Hepatitis E in High-Income Countries: What Do We Know? And What Are the Knowledge Gaps?




Abstract
:1. Introduction
2. Discovery
3. HEV, a Naked Virus?
4. Taxonomy
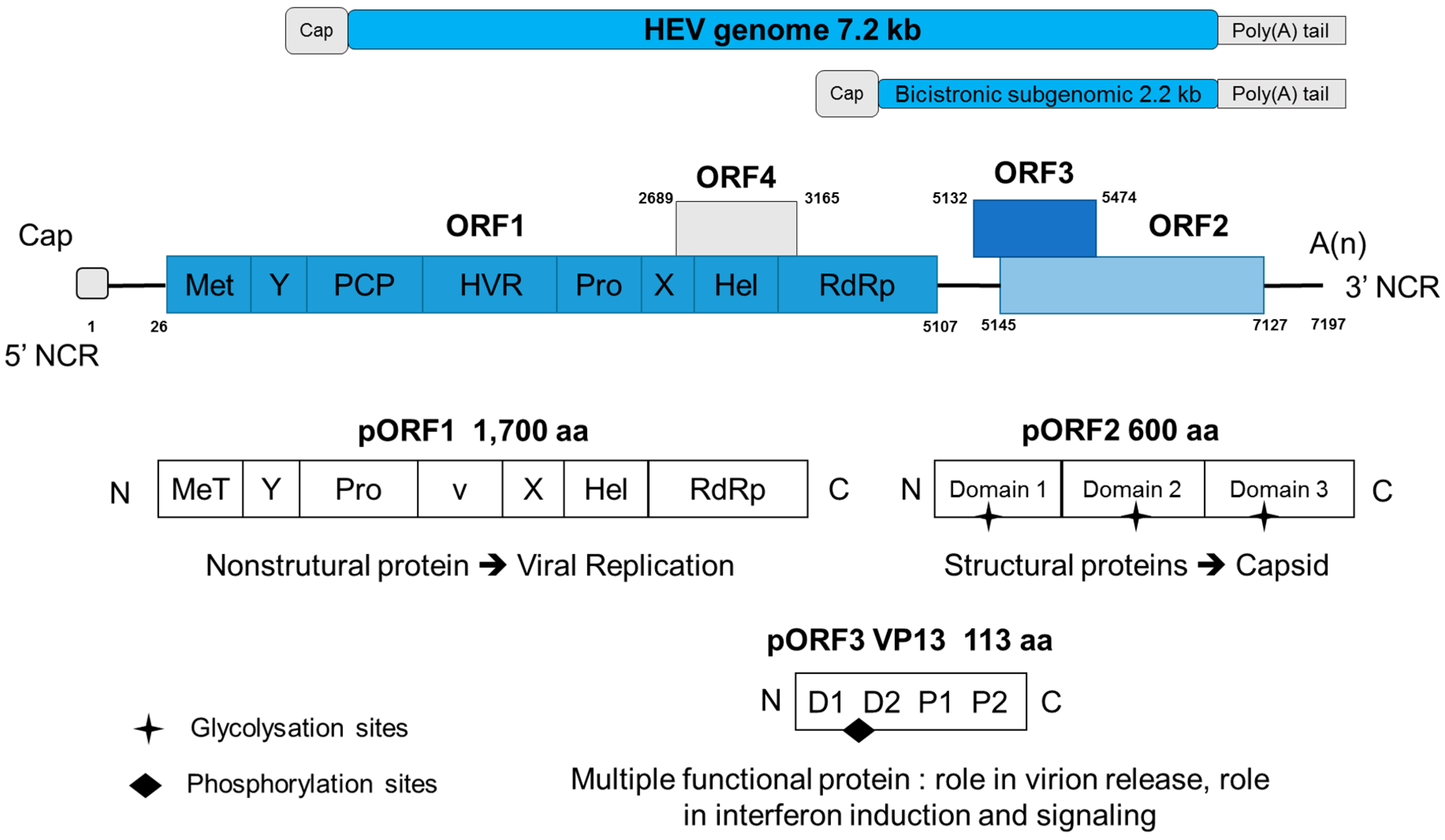
5. Genome
6. Putative Replication Cycle of HEV
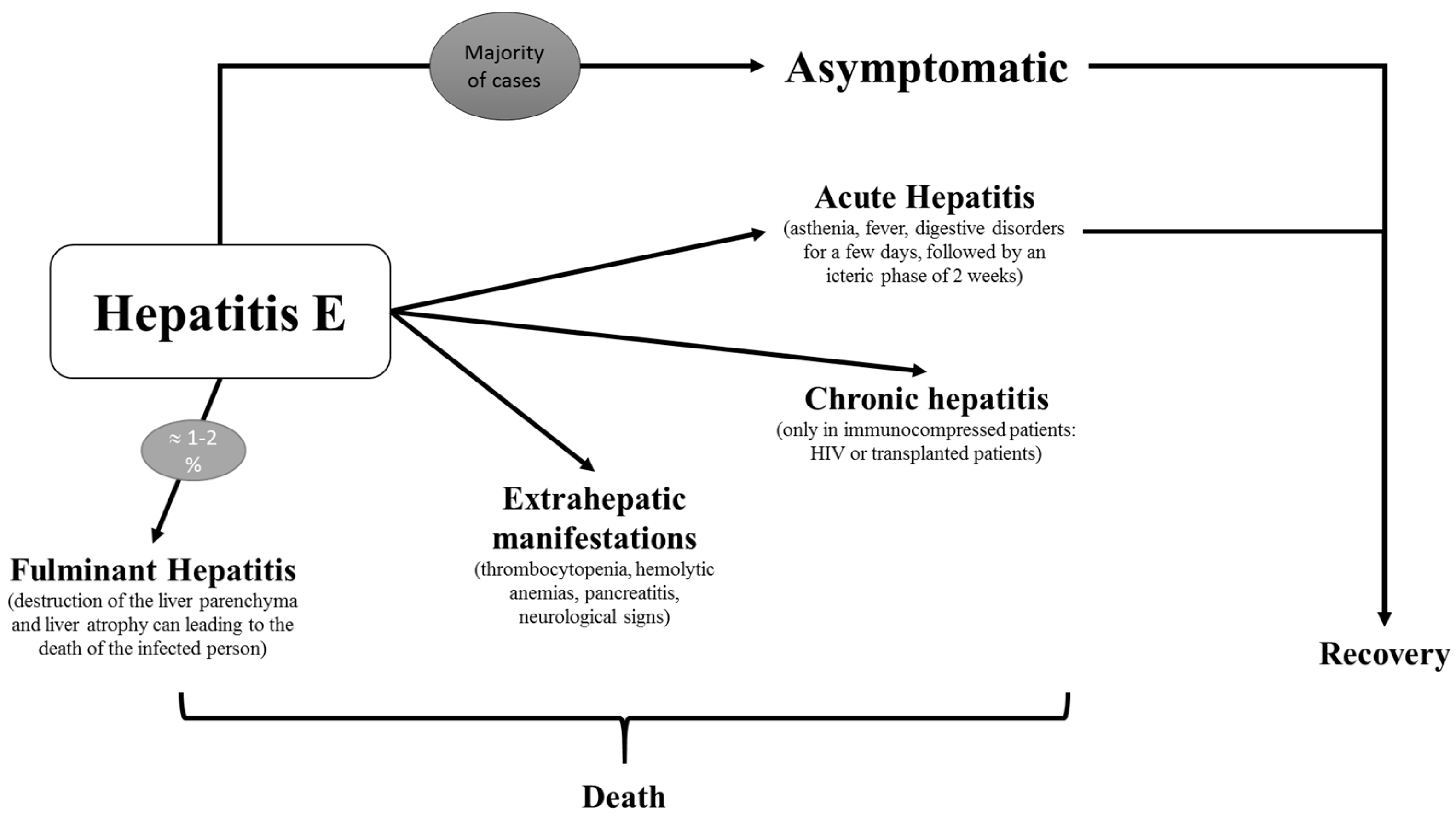
7. Clinical
7.1. Acute Hepatitis
7.2. Fulminant Hepatitis
7.3. Chronicity
7.4. Extrahepatic Manifestations
7.5. HEV Infection during Pregnancy
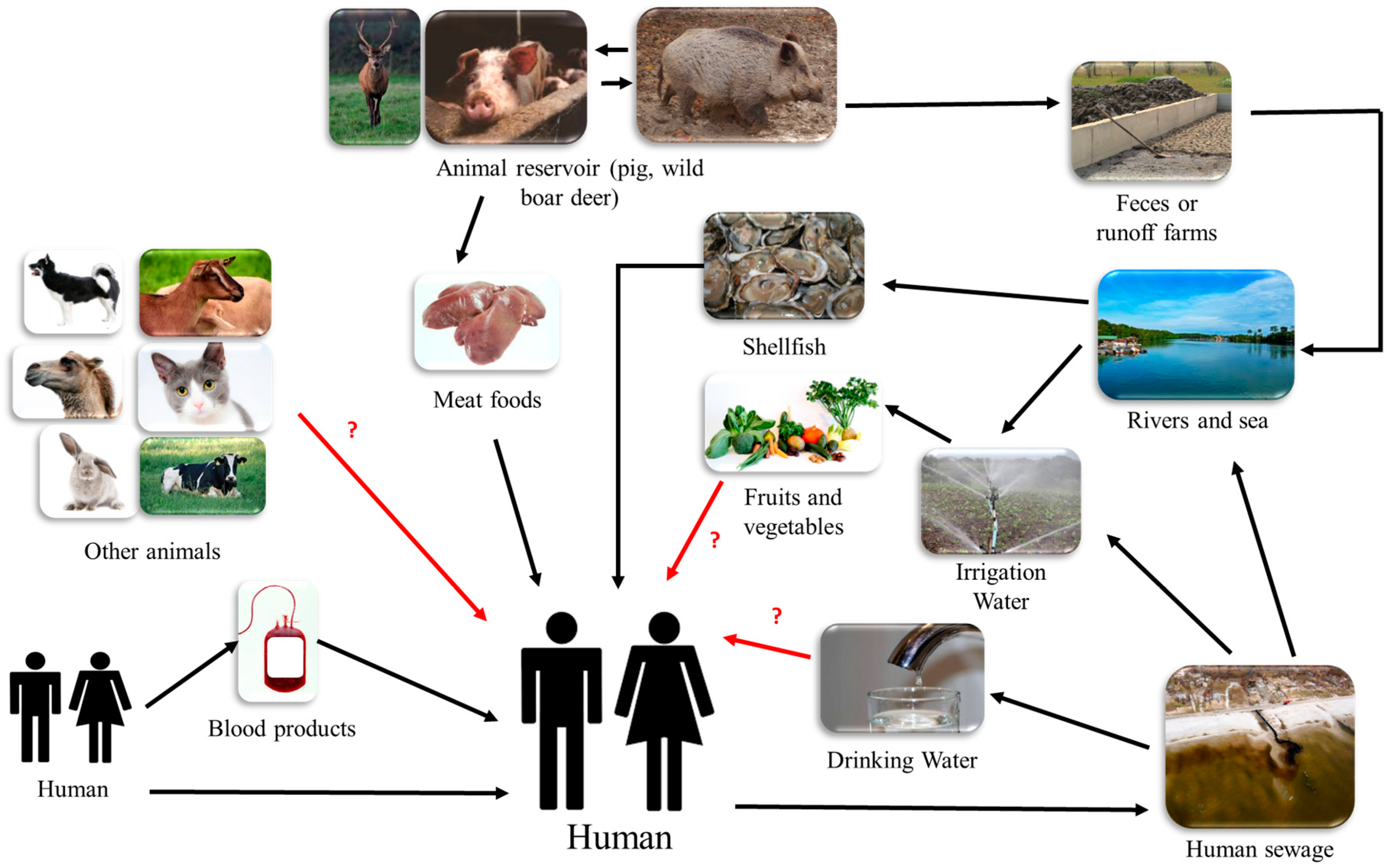
8. Different Routes of Transmission
8.1. Zoonotic Transmission
8.2. Waterborne Transmission
8.2.1. Irrigation Water
8.2.2. Contaminated Environmental Water
8.2.3. Filter Feeders: Evidence of Water Contamination and Possible Route of Foodborne Transmission
8.3. Risk of Transfusion-Related Transmission
8.4. Sexual Transmission
8.5. Other Routes of Transmission
9. Infection Markers and Diagnosis
10. Treatment, Prevention, and Vaccine
11. What Do We still Need to Know?
- Even if zoonotic transmission has been identified as an important risk factor in high-come countries, the detection of HEV has been reported in several studies performed in environmental and irrigation water, sewage, and filter feeders. These results arise questions about the role of HEV in other routes of transmission and about its denomination as an environmental virus.
- The epidemiology of HEV in high-income countries is difficult to understand because of the small number of studies, the heterogeneity between them caused by the different assays used, and the type of population included. Gaps related to different suspected animal reservoirs, including food other than pork, need to be explored in order to improve the control and prevention of HEV.
- About the clinical part, the transition to chronicity and the real numbers of symptomatic cases are still poorly understood. Studies could be conducted to deepen knowledge.
- Concerning the replication cycle of HEV, many points are still unknown, especially on the HEV receptor cell and on the exact role of the pORF3 protein. Cell culture progress and discoveries in these areas could lead to advances in treatment and vaccines. A HEV vaccine is currently available in China, but not in other countries worldwide. The development of a vaccine in pigs could reduce the spread of the virus between species and humans.
- The discovery of “quasi-enveloped” HEV particles in blood raises questions about the effects on infectivity, immunity, transmission, and the virus replication cycle.
- Regarding prevention, despite the growing number of cases, Hepatitis E remains unknown to the general public. The population could be sensitized, particularly in the hyperendemic regions. Risky behaviors may thus decrease, especially among people at risk.
Author Contributions
Conflicts of Interest
References
- Debing, Y.; Moradpour, D.; Neyts, J.; Gouttenoire, J. Update on Hepatitis E virology: Implications for clinical practice. J. Hepatol. 2016, 65, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Meng, X.J.; Renou, C. Zoonotic Hepatitis E: Animal reservoirs and emerging risks. Vet. Res. 2010, 41, 46. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Bendall, R.; Legrand-Abravanel, F.; Xia, N.S.; Ijaz, S.; Izopet, J.; Dalton, H.R. Hepatitis E. Lancet 2012, 379, 2477–2488. [Google Scholar] [CrossRef]
- Dalton, H.R.; Bendall, R.; Ijaz, S.; Banks, M. Hepatitis E: An emerging infection in developed countries. Lancet Infect. Dis. 2008, 8, 698–709. [Google Scholar] [CrossRef]
- Purcell, R.H.; Emerson, S.U. Hepatitis E: An emerging awareness of an old disease. J. Hepatol. 2008, 48, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Lapa, D.; Capobianchi, M.R.; Garbuglia, A.R. Epidemiology of Hepatitis E virus in european countries. Int. J. Mol. Sci. 2015, 16, 25711–25743. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.E.; Kmush, B.; Labrique, A.B. The epidemiology of Hepatitis E virus infections in developed countries and among immunocompromised patients. Expert Rev. Anti-Infect. Ther. 2011, 9, 1133–1148. [Google Scholar] [CrossRef] [PubMed]
- Aspinall, E.J.; Couturier, E.; Faber, M.; Said, B.; Ijaz, S.; Tavoschi, L.; Takkinen, J.; Adlhoch, C. Hepatitis E virus infection in europe: Surveillance and descriptive epidemiology of confirmed cases, 2005 to 2015. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S. Study of an epidemic of non-A, non-B hepatitis. Possibility of another human hepatitis virus distinct from post-transfusion non-A, non-B type. Am. J. Med. 1980, 68, 818–824. [Google Scholar] [CrossRef]
- Balayan, M.S.; Andjaparidze, A.G.; Savinskaya, S.S.; Ketiladze, E.S.; Braginsky, D.M.; Savinov, A.P.; Poleschuk, V.F. Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology 1983, 20, 23–31. [Google Scholar] [PubMed]
- Reyes, G.R.; Purdy, M.A.; Kim, J.P.; Luk, K.C.; Young, L.M.; Fry, K.E.; Bradley, D.W. Isolation of a cdna from the virus responsible for enterically transmitted non-A, non-B hepatitis. Science 1990, 247, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Tam, A.W.; Smith, M.M.; Guerra, M.E.; Huang, C.C.; Bradley, D.W.; Fry, K.E.; Reyes, G.R. Hepatitis E virus (HEV): Molecular cloning and sequencing of the full-length viral genome. Virology 1991, 185, 120–131. [Google Scholar] [CrossRef]
- Huang, C.C.; Nguyen, D.; Fernandez, J.; Yun, K.Y.; Fry, K.E.; Bradley, D.W.; Tam, A.W.; Reyes, G.R. Molecular cloning and sequencing of the mexico isolate of Hepatitis E virus (HEV). Virology 1992, 191, 550–558. [Google Scholar] [CrossRef]
- Khuroo, M.S. Discovery of Hepatitis E: The epidemic non-A, non-B hepatitis 30 years down the memory lane. Virus Res. 2011, 161, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Teo, C.G. Fatal outbreaks of jaundice in pregnancy and the epidemic history of Hepatitis E. Epidemiol. Infect. 2012, 140, 767–787. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.W. Enterically-transmitted non-A, non-B hepatitis. Br. Med. Bull. 1990, 46, 442–461. [Google Scholar] [CrossRef] [PubMed]
- Emerson, S.U.; Purcell, R.H. Hepatitis E virus. Rev. Med. Virol. 2003, 13, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Hoofnagle, J.H.; Nelson, K.E.; Purcell, R.H. Hepatitis E. N. Engl. J. Med. 2012, 367, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Tanaka, T.; Takahashi, H.; Hoshino, Y.; Nagashima, S.; Jirintai; Mizuo, H.; Yazaki, Y.; Takagi, T.; Azuma, M.; et al. Hepatitis E virus (HEV) strains in serum samples can replicate efficiently in cultured cells despite the coexistence of HEV antibodies: Characterization of HEV virions in blood circulation. J. Clin. Microbiol. 2010, 48, 1112–1125. [Google Scholar] [CrossRef] [PubMed]
