miRNAs in Insects Infected by Animal and Plant Viruses



Abstract
:1. Introduction
2. miRNA Profile Changes in Insects during Virus Infection
- Some miRNAs are commonly differentially abundant due to various arbovirus infections. Bantam/bantam-3p, for example, is differentially-regulated in whole Ae. aegypti infected by DENV [64], whole Ae. albopictus-infected by DENV [65], Ae. aegypti infected by CHIKV, and whole Ae. albopictus-infected by CHIKV [66]. miR-263a and miR-34 are also differentially abundant in 4 different insect-virus pairs [65,66,67]. This suggests that there may be core miRNAs which are regulated during viral infection of insects, regardless of the virus, sample source (whole insect, body part, or cells), and time of harvest. It would be interesting to systematically explore the regulation and function of this set of core miRNAs as they pertain to host-virus interaction.
- The number of differentially regulated miRNAs changes throughout the course of infection. In a study looking at the effect of ZIKV infection on Aedes at 2, 7, and 14 days post-infection, it was noted that 10 miRNAs are differentially abundant at day 2 post-infection. At 7 dpi, this number is reduced to 7 miRNAs. At the last time-point (14 dpi), there were only 6 differentially abundant miRNAs [67]. In a study looking at the effect of DENV infection on Aedes at 2, 4, and 9 dpi, 5 miRNAs were differentially abundant at 2 dpi. This increased to 27 at 9 dpi [64]. It is interesting to note that the lowest number of differentially regulated miRNAs in ZIKV-infected mosquitoes was observed at 14 dpi when ZIKV load was highest, while the highest number of differentially regulated miRNAs were found in 2 dpi samples when ZIKV titres were still very low [67]. Additionally, the highest number of differentially regulated miRNAs in DENV-infected mosquitoes were found at 9 dpi when 50% of the samples have DENV levels that are below plaque titration-detectable amounts. Taken together these studies show that the miRNA profile is very dynamic and highlights that a single time point may not be sufficient to fully understand the impact of virus infection on the miRNA profile. This is consistent with the dynamic gene regulation observed during a virus infection.
- The direction of regulation of a miRNA can change depending on the time post infection. Some miRNAs are regulated only at specific time-points, while others are regulated at different time points. Of those which are regulated at different time points, some can take a singular direction, such as miR-2945-5p which is down-regulated in both 4 dpi and 9 dpi in the study looking at the effect of DENV infection on Ae. aegypti at 2, 4, and 9 dpi [64]. Other miRNAs can have opposite directions of regulation, depending on time of sampling. For example, in the study looking at the effect of ZIKV infection on Aedes at 2, 7, and 14 days post-infection, miR-308-5p was up-regulated at 2 dpi, but down-regulated at 5 dpi [67]. The differential regulation of individual miRNAs along the course of infection could be a reflection of the attack and counter-attack mechanisms occurring between the insect and virus during the progression of infection. Thus, for a complete picture of the interaction, miRNA dynamics over time are important.
- The general direction of miRNA regulation varies depending on tissue sample source. In whole mosquito samples infected with ZIKV [67] and DENV [64,65], the majority of miRNAs (75% or more) are down-regulated, while only 25% or less are up-regulated. However, in midgut samples, where the viral replication is most active, around 90% of the analysed miRNAs are up-regulated during DENV infection [68]. Similarly, in saliva samples, which have been shown to contain factors that enhance viral replication in the vertebrate host [69,70,71], around 80% of the miRNAs are up-regulated during CHIKV-infection [66]. Whereas the whole-body samples provide the global net change in miRNA levels, the tissue-specific samples provide the local miRNA profiles. Considering that the level of viral activity and viral titres change from initial ingestion in the foregut to infection of the midgut and then dissemination to secondary tissues including the salivary glands and finally transmission through saliva [72], it would be interesting to examine the regulation of miRNAs in different tissues and correlate the results with the location of viral activity and viral titres. In addition, because only a few cells are actually infected and become sites of viral replication (reviewed in Reference [72]), it would be interesting to examine the regulation of miRNAs in the infected cells and compare with the regulation of miRNAs in the uninfected cells in the same tissue and in the whole organism. miRNA profiling at the cellular level would provide a higher resolution view of miRNA regulation during infection and insights into the heterogeneity of miRNA profile regulation in infected versus uninfected cells.
