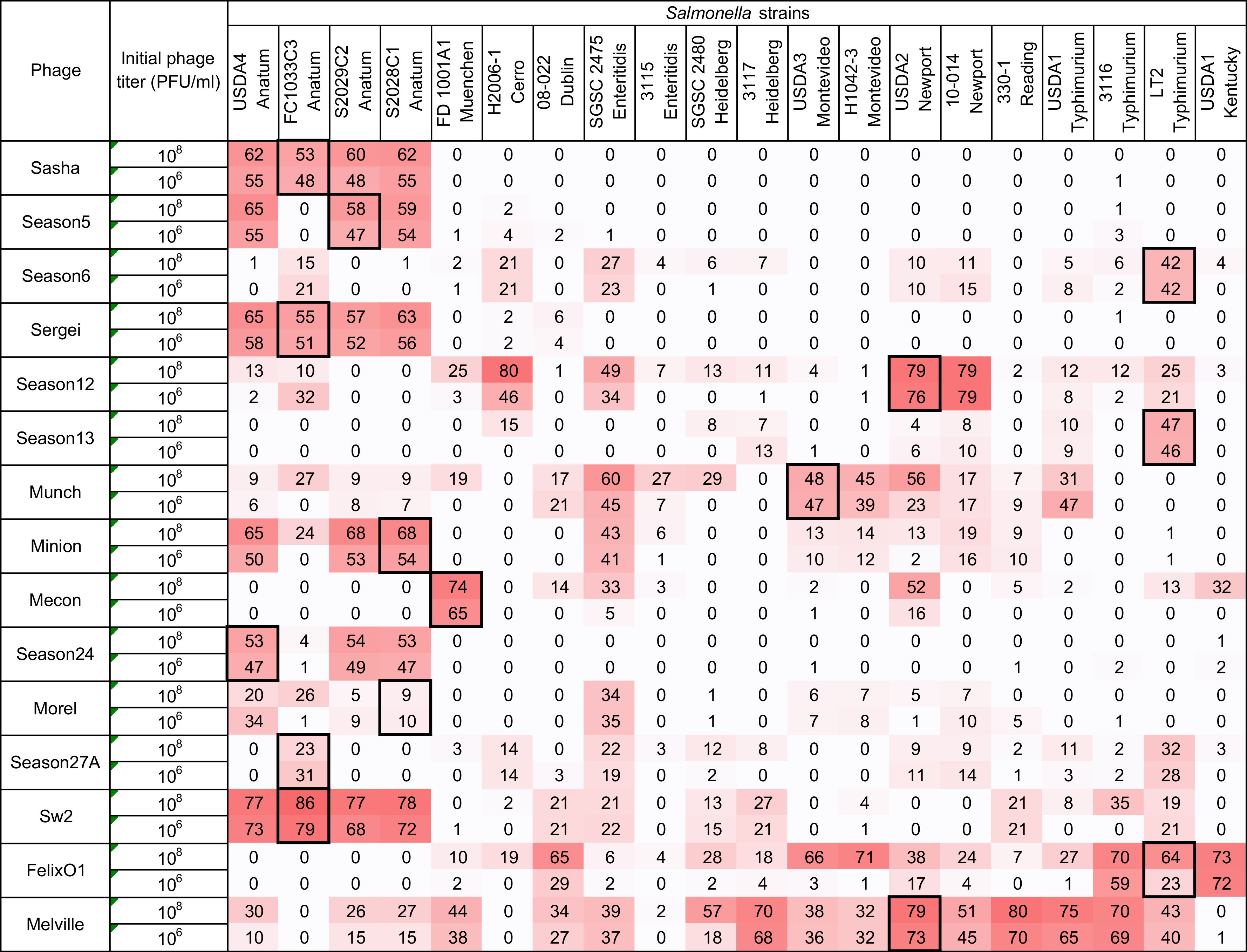
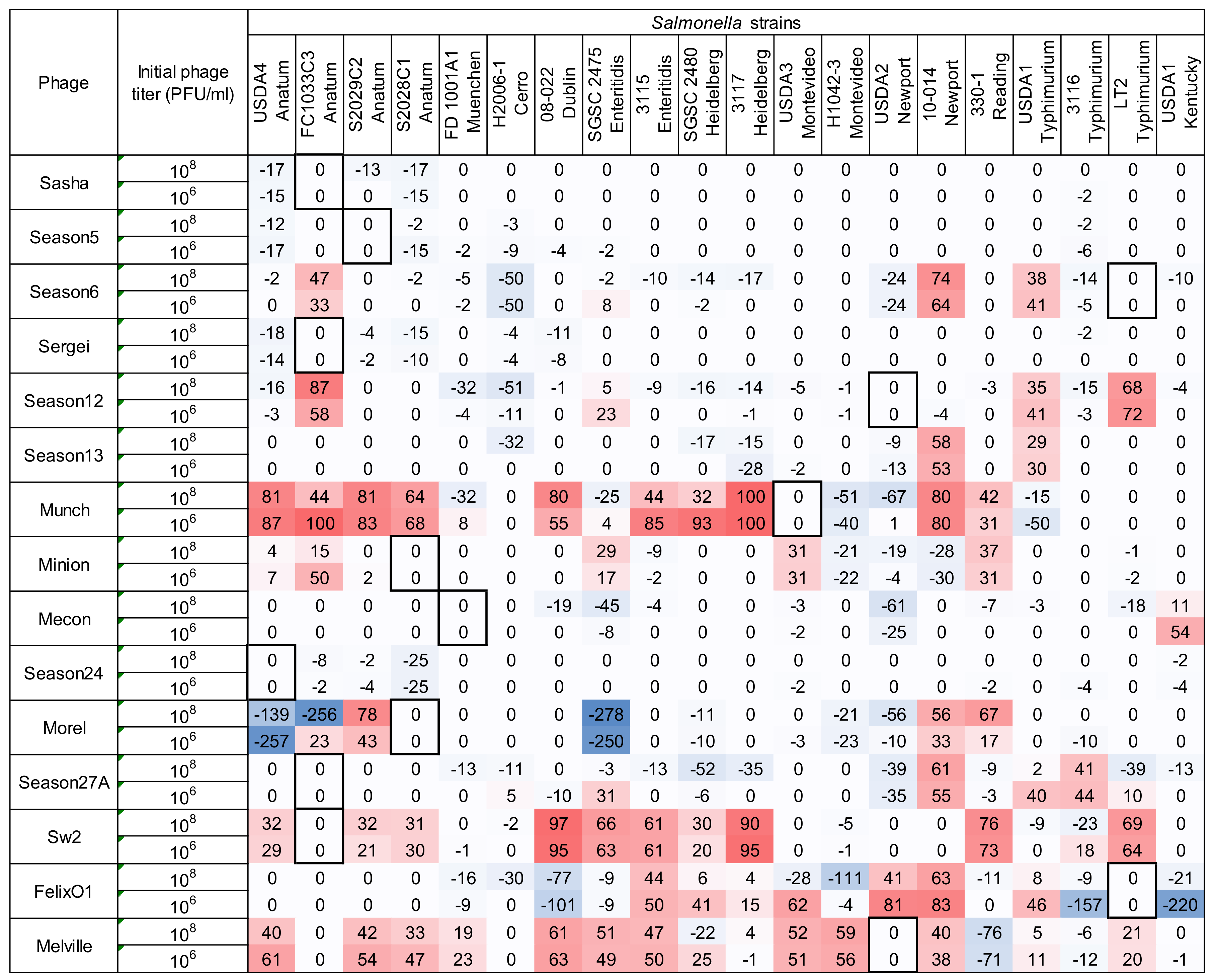
3.3. Measurement of Phage Host Range and Virulence by Microtiter Plate Assay
3.4. Comparison Between Two Host Range Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- The Centers for Disease Control and Prevention (CDC). National Antimicrobial Resistance Monitoring System for Enteric Bacteria (NARMS): Human Isolates Surveillance Report for 2014 (Final Report); Department of Health and Human Services, CDC: Atlanta, GA, USA, 2016.