- Chapuy-Regaud, S.; Dubois, M.; Plisson-Chastang, C.; Bonnefois, T.; Lhomme, S.; Bertrand-Michel, J.; You, B.; Simoneau, S.; Gleizes, P.E.; Flan, B.; et al. Characterization of the lipid envelope of exosome encapsulated HEV particles protected from the immune response. Biochimie 2017, 141, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Hensley, L.; McKnight, K.L.; Hu, F.; Madden, V.; Ping, L.; Jeong, S.H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Simmonds, P.; International Committee on Taxonomy of Viruses Hepeviridae Study Group; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.; Purdy, M.A. Consensus proposals for classification of the family hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Simmonds, P.; Izopet, J.; Oliveira-Filho, E.F.; Ulrich, R.G.; Johne, R.; Koenig, M.; Jameel, S.; Harrison, T.J.; Meng, X.-J.; et al. Proposed reference sequences for Hepatitis E virus subtypes. J. Gen. Virol. 2016, 97, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New Hepatitis E virus genotype in camels, the middle east. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Terada, S.; Kokuryu, H.; Arai, M.; Mishiro, S. A wild boar-derived Hepatitis E virus isolate presumably representing so far unidentified genotype 5. Kanzo 2010, 51, 536–538. [Google Scholar] [CrossRef]
- Takahashi, M.; Nishizawa, T.; Sato, H.; Sato, Y.; Jirintai; Nagashima, S.; Okamoto, H. Analysis of the full-length genome of a Hepatitis E virus isolate obtained from a wild boar in japan that is classifiable into a novel genotype. J. Gen. Virol. 2011, 92, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Nishizawa, T.; Nagashima, S.; Jirintai, S.; Kawakami, M.; Sonoda, Y.; Suzuki, T.; Yamamoto, S.; Shigemoto, K.; Ashida, K.; et al. Molecular characterization of a novel Hepatitis E virus (HEV) strain obtained from a wild boar in japan that is highly divergent from the previously recognized HEV strains. Virus Res. 2014, 180, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Cao, K.Y.; Wernery, U.; Schountz, T.; Chiu, T.H.; Tsang, A.K.; Wong, P.C.; Wong, E.Y.; et al. New Hepatitis E virus genotype in bactrian camels, xinjiang, china, 2013. Emerg. Infect. Dis. 2016, 22, 2219–2221. [Google Scholar] [CrossRef] [PubMed]
- Purdy, M.A.; Harrison, T.J.; Jameel, S.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Smith, D.B.; ICTV Report, C. ICTV virus taxonomy profile: Hepeviridae. J. Gen. Virol. 2017, 98, 2645–2646. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Thakral, D.; Rehman, S. Hepatitis E virus. Rev. Med. Virol. 2007, 17, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Gorbalenya, A.E.; Purdy, M.A.; Rozanov, M.N.; Reyes, G.R.; Bradley, D.W. Computer-assisted assignment of functional domains in the nonstructural polyprotein of Hepatitis E virus: Delineation of an additional group of positive-strand RNA plant and animal viruses. Proc. Natl. Acad. Sci. USA 1992, 89, 8259–8263. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Hyams, K.C.; Purdy, M.A.; Krawczynski, K.; Ching, W.M.; Fry, K.E.; Reyes, G.R.; Bradley, D.W.; Carl, M. Human linear B-cell epitopes encoded by the Hepatitis E virus include determinants in the RNA-dependent RNA polymerase. Proc. Natl. Acad. Sci. USA 1992, 89, 3855–3858. [Google Scholar] [CrossRef] [PubMed]
- Holla, R.P.; Ahmad, I.; Ahmad, Z.; Jameel, S. Molecular virology of Hepatitis E virus. Semin. Liver Dis. 2013, 33, 3–14. [Google Scholar] [PubMed]
- Suppiah, S.; Zhou, Y.; Frey, T.K. Lack of processing of the expressed ORF1 gene product of Hepatitis E virus. Virol. J. 2011, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, D.; Thomas, S.; Chakraborty, M.; Jameel, S. Expression and processing of the Hepatitis E virus ORF1 nonstructural polyprotein. Virol. J. 2006, 3, 38. [Google Scholar] [CrossRef] [PubMed]
- Zafrullah, M.; Ozdener, M.H.; Kumar, R.; Panda, S.K.; Jameel, S. Mutational analysis of glycosylation, membrane translocation, and cell surface expression of the Hepatitis E virus ORF2 protein. J. Virol. 1999, 73, 4074–4082. [Google Scholar] [PubMed]
- Jameel, S.; Zafrullah, M.; Ozdener, M.H.; Panda, S.K. Expression in animal cells and characterization of the Hepatitis E virus structural proteins. J. Virol. 1996, 70, 207–216. [Google Scholar] [PubMed]
- Surjit, M.; Jameel, S.; Lal, S.K. The ORF2 protein of Hepatitis E virus binds the 5’ region of viral RNA. J. Virol. 2004, 78, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Wang, J.C.; Li, T.C.; Yasutomi, Y.; Lara, J.; Khudyakov, Y.; Schofield, D.; Emerson, S.U.; Purcell, R.H.; Takeda, N.; et al. Spatial configuration of Hepatitis E virus antigenic domain. J. Virol. 2011, 85, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.; Torian, U.; Nguyen, H.; Emerson, S.U. A bicistronic subgenomic mRNA encodes both the ORF2 and ORF3 proteins of Hepatitis E virus. J. Virol. 2006, 80, 5919–5926. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Yang, C.; Gu, Y.; Song, C.; Zhang, X.; Wang, Y.; Zhang, J.; Hew, C.L.; Li, S.; Xia, N.; et al. Structural basis for the neutralization and genotype specificity of Hepatitis E virus. Proc. Natl. Acad. Sci. USA 2011, 108, 10266–10271. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Dalton, H.R.; Abravanel, F.; Izopet, J. Hepatitis E virus infection. Clin. Microbiol. Rev. 2014, 27, 116–138. [Google Scholar] [CrossRef] [PubMed]
- Graff, J.; Nguyen, H.; Yu, C.; Elkins, W.R.; St Claire, M.; Purcell, R.H.; Emerson, S.U. The open reading frame 3 gene of Hepatitis E virus contains a cis-reactive element and encodes a protein required for infection of macaques. J. Virol. 2005, 79, 6680–6689. [Google Scholar] [CrossRef] [PubMed]
- Emerson, S.U.; Nguyen, H.; Torian, U.; Purcell, R.H. ORF3 protein of Hepatitis E virus is not required for replication, virion assembly, or infection of hepatoma cells in vitro. J. Virol. 2006, 80, 10457–10464. [Google Scholar] [CrossRef] [PubMed]
- Osterman, A.; Stellberger, T.; Gebhardt, A.; Kurz, M.; Friedel, C.C.; Uetz, P.; Nitschko, H.; Baiker, A.; Vizoso-Pinto, M.G. The Hepatitis E virus intraviral interactome. Sci. Rep. 2015, 5, 13872. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zafrullah, M.; Mixson-Hayden, T.; Dai, X.; Liang, J.; Meng, J.; Kamili, S. Suppression of interferon-alpha signaling by Hepatitis E virus. Hepatology 2012, 55, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Nan, Y.; Ma, Z.; Wang, R.; Yu, Y.; Kannan, H.; Fredericksen, B.; Zhang, Y.J. Enhancement of interferon induction by ORF3 product of Hepatitis E virus. J. Virol. 2014, 88, 8696–8705. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, F.; Tian, D.; Wang, J.; Zheng, Z.; Xia, N. Open reading frame 3 of genotype 1 Hepatitis E virus inhibits nuclear factor-κB signaling induced by tumor necrosis factor-α in human A549 lung epithelial cells. PLoS ONE 2014, 9, e100787. [Google Scholar]