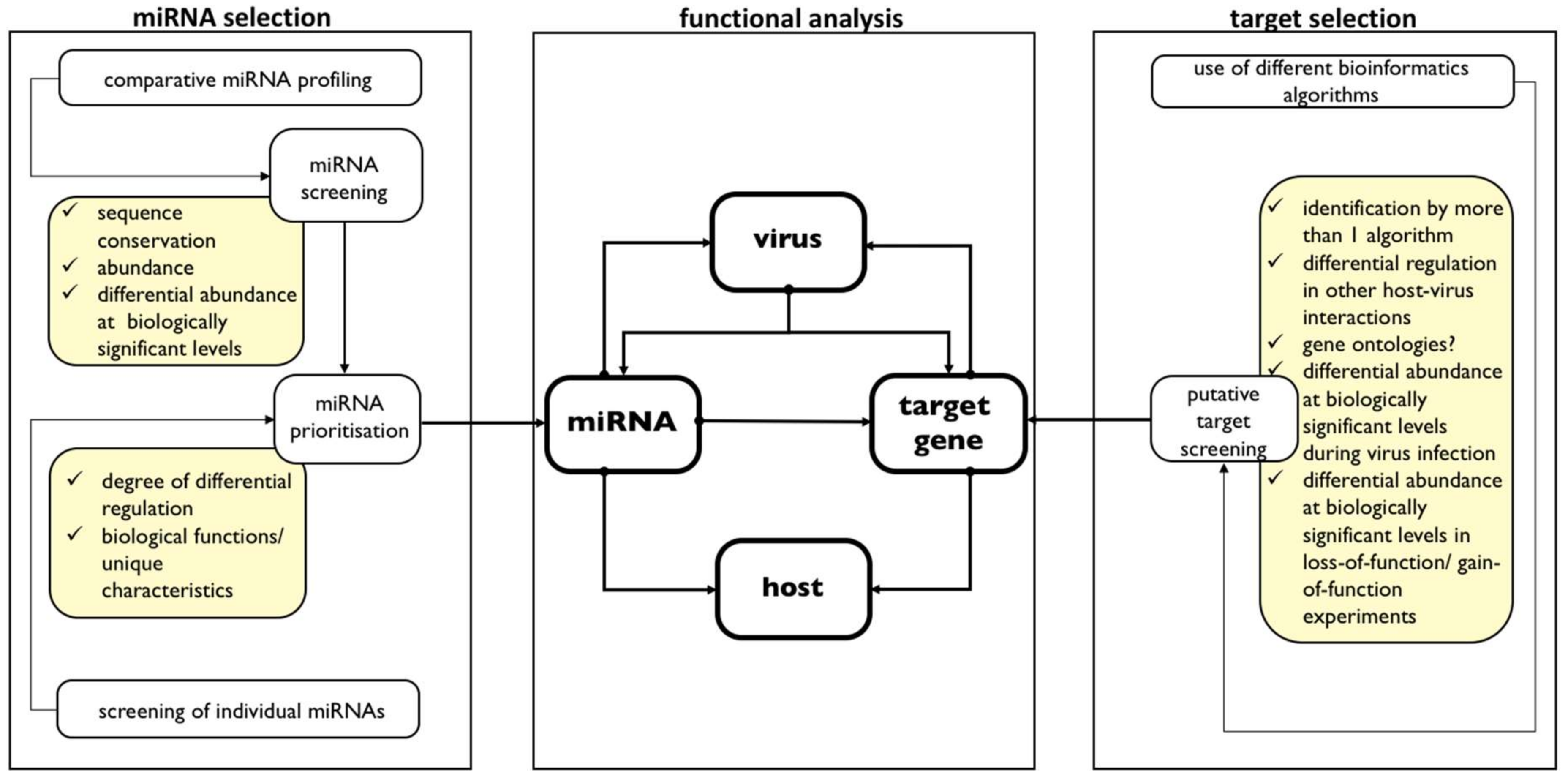
3. Functional Role of miRNAs in Insect-Virus Interaction
- Virus infection changes miRNA abundance;
- Virus infection changes target gene levels;
- Changes in miRNA affects target levels; changes in target levels during virus infection is because of changes in miRNA abundance levels;
- Changes in miRNA levels affect pathogenicity determinants in virus (example: replication) and/or in host (example: delay in mortality); because
- Changes in target levels affect pathogenicity determinants in virus (example: replication) and/or in host (example: delay in mortality).