- Grayson, M.L.; Heymann, D.; Pittet, D. The evolving threat of antimicrobial resistance introduction. In Evolving Threat of Antimicrobial Resistance: Options for Action; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Stanton, T.B. A call for antibiotic alternatives research. Trends Microbiol. 2013, 21, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Summers, W.C. Bacteriophage therapy. Annu. Rev. Microbiol. 2001, 55, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Morange, M. Felix d’Herelle and the origins of molecular biology. Hist. Philos. Life Sci. 2000, 22, 441–442. [Google Scholar]
- Kutter, E.M.; Kuhl, S.J.; Abedon, S.T. Re-establishing a place for phage therapy in western medicine. Future Microbiol. 2015, 10, 685–688. [Google Scholar] [CrossRef] [PubMed]
- De Groat, A.F. The bacteriophage: A method of isolation. J. Immunol. 1927, 14, 175–179. [Google Scholar]
- Chibani-Chennoufi, S.; Bruttin, A.; Dillmann, M.L.; Brussow, H. Phage-host interaction: An ecological perspective. J. Bacteriol. 2004, 186, 3677–3686. [Google Scholar] [CrossRef] [PubMed]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar] [PubMed]
- Skurnik, M.; Pajunen, M.; Kiljunen, S. Biotechnological challenges of phage therapy. Biotechnol. Lett. 2007, 29, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.S. Phage therapy-constraints and possibilities. Upsala J. Med. Sci. 2014, 119, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Debarbrieux, L. Phage therapy: Awakening a sleeping giant. Emerg. Top. Life Sci. 2017, 1, 93–103. [Google Scholar] [CrossRef]
- Henry, M.; Lavigne, R.; Debarbieux, L. Predicting in vivo efficacy of therapeutic bacteriophages used to treat pulmonary infections. Antimicrob. Agents Chemother. 2013, 57, 5961–5968. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, H.M.; McKean, K.A.; Wang, I.-N. Phage fitness may help predict phage therapy efficacy. Bacteriophage 2014, 4, e964081. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Gill, J.J. The habits of highly effective phages: Population dynamics as a framework for identifying therapeutic phages. Front. Microbiol. 2014, 5, 618. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Seed, K.D. Battling phages: How bacteria defend against viral attack. PLoS Pathog. 2015, 11, e1004847. [Google Scholar] [CrossRef] [PubMed]
- Young, R.; Gill, J.J. Microbiology. Phage therapy redux-what is to be done? Science 2015, 350, 1163–1164. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.; Ward, S.; Hyman, P. More is better: Selecting for broad host range bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [CrossRef] [PubMed]
- Kutter, E. Phage host range and efficiency of plating. Methods Mol. Biol. 2009, 501, 141–149. [Google Scholar] [PubMed]
- Wilson, G.S.; Artkinson, J.D. Typing of staphylococci by bacteriophage method. Lancet 1945, 245, 647–648. [Google Scholar] [CrossRef]
- Zierdt, C.H.; Marsh, H.H. The optimal single phage concentration for the typing of Staphylococcus aureus. Bacteriol. Proc. 1961, 61, 111. [Google Scholar]
- Mirzaei, M.K.; Nilsson, A.S. Correction: Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0127606. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Yin, J. Bacteriophage plaques: Theory and analysis. Methods Mol. Biol. 2009, 501, 161–174. [Google Scholar] [PubMed]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef] [PubMed]
- Ruengvisesh, S.; Loquercio, A.; Castell-Perez, E.; Taylor, T.M. Inhibition of bacterial pathogens in medium and on spinach leaf surfaces using plant-derived antimicrobials loaded in surfactant micelles. J. Food Sci. 2015, 80, M2522–M2529. [Google Scholar] [CrossRef] [PubMed]