- Takahashi, M.; Yamada, K.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Tanaka, T.; Okamoto, H. Monoclonal antibodies raised against the ORF3 protein of Hepatitis E virus (HEV) can capture HEV particles in culture supernatant and serum but not those in feces. Arch. Virol. 2008, 153, 1703–1713. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Heller, B.; Capuccino, J.M.; Song, B.; Nimgaonkar, I.; Hrebikova, G.; Contreras, J.E.; Ploss, A. Hepatitis E virus ORF3 is a functional ion channel required for release of infectious particles. Proc. Natl. Acad. Sci. USA 2017, 114, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Nair, V.P.; Anang, S.; Subramani, C.; Madhvi, A.; Bakshi, K.; Srivastava, A.; Shalimar; Nayak, B.; Ranjith Kumar, C.T.; Surjit, M. Endoplasmic reticulum stress induced synthesis of a novel viral factor mediates efficient replication of genotype-1 Hepatitis E virus. PLoS Pathog. 2016, 12, e1005521. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Holla, R.P.; Jameel, S. Molecular virology of Hepatitis E virus. Virus Res. 2011, 161, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.P.; Kasorndorkbua, C.; Halbur, P.G.; Haqshenas, G.; Guenette, D.K.; Toth, T.E.; Meng, X.J. Evidence of extrahepatic sites of replication of the Hepatitis E virus in a swine model. J. Clin. Microbiol. 2001, 39, 3040–3046. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Cintas, P.; Garrouste, C.; Uro-Coste, E.; Cointault, O.; Rostaing, L. Hepatitis E virus-induced neurological symptoms in a kidney-transplant patient with chronic hepatitis. Am. J. Transplant. 2010, 10, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Kalia, M.; Chandra, V.; Rahman, S.A.; Sehgal, D.; Jameel, S. Heparan sulfate proteoglycans are required for cellular binding of the Hepatitis E virus ORF2 capsid protein and for viral infection. J. Virol. 2009, 83, 12714–12724. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Emerson, S. P.302 heat shock cognate protein 70 may mediate the entry of Hepatitis E virus into host cells. J. Clin. Virol. 2006, 36. [Google Scholar] [CrossRef]
- Kapur, N.; Thakral, D.; Durgapal, H.; Panda, S.K. Hepatitis E virus enters liver cells through receptor-dependent clathrin-mediated endocytosis. J. Viral Hepat. 2012, 19, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Holla, P.; Ahmad, I.; Ahmed, Z.; Jameel, S. Hepatitis E virus enters liver cells through a dynamin-2, clathrin and membrane cholesterol-dependent pathway. Traffic 2015, 16, 398–416. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Z.; Miao, J.; Zhao, M.; Tang, M.; Yeo, A.E.; Yu, H.; Zhang, J.; Xia, N.S. Role of heat-shock protein 90 in Hepatitis E virus capsid trafficking. J. Gen. Virol. 2010, 91, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Purdy, M.A.; Tam, A.W.; Huang, C.; Yarbough, P.; Reyes, G. Hepatitis E virus: A nonenveloped member of the α- like RNA virus supergroup. Semin. Virol. 1993, 4, 319–326. [Google Scholar] [CrossRef]
- Rehman, S.; Kapur, N.; Durgapal, H.; Panda, S.K. Subcellular localization of Hepatitis E virus (HEV) replicase. Virology 2008, 370, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Takahashi, M.; Hoshino, Y.; Takahashi, H.; Ichiyama, K.; Nagashima, S.; Tanaka, T.; Okamoto, H. ORF3 protein of Hepatitis E virus is essential for virion release from infected cells. J. Gen. Virol. 2009, 90, 1880–1891. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, S.; Yang, C.; Wei, M.; Song, C.; Zheng, Z.; Gu, Y.; Du, H.; Zhang, J.; Xia, N. Homology model and potential virus-capsid binding site of a putative HEV receptor grp78. J. Mol. Model. 2011, 17, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Korkaya, H.; Zafrullah, M.; Jameel, S.; Lal, S.K. The phosphorylated form of the ORF3 protein of Hepatitis E virus interacts with its non-glycosylated form of the major capsid protein, ORF2. J. Biol. Chem. 2002, 277, 22759–22767. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Takahashi, M.; Kusano, E.; Okamoto, H. Development and evaluation of an efficient cell-culture system for Hepatitis E virus. J. Gen. Virol. 2007, 88, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Jirintai, S.; Takahashi, M.; Kobayashi, T.; Tanggis; Nishizawa, T.; Kouki, T.; Yashiro, T.; Okamoto, H. Hepatitis E virus egress depends on the exosomal pathway, with secretory exosomes derived from multivesicular bodies. J. Gen. Virol. 2014, 95, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Takahashi, M.; Kobayashi, T.; Nishizawa, T.; Nishiyama, T.; Primadharsini, P.P.; Okamoto, H. Characterization of the quasi-enveloped Hepatitis E virus particles released by the cellular exosomal pathway. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Takahashi, M.; Jirintai, S.; Tanaka, T.; Nishizawa, T.; Yasuda, J.; Okamoto, H. Tumour susceptibility gene 101 and the vacuolar protein sorting pathway are required for the release of Hepatitis E virions. J. Gen. Virol. 2011, 92, 2838–2848. [Google Scholar] [CrossRef] [PubMed]
- Montpellier, C.; Wychowski, C.; Sayed, I.M.; Meunier, J.C.; Saliou, J.M.; Ankavay, M.; Bull, A.; Pillez, A.; Abravanel, F.; Helle, F.; et al. Hepatitis E virus lifecycle and identification of 3 forms of the ORF2 capsid protein. Gastroenterology 2018, 154, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R. Clinical presentation of Hepatitis E. Virus Res. 2011, 161, 15–22. [Google Scholar] [CrossRef] [PubMed]
- EcHEVarria, J.M. Light and darkness: Prevalence of Hepatitis E virus infection among the general population. Scientifica (Cairo) 2014, 2014, 481016. [Google Scholar] [CrossRef] [PubMed]
- Said, B.; Ijaz, S.; Kafatos, G.; Booth, L.; Thomas, H.L.; Walsh, A.; Ramsay, M.; Morgan, D.; Hepatitis, E.I.I.T. Hepatitis E outbreak on cruise ship. Emerg. Infect. Dis. 2009, 15, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Ali, I.A.; Ghazal, H.; Fazili, J.; Nusrat, S. Mystery of Hepatitis E virus: Recent advances in its diagnosis and management. Int. J. Hepatol. 2015, 2015, 872431. [Google Scholar] [CrossRef] [PubMed]
- Nicand, E.; Bigaillon, C.; Tesse, S. Hepatitis E: An emerging disease ? Pathol. Biol. 2009, 57, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Irshad, M. Hepatitis E virus: An update on its molecular, clinical and epidemiological characteristics. Intervirology 1999, 42, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Wedemeyer, H.; Pischke, S.; Manns, M.P. Pathogenesis and treatment of Hepatitis E virus infection. Gastroenterology 2012, 142, 1388–1397. [Google Scholar] [CrossRef] [PubMed]