4. Use of Drosophila for Insect-Virus Interaction Studies
5. Conclusions
Funding
Conflicts of Interest
References
- Conway, M.J.; Colpitts, T.M.; Fikrig, E. Role of the vector in arbovirus transmission. Annu. Rev. Virol. 2014, 1, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Lequime, S.; Paul, R.E.; Lambrechts, L. Determinants of arbovirus vertical transmission in mosquitoes. PLoS Pathog. 2016, 12, e1005548. [Google Scholar] [CrossRef] [PubMed]
- Beckham, J.D.; Tyler, K.L. Arbovirus infections. Contin. Lifelong Learn. Neurol. 2015, 21, 1599–1611. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Gao, X.; Gould, E.A. Factors responsible for the emergence of arboviruses; strategies, challenges and limitations for their control. Emerg. Microbes Infect. 2015, 4, e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, E.A.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carabali, M.; Hernandez, L.M.; Arauz, M.J.; Villar, L.A.; Ridde, V. Why are people with dengue dying? A scoping review of determinants for dengue mortality. BMC Infect. Dis. 2015, 15, 301. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479–480, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Ammar, E.-D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef] [PubMed]
- Bragard, C.; Caciagli, P.; Lemaire, O.; Lopez-Moya, J.J.; MacFarlane, S.; Peters, D.; Susi, P.; Torrance, L. Status and prospects of plant virus control through interference with vector transmission. Annu. Rev. Phytopathol. 2013, 51, 177–201. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.M.; Banerjee, N. Mechanisms of arthropod transmission of plant and animal viruses. Microbiol. Mol. Biol. Rev. 1999, 63, 128–148. [Google Scholar] [PubMed]
- Andret-Link, P.; Fuchs, M. Transmission specificity of plant viruses by vectors. J. Plant Pathol. 2005, 87, 153–165. [Google Scholar]
- Roselló, S.; Díez, M.J.; Nuez, F. Viral diseases causing the greatest economic losses to the tomato crop. I. The tomato spotted wilt virus—A review. Sci. Hort. 1996, 67, 117–150. [Google Scholar] [CrossRef]
- Kaye, A.C.; Moyer, J.W.; Parks, E.J.; Carbone, I.; Cubeta, M.A. Population genetic analysis of tomato spotted wilt virus on peanut in North Carolina and Virginia. Phytopathology 2011, 101, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.S. Replication of arboviruses in insect vectors. J. Comp. Pathol. 2000, 123, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G.; J, G.-J. Biological transmission of arboviruses: Reexamination of and new insights into components, mechanisms, and unique traits as well as their evolutionary trends. Clin. Microbiol. Rev. 2005, 18, 608–637. [Google Scholar] [CrossRef] [PubMed]
- Flegel, T.W. Update on viral accommodation, a model for host-viral interaction in shrimp and other arthropods. Dev. Comp. Immunol. 2007, 31, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cherry, S. Viruses and antiviral immunity in Drosophila. Dev. Comp. Immunol. 2014, 42, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.F. Advances in Disease Vector Research; Springer: New York, NY, USA, 1994. [Google Scholar]
- Cheng, G.; Liu, Y.; Wang, P.; Xiao, X. Mosquito defense strategies against viral infection. Trends Parasitol. 2016, 32, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.; Olson, K. The Role of RNA Interference (RNAi) in arbovirus-vector interactions. Viruses 2015, 7, 820–843. [Google Scholar] [CrossRef] [PubMed]
- Mongelli, V.; Saleh, M.-C. Bugs are not to be silenced: Small RNA pathways and antiviral responses in insects. Annu. Rev. Virol. 2016, 3, 573–589. [Google Scholar] [CrossRef] [PubMed]
- Gammon, D.B.; Mello, C.C. RNA interference-mediated antiviral defense in insects. Curr. Opin. Insect Sci. 2015, 8, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vodovar, N.; Saleh, M.C. Of insects and viruses: The role of small RNAs in insect defence. Adv. Insect Phys. 2012, 42, 1–36. [Google Scholar]