- Brandt, A.L.; Castillo, A.; Harris, K.B.; Keeton, J.T.; Hardin, M.D.; Taylor, T.M. Inhibition of listeria monocytogenes by food antimicrobials applied singly and in combination. J. Food Sci. 2010, 75, M557–M563. [Google Scholar] [CrossRef] [PubMed]
- Miedzybrodzki, R.; Fortuna, W.; Weber-Dabrowska, B.; Gorski, A. A retrospective analysis of changes in inflammatory markers in patients treated with bacterial viruses. Clin. Exp. Med. 2009, 9, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Vipra, A.; Desai, S.N.; Junjappa, R.P.; Roy, P.; Poonacha, N.; Ravinder, P.; Sriram, B.; Padmanabhan, S. Determining the minimum inhibitory concentration of bacteriophages: Potential advantages. Adv. Microbiol. 2013, 3, 181–190. [Google Scholar] [CrossRef]
- Atterbury, R.J.; Van Bergen, M.A.; Ortiz, F.; Lovell, M.A.; Harris, J.A.; de Boer, A.; Wagenaar, J.A.; Allen, V.M.; Barrow, P.A. Bacteriophage therapy to reduce Salmonella colonization of broiler chickens. Appl. Environ. Microbiol. 2007, 73, 4543–4549. [Google Scholar] [CrossRef] [PubMed]
- Anany, H.; Lingohr, E.J.; Villegas, A.; Ackermann, H.W.; She, Y.M.; Griffiths, M.W.; Kropinski, A.M. A Shigella boydii bacteriophage which resembles Salmonella phage vii. Virol. J. 2011, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.J.; Denyer, S.P.; Maillard, J.Y. Rapid and quantitative automated measurement of bacteriophage activity against cystic fibrosis isolates of Pseudomonas aeruginosa. J. Appl. Microbiol. 2011, 110, 631–640. [Google Scholar] [CrossRef] [PubMed]
- El-Arabi, T.F.; Griffiths, M.W.; She, Y.M.; Villegas, A.; Lingohr, E.J.; Kropinski, A.M. Genome sequence and analysis of a broad-host range lytic bacteriophage that infects the Bacillus cereus group. Virol. J. 2013, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Awosika, J.; Baldwin, C.; Bishop-Lilly, K.A.; Biswas, B.; Broomall, S.; Chain, P.S.; Chertkov, O.; Chokoshvili, O.; Coyne, S.; et al. Genomic comparison of Escherichia coli o104:H4 isolates from 2009 and 2011 reveals plasmid, and prophage heterogeneity, including shiga toxin encoding phage stx2. PLoS ONE 2012, 7, e48228. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Savell, J.W.; Arnold, A.N.; Gehring, K.B.; Gill, J.J.; Taylor, T.M. Prevalence and characterization of Salmonella enterica and Salmonella bacteriophages recovered from beef cattle feedlots in south Texas. J. Food Prot. 2016, 79, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Haneklaus, A.N.; Harris, K.B.; Griffin, D.B.; Edrington, T.S.; Lucia, L.M.; Savell, J.W. Salmonella prevalence in bovine lymph nodes differs among feedyards. J. Food Prot. 2012, 75, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.F.; Wang, F.S.; Beltran, P.; Plock, S.A.; Nelson, K.; Selander, R.K. Salmonella reference collection B (SARB): Strains of 37 serovars of subspecies I. J. Gen. Microbiol. 1993, 139 Pt 6, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H. Bacteriophages; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Welkos, S.; Schreiber, M.; Baer, H. Identification of Salmonella with the O-1 bacteriophage. Appl. Microbiol. 1974, 28, 618–622. [Google Scholar] [PubMed]
- Bryan, D.; El-Shibiny, A.; Hobbs, Z.; Porter, J.; Kutter, E.M. Bacteriophage T4 infection of stationary phase E. coli: Life after log from a phage perspective. Front. Microbiol. 2016, 7, 1391. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.J. Modeling of bacteriophage therapy. In Bacteriophage Ecology: Population Growth, Evolution, and Impact of Bacterial Viruses; Abedon, S.T., Ed.; Cambridge University Press: Cambridge, UK, 2008; p. 508. [Google Scholar]