- Mansuy, J.M.; Mengelle, C.; Miedouge, M.; Abravanel, F.; Izopet, J. Viral Hepatitis E. Arch. Pediatr. 2009, 16, 717–720. [Google Scholar] [CrossRef]
- Naik, S.R.; Aggarwal, R.; Salunke, P.N.; Mehrotra, N.N. A large waterborne viral Hepatitis E epidemic in kanpur, india. Bull. World Health Organ. 1992, 70, 597–604. [Google Scholar] [PubMed]
- Dalton, H.R.; Hazeldine, S.; Banks, M.; Ijaz, S.; Bendall, R. Locally acquired Hepatitis E in chronic liver disease. Lancet 2007, 369, 1260. [Google Scholar] [CrossRef]
- Xin, S.; Xiao, L. Clinical manifestations of Hepatitis E. Adv. Exp. Med. Biol. 2016, 948, 175–189. [Google Scholar] [PubMed]
- Festa, S.; Garbuglia, A.R.; Baccini, F.; Panzuto, F.; Capobianchi, M.R.; Santino, I.; Purchiaroni, F.; Orgera, G.; Delle Fave, G.; Marignani, M. Acute fulminant Hepatitis E virus genotype 3E infection: Description of the first case in Europe. Scand. J. Infect. Dis. 2014, 46, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Mateos, M.L.; Molina, A.; Patier, J.L.; Moreira, V. Sporadic Hepatitis E in spain: Study of 9 autochthonous and 3 imported cases. Med. Clin. (Barc) 2005, 125, 118–119. [Google Scholar]
- Mateos Lindemann, M.L.; Morales, J.G.; Fernandez-Barredo, S.; Dominguez, M.R.; Garcia de la Hoz, F.; Halfon, P.; Perez Gracia, M.T. Fulminant Hepatitis E in a woman taking oral contraceptive medication. Am. J. Trop. Med. Hyg. 2010, 82, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Lindemann, M.L.; Diez-Aguilar, M.; Gonzalez-Galdamez, A.; Graus-Morales, J.; Moreno-Zamora, A.; Perez-Gracia, M.T. Acute, chronic and fulminant hepatitise: Seven years of experience (2004–2011). Enferm. Infecc. Microbiol. Clin. 2013, 31, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Doudier, B.; Verrot, D.; Serratrice, C.; Poucel, C.; Auguste, R.; Colson, P. Fatal outcome of autochthonous Hepatitis E in a patient with b cell lymphoma in southeastern france. J. Clin. Microbiol. 2015, 53, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Doudier, B.; Vencatassin, H.; Aherfi, S.; Colson, P. Fatal fulminant Hepatitis E associated with autoimmune hepatitis and excessive paracetamol intake in southeastern france. J. Clin. Microbiol. 2014, 52, 1294–1297. [Google Scholar] [CrossRef] [PubMed]
- Sainokami, S.; Abe, K.; Kumagai, I.; Miyasaka, A.; Endo, R.; Takikawa, Y.; Suzuki, K.; Mizuo, H.; Sugai, Y.; Akahane, Y.; et al. Epidemiological and clinical study of sporadic acute Hepatitis E caused by indigenous strains of Hepatitis E virus in japan compared with acute hepatitis a. J. Gastroenterol. 2004, 39, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Aikawa, T.; Okamoto, H. Fulminant Hepatitis E in japan. N. Engl. J. Med. 2002, 347, 1456. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pujhari, S.K.; Chawla, Y.K.; Chakraborti, A.; Ratho, R.K. Molecular detection and sequence analysis of Hepatitis E virus in patients with viral hepatitis from north india. Diagn. Microbiol. Infect. Dis. 2011, 71, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Jeblaoui, A.; Haim-Boukobza, S.; Marchadier, E.; Mokhtari, C.; Roque-Afonso, A.M. Genotype 4 Hepatitis E virus in france: An autochthonous infection with a more severe presentation. Clin. Infect. Dis. 2013, 57, e122–e126. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Garrouste, C.; Haagsma, E.B.; Garrigue, V.; Pischke, S.; Chauvet, C.; Dumortier, J.; Cannesson, A.; Cassuto-Viguier, E.; Thervet, E.; et al. Factors associated with chronic hepatitis in patients with Hepatitis E virus infection who have received solid organ transplants. Gastroenterology 2011, 140, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Murali, A.R.; Kotwal, V.; Chawla, S. Chronic Hepatitis E: A brief review. World J. Hepatol. 2015, 7, 2194–2201. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Mallet, V.; Izopet, J. Ribavirin for chronic Hepatitis E virus infection. N. Engl. J. Med. 2014, 370, 2447–2448. [Google Scholar] [CrossRef] [PubMed]
- De Niet, A.; Zaaijer, H.L.; ten Berge, I.; Weegink, C.J.; Reesink, H.W.; Beuers, U. Chronic Hepatitis E after solid organ transplantation. Neth. J. Med. 2012, 70, 261–266. [Google Scholar] [PubMed]
- Gerolami, R.; Moal, V.; Colson, P. Chronic Hepatitis E with cirrhosis in a kidney-transplant recipient. N. Engl. J. Med. 2008, 358, 859–860. [Google Scholar] [CrossRef] [PubMed]
- Te, H.S.; Drobeniuc, J.; Kamili, S.; Dong, C.; Hart, J.; Sharapov, U.M. Hepatitis E virus infection in a liver transplant recipient in the united states: A case report. Transplant. Proc. 2013, 45, 810–813. [Google Scholar] [CrossRef] [PubMed]
- Renou, C.; Lafeuillade, A.; Cadranel, J.F.; Pavio, N.; Pariente, A.; Allegre, T.; Poggi, C.; Penaranda, G.; Cordier, F.; Nicand, E. Hepatitis E virus in HIV-infected patients. AIDS 2010, 24, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Tavitian, S.; Peron, J.M.; Huynh, A.; Mansuy, J.M.; Ysebaert, L.; Huguet, F.; Vinel, J.P.; Attal, M.; Izopet, J.; Recher, C. Hepatitis E virus excretion can be prolonged in patients with hematological malignancies. J. Clin. Virol. 2010, 49, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Gauss, A.; Wenzel, J.J.; Flechtenmacher, C.; Navid, M.H.; Eisenbach, C.; Jilg, W.; Stremmel, W.; Schnitzler, P. Chronic Hepatitis E virus infection in a patient with leukemia and elevated transaminases: A case report. J. Med. Case Rep. 2012, 6, 334. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Zhang, H.; Huang, W.; J Harrison, T.; Geng, K.; Li, Z.; Wang, Y. Persistent Hepatitis E virus genotype 4 infection in a child with acute lymphoblastic leukemia. Hepat. Mon. 2014, 14, e15618. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Selves, J.; Mansuy, J.-M.; Ouezzani, L.; Péron, J.-M.; Guitard, J.; Cointault, O.; Esposito, L.; Abravanel, F.; Danjoux, M.; et al. Hepatitis E virus and chronic hepatitis in organ-transplant recipients. N. Engl. J. Med. 2008, 358, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Péron, J.-M.; Mansuy, J.-M.; Récher, C.; Bureau, C.; Poirson, H.; Alric, L.; Izopet, J.; Vinel, J.-P. Prolonged Hepatitis E in an immunocompromised patient. J. Gastroenterol. Hepatol. 2006, 21, 1223–1224. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Bendall, R.P.; Keane, F.E.; Tedder, R.S.; Ijaz, S. Persistent carriage of Hepatitis E virus in patients with HIV infection. N. Engl. J. Med. 2009, 361, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, P.; Steinmann, E.; Manns, M.P.; Wedemeyer, H. The impact of Hepatitis E in the liver transplant setting. J. Hepatol. 2014, 61, 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Shimizu, Y.K.; Tanaka, T.; Kuroda, K.; Arakawa, Y.; Takahashi, K.; Mishiro, S.; Shimizu, K.; Moriyama, M. Persistent infection of Hepatitis E virus transmitted by blood transfusion in a patient with t-cell lymphoma. Hepatol. Res. Off. J. Jpn. Soc. Hepatol. 2007, 37, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Jameel, S. Hepatitis E. Hepatology 2011, 54, 2218–2226. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Dhiver, C.; Gerolami, R. Hepatitis E virus as a newly identified cause of acute viral hepatitis during human immunodeficiency virus infection. Clin. Microbiol. Infect. 2008, 14, 1176–1180. [Google Scholar] [CrossRef] [PubMed]