- Marques, J.T.; Imler, J.-L. The diversity of insect antiviral immunity: Insights from viruses. Curr. Opin. Microbiol. 2016, 32, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Fragkoudis, R.; Attarzadeh-Yazdi, G.; Nash, A.; Fazakerley, J.; Kohl, A. Advances in dissecting mosquito innate immune responses to arbovirus infection. J. Gen. Virol. 2009, 90, 2061–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donald, C.; Kohl, A.; Schnettler, E. New insights into control of arbovirus replication and spread by insect RNA interference pathways. Insects 2012, 3, 511–531. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Travanty, E.A.; Keene, K.M.; Franz, A.W.E.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA interference, arthropod-borne viruses, and mosquitoes. Virus Res. 2004, 102, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Bell-Sakyi, L.; Fazakerley, J.; Fragkoudis, R. Antiviral responses of arthropod vectors: An update on recent advances. Virusdisease 2014, 25, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: MicroRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-S.; Lai, E.C. Alternative miRNA biogenesis pathways and the interpretation of core miRNA pathway mutants. Mol. Cell 2011, 43, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Saj, A.; Lai, E.C. Control of microRNA biogenesis and transcription by cell signalling pathways. Curr. Opin. Genet. Dev. 2011, 21, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Gulyaeva, L.; Kushlinskiy, N. Regulatory mechanisms of microRNA expression. J. Transl. Med. 2016, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S. MicroRNA functions in insects. Insect Biochem. Mol. Biol. 2013, 43, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Lucas, K.; Raikhel, A.S. Insect microRNAs: Biogenesis, expression profiling and biological functions. Insect Biochem. Mol. Biol. 2013, 43, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D. MicroRNA: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Vidigal, J.A.; Ventura, A. The biological functions of miRNAs: Lessons from in vivo studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Ameres, S.L.; Zamore, P.D. Diversifying microRNA sequence and function. Nat. Rev. Mol. Cell Biol. 2013, 14, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R. Viruses and microRNAs: RISCy interactions with serious consequences. Genes Dev. 2011, 25, 1881–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, B.R. Viruses and microRNAs. Nat. Genet. 2006, 38, S25–S30. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R. MicroRNAs as mediators of viral evasion of the immune system. Nat. Immunol. 2013, 14, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottwein, E.; Cullen, B.R. Viral and cellular microRNAs as determinants of viral pathogenesis and immunity. Cell Host Microbe 2008, 3, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Umbach, J.L.; Cullen, B.R. The role of RNAi and microRNAs in animal virus replication and antiviral immunity. Genes Dev. 2009, 23, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Steitz, J. Virus meets host microrna: The destroyer, the booster, the hijacker. Mol. Cell. Biol. 2014, 34, 3780–3787. [Google Scholar] [CrossRef] [PubMed]
- tenOever, B.R. RNA viruses and the host microRNA machinery. Nat. Rev. Microbiol. 2013, 11, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Trobaugh, D.W.; Klimstra, W.B. microRNA regulation of RNA virus replication and pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Bruscella, P.; Bottini, S.; Baudesson, C.; Pawlotsky, J.; Feray, C.; Trabucchi, M. Viruses and miRNAs: More friends than foes. Front. Microbiol. 2017, 15, 824. [Google Scholar] [CrossRef] [PubMed]