- Payne, R.J.; Jansen, V.A. Understanding bacteriophage therapy as a density-dependent kinetic process. J. Theor. Biol. 2001, 208, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; McCaskill, J.S. Replication of viruses in a growing plaque: A reaction-diffusion model. Biophys. J. 1992, 61, 1540–1549. [Google Scholar] [CrossRef]
- Hendrix, R.W.; Duda, R.L. Bacteriophage lambda papa: Not the mother of all lambda phages. Science 1992, 258, 1145–1148. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Serovars | Sources/References |
|---|---|---|
| USDA4 | Anatum | T. Edrington (USDA)/[37] |
| FC1033C3 | Anatum | Cattle Feedlot Environment/[37] |
| S2029C2 | Anatum | Cattle Feedlot Environment/[37] |
| S2028C1 | Anatum | Cattle Feedlot Environment/[37] |
| FD1001A1 | Muenchen | Cattle Feedlot Environment/[37] |
| H2006-1 | Cerro | Cattle Feedlot Environment/[38] |
| 08-022 | Dublin | S. Lawhon (Texas A&M Veterinary Medicine) |
| SGSC 2475 | Enteritidis | Salmonella Genetic Stock Centre/(University of Calgary, CA)/[39] |
| 3115 | Enteritidis | T. M. Taylor (Texas A&M University) |
| SGSC 2480 | Heidelberg | Salmonella Genetic Stock Centre (University of Calgary, CA)/[39] |
| 3117 | Heidelberg | K. Cummings (Texas A&M Veterinary Medicine) |
| USDA3 | Montevideo | T. Edrington (USDA)/[37] |
| H1042-3 | Montevideo | Cattle Feedlot Environment/[38] |
| USDA2 | Newport | T. Edrington (USDA)/[37] |
| 10-014 | Newport | S. Lawhon (Texas A&M Veterinary Medicine) |
| 330-1 | Reading | S. Lawhon (Texas A&M Veterinary Medicine) |
| USDA1 | Typhimurium | T. Edrington (USDA)/[37] |
| 3116 | Typhimurium | T. M. Taylor (Texas A&M University) |
| LT2 | Typhimurium | American Type Culture Collection/ATCC 19585 |
| USDA5 | Kentucky | T. Edrington (USDA) |
| Phages | Propagation Host | Source |
|---|---|---|
| Sasha | FC1033C3 | Cattle Feedlot Environment/[37] |
| Season5 | S2029C2 | Cattle Feedlot Environment/[37] |
| Season6 | LT2 | Cattle Feedlot Environment/[37] |
| Sergei | FC1033C3 | Cattle Feedlot Environment/[37] |
| Season12 | USDA2 | Cattle Feedlot Environment/[37] |
| Season13 | LT2 | Cattle Feedlot Environment/[37] |
| Munch | USDA3 | Cattle Feedlot Environment/[37] |
| Minion | S2028C1 | Cattle Feedlot Environment/[37] |
| Mecon | FD1001A1 | Cattle Feedlot Environment/[37] |
| Season24 | USDA4 | Cattle Feedlot Environment/[37] |
| Morel | S2028C1 | Cattle Feedlot Environment/[37] |
| Season27A | FC1033C3 | Cattle Feedlot Environment/[37] |
| Sw2 | FC1033C3 | Municipal wastewater influent, TX |
| FelixO1 | LT2 | Salmonella Genetic Stock Centre (University of Calgary, CA) |
| Melville | USDA2 | Municipal wastewater influent, TX |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Y.; Wahab, L.; Gill, J.J. Development and Validation of a Microtiter Plate-Based Assay for Determination of Bacteriophage Host Range and Virulence. Viruses 2018, 10, 189. https://doi.org/10.3390/v10040189
Xie Y, Wahab L, Gill JJ. Development and Validation of a Microtiter Plate-Based Assay for Determination of Bacteriophage Host Range and Virulence. Viruses. 2018; 10(4):189. https://doi.org/10.3390/v10040189
Chicago/Turabian StyleXie, Yicheng, Laith Wahab, and Jason J. Gill. 2018. "Development and Validation of a Microtiter Plate-Based Assay for Determination of Bacteriophage Host Range and Virulence" Viruses 10, no. 4: 189. https://doi.org/10.3390/v10040189
APA StyleXie, Y., Wahab, L., & Gill, J. J. (2018). Development and Validation of a Microtiter Plate-Based Assay for Determination of Bacteriophage Host Range and Virulence. Viruses, 10(4), 189. https://doi.org/10.3390/v10040189