- Abid, S.; Khan, A.H. Severe hemolysis and renal failure in glucose-6-phosphate dehydrogenase deficient patients with Hepatitis E. Am. J. Gastroenterol. 2002, 97, 1544–1547. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Marion, O.; Abravanel, F.; Izopet, J.; Dalton, H.R. Extrahepatic manifestations of Hepatitis E virus. Liver Int. 2016, 36, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Deroux, A.; Brion, J.P.; Hyerle, L.; Belbezier, A.; Vaillant, M.; Mosnier, E.; Larrat, S.; Morand, P.; Pavese, P. Association between Hepatitis E and neurological disorders: Two case studies and literature review. J. Clin. Virol. 2014, 60, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Abravanel, F.; Lhomme, S.; Rostaing, L.; Izopet, J. Hepatitis E virus: Chronic infection, extra-hepatic manifestations, and treatment. Clin. Res. Hepatol. Gastroenterol. 2015, 39, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Sharma, B.C.; Sarin, S.K. Hepatitis E virus as an etiology of acute exacerbation of previously unrecognized asymptomatic patients with hepatitis B virus-related chronic liver disease. J. Gastroenterol. Hepatol. 2008, 23, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Monga, R.; Garg, S.; Tyagi, P.; Kumar, N. Superimposed acute Hepatitis E infection in patients with chronic liver disease. Indian J. Gastroenterol. 2004, 23, 50–52. [Google Scholar] [PubMed]
- Dahiya, M.; Kumar, A.; Kar, P.; Gupta, R.K. Acute viral hepatitis in third trimester of pregnancy. Indian J. Gastroenterol. 2005, 24, 128–129. [Google Scholar] [PubMed]
- Jilani, N.; Das, B.C.; Husain, S.A.; Baweja, U.K.; Chattopadhya, D.; Gupta, R.K.; Sardana, S.; Kar, P. Hepatitis E virus infection and fulminant hepatic failure during pregnancy. J. Gastroenterol. Hepatol. 2007, 22, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Beniwal, M.; Kar, P.; Sharma, J.B.; Murthy, N.S. Hepatitis E in pregnancy. Int. J. Gynaecol. Obstet. 2004, 85, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, P.; Bhandari, D.; Sharma, D.; Bhandari, B.P. A study of viral hepatitis during pregnancy in nepal medical college teaching hospital. Nepal Med. Coll. J. 2009, 11, 192–194. [Google Scholar] [PubMed]
- Prasad, G.S.; Prasad, S.; Bhupali, A.; Patil, A.N.; Parashar, K. A study of Hepatitis E in pregnancy: Maternal and fetal outcome. J. Obstet. Gynaecol. India 2016, 66, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.P.; Jain, A.K.; Naik, G.; Soni, N.; Chitnis, D.S. Viral hepatitis during pregnancy. Int. J. Gynaecol. Obstet. 2001, 72, 103–108. [Google Scholar] [CrossRef]
- Lachish, T.; Erez, O.; Daudi, N.; Shouval, D.; Schwartz, E. Acute Hepatitis E virus in pregnant women in israel and in other industrialized countries. J. Clin. Virol. 2015, 73, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Anty, R.; Ollier, L.; Peron, J.M.; Nicand, E.; Cannavo, I.; Bongain, A.; Giordanengo, V.; Tran, A. First case report of an acute genotype 3 Hepatitis E infected pregnant woman living in south-eastern france. J. Clin. Virol. 2012, 54, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, A.; Nelson-Piercy, C. Fulminant liver failure following Hepatitis E in pregnancy. J. Obstet. Gynaecol. 2006, 26, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, J.; Wenzel, J.J.; Soboletzki, M.; Flux, C.; Navid, M.H.; Schnitzler, P. First case report of an acute Hepatitis E subgenotype 3c infection during pregnancy in germany. J. Clin. Virol. 2014, 61, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, M.L.; Gabilondo, G.; Romero, B.; de la Maza, O.M.; Perez-Gracia, M.T. Low prevalence of Hepatitis E infection among pregnant women in madrid, spain. J. Clin. Virol. 2010, 82, 1666–1668. [Google Scholar] [CrossRef] [PubMed]
- Renou, C.; Gobert, V.; Locher, C.; Moumen, A.; Timbely, O.; Savary, J.; Roque-Afonso, A.M. Prospective study of Hepatitis E virus infection among pregnant women in france. Virol. J. 2014, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Renou, C.; Roque-Afonso, A.M.; Pavio, N. Foodborne transmission of Hepatitis E virus from raw pork liver sausage, france. Emerg. Infect. Dis. 2014, 20, 1945–1947. [Google Scholar] [CrossRef] [PubMed]
- Berto, A.; Grierson, S.; Hakze-van der Honing, R.; Martelli, F.; Johne, R.; Reetz, J.; Ulrich, R.G.; Pavio, N.; Van der Poel, W.H.; Banks, M. Hepatitis E virus in pork liver sausage, france. Emerg. Infect. Dis. 2013, 19, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of Hepatitis E virus transmission to humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Pavio, N.; Merbah, T.; Thebault, A. Frequent Hepatitis E virus contamination in food containing raw pork liver, france. Emerg. Infect. Dis. 2014, 20, 1925–1927. [Google Scholar] [CrossRef] [PubMed]
- Bouwknegt, M.; Lodder-Verschoor, F.; van der Poel, W.H.; Rutjes, S.A.; de Roda Husman, A.M. Hepatitis E virus RNA in commercial porcine livers in the netherlands. J. Food Prot. 2007, 70, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Trojnar, E.; Anheyer-Behmenburg, H.; Binder, A.; Schotte, U.; Ellerbroek, L.; Klein, G.; Johne, R. Detection of Hepatitis E virus RNA in raw sausages and liver sausages from retail in germany using an optimized method. Int. J. Food Microbiol. 2015, 215, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Cossaboom, C.M.; Heffron, C.L.; Cao, D.; Yugo, D.M.; Houk-Miles, A.E.; Lindsay, D.S.; Zajac, A.M.; Bertke, A.S.; Elvinger, F.; Meng, X.J. Risk factors and sources of foodborne Hepatitis E virus infection in the united states. J. Med. Virol. 2016, 88, 1641–1645. [Google Scholar] [CrossRef] [PubMed]