- Liu, Sr.; Zhou, J.; Hu, C.G.; Wei, C.; Zhang, J. MicroRNA-mediated gene silencing in plant defense and viral counter-defense. Front. Microbiol. 2017, 8, 1801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. Plant microRNA: A small regulatory molecule with big impact. Dev. Biol. 2006, 289, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S. Regulatory role of cellular and viral microRNAs in insect-virus interactions. Curr. Opin. Insect Sci. 2015, 8, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Dykxhoorn, D.M. MicroRNAs in viral replication and pathogenesis. DNA Cell Biol. 2007, 26, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S. Role of microRNAs in insect host-microorganism interactions. Front. Physiol. 2011, 2, 48. [Google Scholar] [CrossRef] [PubMed]
- Scaria, V.; Hariharan, M.; Maiti, S.; Pillai, B.; Brahmachari, S.K. Host-virus interaction: A new role for microRNAs. Retrovirology 2006, 3, 68–68. [Google Scholar] [CrossRef] [PubMed]
- Libri, V.; Miesen, P.; van Rij, R.; Buck, A. Regulation of microRNA biogenesis and turnover by animals and their viruses. Cell. Mol. Life Sci. 2013, 70, 3525–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, M.A. microRNAs and the immune response. Trends Immunol. 2008, 29, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Lodish, H.F.; Zhou, B.; Liu, G.; Chen, C.Z. Micromanagement of the immune system by microRNAs. Nat. Rev. Immunol. 2008, 8, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, I.M.; Cheng, G.; Wieland, S.; Volinia, S.; Croce, C.M.; Chisari, F.V.; David, M. Interferon modulation of cellular microRNAs as an antiviral mechanism. Nature 2007, 449, 919–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsitsiou, E.; Lindsay, M.A. microRNAs and the immune response. Curr. Opin. Pharmacol. 2009, 9, 514–520. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; Baltimore, D. Physiological and pathological roles for microRNAs in the immune system. Nat. Rev. Immunol. 2010, 10, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Rajewsky, K. microRNA control in the immune system: Basic principles. Cell 2009, 136, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009, 10, R64–R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, C.L.; Harrison, T.; Hess, A.M.; Ebel, G.D. microRNA levels are modulated in Aedes aegypti after exposure to dengue-2. Insect Mol. Biol. 2014, 23, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, Y.; Wu, J.; Zheng, P.; Li, Y.; Zheng, X.; Puthiyakunnon, S.; Tu, Z.; Chen, X. The expression profile of Aedes albopictus miRNAs is altered by dengue virus serotype-2 infection. Cell Biosci. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Maharaj, P.D.; Widen, S.G.; Huang, J.; Wood, T.G.; Thangamani, S. Discovery of mosquito saliva microRNAs during CHIKV infection. PLoS Negl. Trop. Dis. 2015, 9, e0003386. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, M.; Etebari, K.; Hart, C.; Widen, S.; Wood, T.; Thangamani, S.; Asgari, S.; Hughes, G. Zika virus alters the microRNA expression profile and elicits an RNAi response in Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005760. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Li, C.; Zhang, Y.; Yan, T.; Zhu, X.; Zhao, M.; Xing, D.; Dong, Y.; Guo, X.; Zhao, T. Identification of microRNAs expressed in the midgut of Aedes albopictus during dengue infection. Parasites Vectors 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.; Rico-Hesse, R. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.A.; Glasner, D.R.; Shah, S.; Michlmayr, D.; Kramer, L.D.; Harris, E. Mosquito saliva increases endothelial permeability in the skin, immune cell migration, and dengue pathogenesis during antibody-dependent enhancement. PLoS Pathog. 2016, 12, e1005676. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.S.; Higgs, S. The enhancement of arbovirus transmission and disease by mosquito saliva is associated with modulation of the host immune response. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, A.W.E.; Kantor, A.; Passarelli, A.; Clem, R. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, S.; Du, J.; Gao, S.; Tian, Z.; Zheng, Y.; Liu, G.; Luo, J.; Yin, H. Analysis of the miRNA expression profile in an Aedes albopictus cell line in response to bluetongue virus infection. Infect. Genet. Evol. 2016, 39, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Shrinet, J.; Jain, S.; Jain, J.; Bhatnagar, R.; Sunil, S. Next generation sequencing reveals regulation of distinct Aedes microRNAs during chikungunya virus development. PLoS Negl. Trop. Dis. 2014, 8, e2616. [Google Scholar] [CrossRef] [PubMed]
- Miesen, P.; Ivens, A.; Buck, A.H.; van Rij, R.P. Small RNA profiling in dengue virus 2-infected Aedes mosquito cells reveals viral piRNAs and novel host miRNAs. PLoS Negl. Trop. Dis. 2016, 10, e0004452. [Google Scholar] [CrossRef] [PubMed]
- Skalsky Rebecca, L.; Vanlandingham Dana, L.; Scholle, F.; Higgs, S.; Cullen Bryan, R. Identification of microRNAs expressed in two mosquito vectors, Aedes albopictus and Culex quinquefasciatus. BMC Genom. 2010, 11, 119. [Google Scholar] [CrossRef]
- Li, J.-M.; Zhou, Y.-R.; Sun, Z.-T.; Wang, X.; Xie, L.; Chen, J.-P. Identification and profiling of conserved and novel microRNAs in Laodelphax striatellus in response to rice black-streaked dwarf virus (RBSDV) infection. Genom. Data 2015, 3, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Phillips, M.D.; Tyler, D.M.; Duan, H.; Chou, Y.-T.; Lai, E.C. The regulatory activity of microRNA* species has substantial influence on microRNA and 3′ UTR evolution. Nat. Struct. Mol. Biol. 2008, 15, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Ventoso, C.; Vora, M.; Driscoll, M. Sequence relationships among C. elegans, D. melanogaster and human microRNAs highlight the extensive conservation of microRNAs in biology. PLoS ONE 2008, 3, e2818. [Google Scholar] [CrossRef] [PubMed]
- Mullokandov, G.; Baccarini, A.; Ruzo, A.; Jayaprakash, A.D.; Tung, N.; Israelow, B.; Evans, M.J.; Sachidanandam, R.; Brown, B.D. High-throughput assessment of microRNA activity and function using microRNA sensor and decoy libraries. Nat. Methods 2012, 9, 840–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, B.D.; Gentner, B.; Cantore, A.; Colleoni, S.; Amendola, M.; Zingale, A.; Baccarini, A.; Lazzari, G.; Galli, C.; Naldini, L. Endogenous microRNA can be broadly exploited to regulate transgene expression according to tissue, lineage and differentiation state. Nat. Biotechnol. 2007, 25, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Hunt, S.; Ninova, M.; Griffiths-Jones, S.; Ronshaugen, M. Target repression induced by endogenous microRNAs: Large differences, small effects. PLoS ONE 2014, 9, e104286. [Google Scholar] [CrossRef] [PubMed]
- Chugh, P.; Dittmer, D.P. Potential pitfalls in microRNA profiling. Wiley Interdiscip. Rev. RNA 2012, 3, 601–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monsanto-Hearne, V.; Tham, A.L.Y.; Wong, Z.S.; Asgari, S.; Johnson, K.N. Drosophila miR-956 suppression modulates Ectoderm-expressed 4 and inhibits viral replication. Virology 2016, 502, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chan, E.Y.; Li, J.; Ni, C.; Peng, X.; Rosenzweig, E.; Tumpey, T.M.; Katze, M.G. MicroRNA expression and virulence in pandemic influenza virus-infected mice. J. Virol. 2010, 84, 3023–3032. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.-J.; Ren, G.; Liu, J.-L.; Zhao, Z.-A.; Yu, Y.-S.; Su, R.-W.; Ma, X.-H.; Ni, H.; Lei, W.; Yang, Z.-M. MicroRNA expression and regulation in mouse uterus during embryo implantation. J. Biol. Chem. 2008, 283, 23473–23484. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lowey, B.; Sodroski, C.; Krishnamurthy, S.; Alao, H.; Cha, H.; Chiu, S.; El-Diwany, R.; Ghany, M.; Liang, T. Cellular microRNA networks regulate host dependency of hepatitis C virus infection. Nat. Commun. 2017, 8, 1789–1789. [Google Scholar] [CrossRef] [PubMed]