- Yapa, C.M.; Furlong, C.; Rosewell, A.; Ward, K.A.; Adamson, S.; Shadbolt, C.; Kok, J.; Tracy, S.L.; Bowden, S.; Smedley, E.J.; et al. First reported outbreak of locally acquired Hepatitis E virus infection in australia. Med. J. Aust. 2016, 204, 274. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Chijiwa, K.; Sera, N.; Ishibashi, T.; Etoh, Y.; Shinohara, Y.; Kurata, Y.; Ishida, M.; Sakamoto, S.; Takeda, N.; et al. Hepatitis E virus transmission from wild boar meat. Emerg. Infect. Dis. 2005, 11, 1958–1960. [Google Scholar] [CrossRef] [PubMed]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of Hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef]
- Loisy-Hamon, F.; Leturnier, G. Autochthonous cases of Hepatitis E: Where does the virus come from? Impact of pig slurry treatment on reduction of the viral load and prevalence of the virus in food substrates. EuroReference 2015, 18, 13–18. [Google Scholar]
- Muller, A.; Collineau, L.; Stephan, R.; Muller, A.; Stark, K.D. Assessment of the risk of foodborne transmission and burden of Hepatitis E in switzerland. Int. J. Food Microbiol. 2017, 242, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Moor, D.; Liniger, M.; Baumgartner, A.; Felleisen, R. Screening of ready-to-eat meat products for Hepatitis E virus in switzerland. Food Environ. Virol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kokkinos, P.; Kozyra, I.; Lazic, S.; Soderberg, K.; Vasickova, P.; Bouwknegt, M.; Rutjes, S.; Willems, K.; Moloney, R.; de Roda Husman, A.M.; et al. Virological quality of irrigation water in leafy green vegetables and berry fruits production chains. Food Environ. Virol. 2017, 9, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Maunula, L.; Kaupke, A.; Vasickova, P.; Soderberg, K.; Kozyra, I.; Lazic, S.; van der Poel, W.H.; Bouwknegt, M.; Rutjes, S.; Willems, K.A.; et al. Tracing enteric viruses in the european berry fruit supply chain. Int. J. Food Microbiol. 2013, 167, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Brassard, J.; Gagne, M.J.; Genereux, M.; Cote, C. Detection of human food-borne and zoonotic viruses on irrigated, field-grown strawberries. Appl. Environ. Microbiol. 2012, 78, 3763–3766. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cassi, X.; Timoneda, N.; Gonzales-Gustavson, E.; Abril, J.F.; Bofill-Mas, S.; Girones, R. A metagenomic assessment of viral contamination on fresh parsley plants irrigated with fecally tainted river water. Int. J. Food Microbiol. 2017, 257, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Kokkinos, P.; Kozyra, I.; Lazic, S.; Bouwknegt, M.; Rutjes, S.; Willems, K.; Moloney, R.; de Roda Husman, A.M.; Kaupke, A.; Legaki, E.; et al. Harmonised investigation of the occurrence of human enteric viruses in the leafy green vegetable supply chain in three european countries. Food Environ. Virol. 2012, 4, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, A.; Ertem, M.; Ilcin, E.; Ozekinci, T. A special risk group for Hepatitis E infection: Turkish agricultural workers who use untreated waste water for irrigation. Epidemiol. Infect. 2003, 131, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Iaconelli, M.; Purpari, G.; Della Libera, S.; Petricca, S.; Guercio, A.; Ciccaglione, A.R.; Bruni, R.; Taffon, S.; Equestre, M.; Fratini, M.; et al. Hepatitis A and E viruses in wastewaters, in river waters, and in bivalve molluscs in italy. Food Environ. Virol. 2015, 7, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Steyer, A.; Naglic, T.; Mocilnik, T.; Poljsak-Prijatelj, M.; Poljak, M. Hepatitis E virus in domestic pigs and surface waters in slovenia: Prevalence and molecular characterization of a novel genotype 3 lineage. Infect. Genet. Evol. 2011, 11, 1732–1737. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Miyamura, T.; Takeda, N. Detection of Hepatitis E virus RNA from the bivalve yamato-shijimi (corbicula japonica) in japan. Am. J. Trop. Med. Hyg. 2007, 76, 170–172. [Google Scholar] [PubMed]
- Rutjes, S.A.; Lodder, W.J.; Lodder-Verschoor, F.; van den Berg, H.H.; Vennema, H.; Duizer, E.; Koopmans, M.; de Roda Husman, A.M. Sources of Hepatitis E virus genotype 3 in the netherlands. Emerg. Infect. Dis. 2009, 15, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Borgen, K.; Herremans, T.; Duizer, E.; Vennema, H.; Rutjes, S.; Bosman, A.; de Roda Husman, A.M.; Koopmans, M. Non-travel related Hepatitis E virus genotype 3 infections in the netherlands; a case series 2004–2006. BMC Infect. Dis. 2008, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, J.R.; Oliveira, D.; Rivadulla, E.; Abreu-Silva, J.; Varela, M.F.; Romalde, J.L.; Nascimento, M.S. Hepatitis E virus genotype 3 in mussels (mytilus galloprovinciallis), spain. Food Microbiol. 2016, 58, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Crossan, C.; Baker, P.J.; Craft, J.; Takeuchi, Y.; Dalton, H.R.; Scobie, L. Hepatitis E virus genotype 3 in shellfish, united kingdom. Emerg. Infect. Dis. 2012, 18, 2085–2087. [Google Scholar] [CrossRef] [PubMed]
- Mansuy, J.M.; Gallian, P.; Dimeglio, C.; Saune, K.; Arnaud, C.; Pelletier, B.; Morel, P.; Legrand, D.; Tiberghien, P.; Izopet, J. A nationwide survey of Hepatitis E viral infection in french blood donors. Hepatology 2016, 63, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Hogema, B.M.; Molier, M.; Sjerps, M.; de Waal, M.; van Swieten, P.; van de Laar, T.; Molenaar-de Backer, M.; Zaaijer, H.L. Incidence and duration of Hepatitis E virus infection in dutch blood donors. Transfusion 2016, 56, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Hoad, V.C.; Seed, C.R.; Fryk, J.J.; Harley, R.; Flower, R.L.P.; Hogema, B.M.; Kiely, P.; Faddy, H.M. Hepatitis E virus RNA in australian blood donors: Prevalence and risk assessment. Vox Sang. 2017, 112, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.C.; Flower, R.L.; Seed, C.R.; Keller, A.J.; Harley, R.; Chan, H.T.; Hoad, V.; Warrilow, D.; Northill, J.; Holmberg, J.A.; et al. Hepatitis E virus RNA in australian blood donations. Transfusion 2016, 56, 3086–3093. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Hofmann, M.; Danzer, M.; Hofer, K.; Kaar, J.; Gabriel, C. Seroprevalence and incidence of Hepatitis E in blood donors in upper austria. PLoS ONE 2015, 10, e0119576. [Google Scholar] [CrossRef] [PubMed]
- Harritshoj, L.H.; Holm, D.K.; Saekmose, S.G.; Jensen, B.A.; Hogema, B.M.; Fischer, T.K.; Midgley, S.E.; Krog, J.S.; Erikstrup, C.; Ullum, H. Low transfusion transmission of Hepatitis E among 25,637 single-donation, nucleic acid-tested blood donors. Transfusion 2016, 56, 2225–2232. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, S.; Szypulska, R.; Tettmar, K.I.; Kitchen, A.; Tedder, R.S. Detection of Hepatitis E virus RNA in plasma mini-pools from blood donors in england. Vox Sang. 2012, 102, 272. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, P.E.; Ijaz, S.; Brailsford, S.R.; Brett, R.; Dicks, S.; Haywood, B.; Kennedy, I.T.R.; Kitchen, A.; Patel, P.; Poh, J.; et al. Hepatitis E virus in blood components: A prevalence and transmission study in southeast england. Lancet 2014, 384, 1766–1773. [Google Scholar] [CrossRef]
- Gallian, P.; Lhomme, S.; Piquet, Y.; Saune, K.; Abravanel, F.; Assal, A.; Tiberghien, P.; Izopet, J. Hepatitis E virus infections in blood donors, france. Emerg. Infect. Dis. 2014, 20, 1914–1917. [Google Scholar] [CrossRef] [PubMed]
- Gallian, P.; Couchouron, A.; Dupont, I.; Fabra, C.; Piquet, Y.; Djoudi, R.; Assal, A.; Tiberghien, P. Comparison of Hepatitis E virus nucleic acid test screening platforms and RNA prevalence in french blood donors. Transfusion 2017, 57, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, T.; Diekmann, J.; Johne, R.; Eberhardt, M.; Knabbe, C.; Dreier, J. Novel approach for detection of Hepatitis E virus infection in german blood donors. J. Clin. Microbiol. 2012, 50, 2708–2713. [Google Scholar] [CrossRef] [PubMed]
- Baylis, S.A.; Gartner, T.; Nick, S.; Ovemyr, J.; Blumel, J. Occurrence of Hepatitis E virus RNA in plasma donations from sweden, germany and the united states. Vox Sang. 2012, 103, 89–90. [Google Scholar] [CrossRef] [PubMed]
- O’Riordan, J.; Boland, F.; Williams, P.; Donnellan, J.; Hogema, B.M.; Ijaz, S.; Murphy, W.G. Hepatitis E virus infection in the irish blood donor population. Transfusion 2016, 56, 2868–2876. [Google Scholar] [CrossRef] [PubMed]
- Minagi, T.; Okamoto, H.; Ikegawa, M.; Ideno, S.; Takahashi, K.; Sakai, K.; Hagiwara, K.; Yunoki, M.; Wakisaka, A. Hepatitis E virus in donor plasma collected in japan. Vox Sang. 2016, 111, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Slot, E.; Hogema, B.M.; Riezebos-Brilman, A.; Kok, T.M.; Molier, M.; Zaaijer, H.L. Silent Hepatitis E virus infection in dutch blood donors, 2011 to 2012. Eurosurveillance 2013, 18, 20550. [Google Scholar] [CrossRef] [PubMed]
- Grabarczyk, P.; Sulkowska, E.; Gdowska, J.; Kopacz, A.; Liszewski, G.; Kubicka-Russel, D.; Baylis, S.A.; Corman, V.M.; Nocen, E.; Piotrowski, D.; et al. Molecular and serological infection marker screening in blood donors indicates high endemicity of Hepatitis E virus in poland. Transfusion 2018, 58, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Cleland, A.; Smith, L.; Crossan, C.; Blatchford, O.; Dalton, H.R.; Scobie, L.; Petrik, J. Hepatitis E virus in scottish blood donors. Vox Sang. 2013, 105, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Sauleda, S.; Ong, E.; Bes, M.; Janssen, A.; Cory, R.; Babizki, M.; Shin, T.; Lindquist, A.; Hoang, A.; Vang, L.; et al. Seroprevalence of Hepatitis E virus (HEV) and detection of HEV RNA with a transcription-mediated amplification assay in blood donors from catalonia (spain). Transfusion 2015, 55, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Stramer, S.L.; Moritz, E.D.; Foster, G.A.; Ong, E.; Linnen, J.M.; Hogema, B.M.; Mak, M.; Chia, C.P.; Dodd, R.Y. Hepatitis E virus: Seroprevalence and frequency of viral RNA detection among us blood donors. Transfusion 2016, 56, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S.; Kamili, S.; Yattoo, G.N. Hepatitis E virus infection may be transmitted through blood transfusions in an endemic area. J. Gastroenterol. Hepatol. 2004, 19, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, K.; Kang, J.H.; Sakata, H.; Takahashi, K.; Shindo, M.; Kato, M.; Sato, S.; Kato, T.; Nishimori, H.; Tsuji, K.; et al. A case of transfusion-transmitted Hepatitis E caused by blood from a donor infected with Hepatitis E virus via zoonotic food-borne route. Transfusion 2008, 48, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Arankalle, V.A.; Chobe, L.P. Retrospective analysis of blood transfusion recipients: Evidence for post-transfusion Hepatitis E. Vox Sang. 2000, 79, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, T.; Tsukamoto, Y.; Yamazaki, C.; Masuko, K.; Tsuda, F.; Takahashi, M.; Nishizawa, T.; Okamoto, H. Prevalence of Hepatitis E virus infection among hemodialysis patients in japan: Evidence for infection with a genotype 3 HEV by blood transfusion. J. Med. Virol. 2004, 74, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, M.; Gupta, E. Transfusion-transmitted Hepatitis E: Is screening warranted? Indian J. Med. Microbiol. 2011, 29, 353–358. [Google Scholar] [PubMed]
- Hoad, V.C.; Gibbs, T.; Ravikumara, M.; Nash, M.; Levy, A.; Tracy, S.L.; Mews, C.; Perkowska-Guse, Z.; Faddy, H.M.; Bowden, S. First confirmed case of transfusion-transmitted Hepatitis E in australia. Med. J. Aust. 2017, 206, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Izopet, J.; Lhomme, S.; Chapuy-Regaud, S.; Mansuy, J.M.; Kamar, N.; Abravanel, F. HEV and transfusion-recipient risk. Trans. Clin. Biol. 2017, 24, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Balayan, M.S.; Fedorova, O.E.; Mikhailov, M.I.; Rytick, P.G.; Eremin, V.F.; Danilova, T.I.; SHEVelev, B.I.; GorbacHEVa, E.C.; Pankova, G.Y. Antibody to Hepatitis E virus in HIV-infected individuals and AIDS patients. J. Viral Hepat. 1997, 4, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.B.; Engle, R.E.; Jacobsen, S.E.; Krarup, H.B.; Georgsen, J.; Purcell, R.H. High prevalence of Hepatitis E antibodies among danish prisoners and drug users. J. Med. Virol. 2002, 66, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Keane, F.; Gompels, M.; Bendall, R.; Drayton, R.; Jennings, L.; Black, J.; Baragwanath, G.; Lin, N.; Henley, W.; Ngui, S.L.; et al. Hepatitis E virus coinfection in patients with HIV infection. HIV Med. 2012, 13, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Payne, B.A.; Medhi, M.; Ijaz, S.; Valappil, M.; Savage, E.J.; Gill, O.N.; Tedder, R.; Schwab, U. Hepatitis E virus seroprevalence among men who have sex with men, united kingdom. Emerg. Infect. Dis. 2013, 19, 333–335. [Google Scholar] [CrossRef] [PubMed]
- Politou, M.; Boti, S.; Androutsakos, T.; Valsami, S.; Pittaras, T.; Kapsimali, V. Seroprevalence of Hepatitis E in HIV infected patients in greece. J. Med. Virol. 2015, 87, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Montella, F.; Rezza, G.; Di Sora, F.; Pezzotti, P.; Recchia, O. Association between Hepatitis E virus and HIV infection in homosexual men. Lancet 1994, 344, 1433. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, A.; Kar, P.; Agarwal, S.; Ramji, S.; Husain, S.A.; Prasad, S.; Sharma, S. Risk factors for vertical transmission of Hepatitis E virus infection. J. Viral Hepat. 2017, 24, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S.; Kamili, S.; Khuroo, M.S. Clinical course and duration of viremia in vertically transmitted Hepatitis E virus (HEV) infection in babies born to HEV-infected mothers. J. Viral Hepat. 2009, 16, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Juarez, A.; Frias, M.; Rodriguez-Cano, D.; Cuenca-Lopez, F.; Rivero, A. Isolation of Hepatitis E virus from breast milk during acute infection. Clin. Infect. Dis. 2016, 62, 1464. [Google Scholar] [CrossRef] [PubMed]
- Mansuy, J.M.; Huynh, A.; Abravanel, F.; Recher, C.; Peron, J.M.; Izopet, J. Molecular evidence of patient-to-patient transmission of Hepatitis E virus in a hematology ward. Clin. Infect. Dis. 2009, 48, 373–374. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhang, X.; Jiang, H.; Yan, Q.; Ai, X.; Wang, Y.; Cai, J.; Jiang, L.; Wu, T.; Wang, Z.; et al. Profile of acute infectious markers in sporadic Hepatitis E. PLoS ONE 2010, 5, e13560. [Google Scholar] [CrossRef] [PubMed]
- Khuroo, M.S.; Kamili, S.; Dar, M.Y.; Moecklii, R.; Jameel, S. Hepatitis E and long-term antibody status. Lancet 1993, 341, 1355. [Google Scholar]
- Bendall, R.; Ellis, V.; Ijaz, S.; Ali, R.; Dalton, H. A comparison of two commercially available anti-HEV igg kits and a re-evaluation of anti-HEV IgG seroprevalence data in developed countries. J. Med. Virol. 2010, 82, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, T.; Diekmann, J.; Eberhardt, M.; Knabbe, C.; Dreier, J. Monitoring of anti-Hepatitis E virus antibody seroconversion in asymptomatically infected blood donors: Systematic comparison of nine commercial anti-HEV IgM and IgG assays. Viruses 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Avellon, A.; Morago, L.; Garcia-Galera del Carmen, M.; Munoz, M.; EcHEVarria, J.M. Comparative sensitivity of commercial tests for Hepatitis E genotype 3 virus antibody detection. J. Med. Virol. 2015, 87, 1934–1939. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.J.; Preiss, J.; Schemmerer, M.; Huber, B.; Jilg, W. Test performance characteristics of anti-HEV igg assays strongly influence Hepatitis E seroprevalence estimates. J. Infect. Dis. 2013, 207, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Tremeaux, P.; Lhomme, S.; Chapuy-Regaud, S.; Peron, J.M.; Alric, L.; Kamar, N.; Izopet, J.; Abravanel, F. Performance of an antigen assay for diagnosing acute Hepatitis E virus genotype 3 infection. J. Clin. Virol. 2016, 79, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Kini, D.; Sofat, S.; Naik, S.R.; Krawczynski, K. Duration of viraemia and faecal viral excretion in acute Hepatitis E. Lancet 2000, 356, 1081–1082. [Google Scholar] [CrossRef]
- Takahashi, M.; Tanaka, T.; Azuma, M.; Kusano, E.; Aikawa, T.; Shibayama, T.; Yazaki, Y.; Mizuo, H.; Inoue, J.; Okamoto, H. Prolonged fecal shedding of Hepatitis E virus (HEV) during sporadic acute Hepatitis E: Evaluation of infectivity of HEV in fecal specimens in a cell culture system. J. Clin. Microbiol. 2007, 45, 3671–3679. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Primers 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Izopet, J.; Tripon, S.; Bismuth, M.; Hillaire, S.; Dumortier, J.; Radenne, S.; Coilly, A.; Garrigue, V.; D’Alteroche, L.; et al. Ribavirin for chronic Hepatitis E virus infection in transplant recipients. N. Engl. J. Med. 2014, 370, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Rostaing, L.; Abravanel, F.; Garrouste, C.; Esposito, L.; Cardeau-Desangles, I.; Mansuy, J.M.; Selves, J.; Peron, J.M.; Otal, P.; et al. Pegylated interferon-alpha for treating chronic Hepatitis E virus infection after liver transplantation. Clin. Infect. Dis. 2010, 50, e30–e33. [Google Scholar] [CrossRef] [PubMed]
- Barnaud, E.; Rogée, S.; Garry, P.; Rose, N.; Pavio, N. Thermal inactivation of infectious Hepatitis E virus in experimentally contaminated food. Appl. Environ. Microbiol. 2012, 78, 5153–5159. [Google Scholar] [CrossRef] [PubMed]
- Emerson, S.U.; Arankalle, V.A.; Purcell, R.H. Thermal stability of Hepatitis E virus. J. Infect. Dis. 2005, 192, 930–933. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, M.P.; Scott, R.M.; Joshi, D.M.; Mammen, M.P., Jr.; Thapa, G.B.; Thapa, N.; Myint, K.S.; Fourneau, M.; Kuschner, R.A.; Shrestha, S.K.; et al. Safety and efficacy of a recombinant Hepatitis E vaccine. N. Engl. J. Med. 2007, 356, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shih, J.W.; Wu, T.; Li, S.W.; Xia, N.S. Development of the Hepatitis E vaccine: From bench to field. Semin. Liver Dis. 2013, 33, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.F.; Huang, S.J.; Wu, T.; Hu, Y.M.; Wang, Z.Z.; Wang, H.; Jiang, H.M.; Wang, Y.J.; Yan, Q.; et al. Long-term efficacy of a Hepatitis E vaccine. N. Engl. J. Med. 2015, 372, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Backer, J.A.; Berto, A.; McCreary, C.; Martelli, F.; van der Poel, W.H. Transmission dynamics of Hepatitis E virus in pigs: Estimation from field data and effect of vaccination. Epidemics 2012, 4, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.D. Current status of flavivirus vaccines. Ann. N. Y. Acad. Sci. 2001, 951, 262–271. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Rate of Positive Blood Products | References |
|---|---|---|
| Australia | 1/14,799; 1/74,131 | [155] |
| Austria | 1/8416 | [156] |
| Denmark | 1/2231 | [157] |
| England | 1/7040; 1/2848 | [158,159] |
| France | 1/2218; 1/744 | [160,161] |
| Germany | 1/1240; 1/4525 | [162,163] |
| Ireland | 1/4997 | [164] |
| Japan | 1/8173 | [165] |
| Netherlands | 1/2700; 1/726 | [166] |
| Poland | 1/1266 | [167] |
| Scotland | 1/14,520 | [168] |
| Spain | 1/3333 | [169] |
| Sweden | 1/7986 | [111,163] |
| USA | 1/9500; 0/51,075; 1/42,674 | [170] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capai, L.; Charrel, R.; Falchi, A. Hepatitis E in High-Income Countries: What Do We Know? And What Are the Knowledge Gaps? Viruses 2018, 10, 285. https://doi.org/10.3390/v10060285
Capai L, Charrel R, Falchi A. Hepatitis E in High-Income Countries: What Do We Know? And What Are the Knowledge Gaps? Viruses. 2018; 10(6):285. https://doi.org/10.3390/v10060285
Chicago/Turabian StyleCapai, Lisandru, Rémi Charrel, and Alessandra Falchi. 2018. "Hepatitis E in High-Income Countries: What Do We Know? And What Are the Knowledge Gaps?" Viruses 10, no. 6: 285. https://doi.org/10.3390/v10060285
APA StyleCapai, L., Charrel, R., & Falchi, A. (2018). Hepatitis E in High-Income Countries: What Do We Know? And What Are the Knowledge Gaps? Viruses, 10(6), 285. https://doi.org/10.3390/v10060285