- Dölken, L.; Malterer, G.; Erhard, F.; Kothe, S.; Friedel, C.C.; Suffert, G.; Marcinowski, L.; Motsch, N.; Barth, S.; Beitzinger, M.; et al. Systematic analysis of viral and cellular microrna targets in cells latently infected with human γ-herpesviruses by RISC immunoprecipitation assay. Cell Host Microbe 2010, 7, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Buggele, W.; Krause, K.; Horvath, C. Small RNA profiling of Influenza A virus-infected cells identifies miR-449b as a regulator of Histone Deacetylase 1 and Interferon Beta. PLoS ONE 2013, 8, e76560. [Google Scholar] [CrossRef] [PubMed]
- Vergoulis, T.; Vlachos, I.S.; Alexiou, P.; Georgakilas, G.; Maragkakis, M.; Reczko, M.; Gerangelos, S.; Koziris, N.; Dalamagas, T.; Hatzigeorgiou, A.G. TarBase 6.0: Capturing the exponential growth of miRNA targets with experimental support. Nucleic Acids Res. 2012, 40, D222–D229. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-D.; Tseng, Y.-T.; Shrestha, S.; Lin, Y.-L.; Khaleel, A.; Chou, C.-H.; Chu, C.-F.; Huang, H.-Y.; Lin, C.-M.; Ho, S.-Y.; et al. MiRTarBase update 2014: An information resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2014, 42, D78–D85. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Zuo, Z.; Cai, G.; Kang, S.; Gao, X.; Li, T. MiRecords: An integrated resource for microRNA-target interactions. Nucleic Acids Res. 2009, 37, D105–D110. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Gu, J.; Wang, T.; Ding, Z. OncomiRDB: A database for the experimentally verified oncogenic and tumor-suppressive microRNAs. Bioinformatics 2014, 30, 2237–2238. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liu, Y.; Yan, H.; Li, Y.; Zhang, H.; Xu, J.; Puthiyakunnon, S.; Chen, X. miR-281, an abundant midgut-specific miRNA of the vector mosquito Aedes albopictus enhances dengue virus replication. Parasit Vectors 2014, 7, 488–488. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Frentiu, F.D.; Moreira, L.A.; O’Neill, S.L.; Asgari, S. Wolbachia uses host microRNAs to manipulate host gene expression and facilitate colonization of the dengue vector Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 9250–9255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hussain, M.; Asgari, S. Regulation of arginine methyltransferase 3 by a Wolbachia-induced microRNA in Aedes aegypti and its effect on Wolbachia and dengue virus replication. Insect Biochem. Mol. Biol. 2014, 53, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia uses a host microRNA to regulate transcripts of a methyltransferase, contributing to dengue virus inhibition in Aedes aegypti. Proc. Natl. Acad. Sci. USA 2013, 110, 10276–10281. [Google Scholar] [CrossRef] [PubMed]
- Slonchak, A.; Hussain, M.; Torres, S.; Asgari, S.; Khromykh, A.A. Expression of mosquito microRNA aae-miR-2940-5p is down-regulated in response to West Nile virus infection to restrict viral replication. J. Virol. 2014, 88, 8457–8467. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Yoon, S. Got target? Computational methods for microRNA target prediction and their extension. Exp. Mol. Med. 2010, 42, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Sanchez, A.; Murphy, C. MicroRNA target identification: Experimental approaches. Biology 2013, 2, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Ekimler, S.; Sahin, K. Computational methods for microRNA target prediction. Genes 2014, 5, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.E.; Martin, M.M.; Feldman, D.S.; Terry, A.V.; Nuovo, G.J.; Elton, T.S. Experimental validation of miRNA targets. Methods 2008, 44, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, D.W.; Bracken, C.P.; Goodall, G.J. Experimental strategies for microRNA target identification. Nucleic Acids Res. 2011, 39, 6845–6853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenvang, J.; Petri, A.; Lindow, M.; Obad, S.; Kauppinen, S. Inhibition of microRNA function by antimiR oligonucleotides. Silence 2012, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinkraus, B.R.; Toegel, M.; Fulga, T.A. Tiny giants of gene regulation: Experimental strategies for microRNA functional studies. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 311–362. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, B.; Hussain, M.; Asgari, S. An insect trypsin-like serine protease as a target of microRNA: Utilization of microRNA mimics and inhibitors by oral feeding. Insect Biochem. Mol. Biol. 2012, 43, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Osei-Amo, S.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia-induced aae-miR-12 miRNA negatively regulates the expression of MCT1 and MCM6 genes in Wolbachia-infected mosquito cell line. PLoS ONE 2012, 7, e50049–e50049. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Walker, T.; O’Neill, S.L.; Asgari, S. Blood meal induced microRNA regulates development and immune associated genes in the dengue mosquito vector, Aedes aegypti. Insect Biochem. Mol. Biol. 2013, 43, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhou, Y.; Liu, Y.; Deng, Y.; Chen, X. miR-252 of the Asian tiger mosquito Aedes albopictus regulates dengue virus replication by suppressing the expression of the dengue virus envelope protein. J. Med. Virol. 2014, 86, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Ferguson, E.L.; Sternberg, P.W.; Horvitz, H.R. A genetic pathway for the specification of the vulval cell lineages of Caenorhabditis elegans. Nature 1987, 326, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.M.; Thompson, J.A.; Ufkin, M.L.; Sathyanarayana, P.; Liaw, L.; Congdon, C.B. Common features of microRNA target prediction tools. Front. Genet. 2014, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.P.; Cho, J.H.; Hood, L.; Franco, O.L.; Pereira, R.W.; Wang, K. A review of computational tools in microRNA discovery. Front. Genet. 2013, 4, 81. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D. Using Drosophila as a model insect. Nat. Rev. Genet. 2000, 1, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hanson, F.B.; Ferris, F.R. A quantitative study of fecundity in Drosophila melanogaster. J. Exp. Zool. 1929, 54, 485–506. [Google Scholar] [CrossRef]
- Shapiro, H. The rate of oviposition in the fruit fly, Drosophila. Biol. Bull. 1932, 63, 456–471. [Google Scholar] [CrossRef]
- Markow, T.A. “Cost” of virginity in wild Drosophila melanogaster females. Ecol. Evol. 2011, 1, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.; Allen, A.; Bardin, J.; Christian, M.; Daimon, K.; Dozier, K.; Hansen, C.; Holcomb, L.; Ahlander, J. Drosophila as a genetic model for studying pathogenic human viruses. Virology 2012, 423, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Dow, J. Integrative physiology, functional genomics and the phenotype gap: A guide for comparative physiologists. J. Exp. Biol. 2007, 210, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Perrimon, N.; Ni, J.-Q.; Perkins, L. In vivo RNAi: Today and tomorrow. Cold Spring Harb. Perspect. Biol. 2010, 2, a003640. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.; Pearson, R.K.; Bellen, H.J.; Kane, C.J.; Grossniklaus, U.; Gehring, W.J. P-element-mediated enhancer detection: An efficient method for isolating and characterizing developmentally regulated genes in Drosophila. Genes Dev. 1989, 3, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, T.B.; Krasnow, M.A. The Drosophila genome sequence: Implications for biology and medicine. Science 2000, 287, 2218–2220. [Google Scholar] [CrossRef] [PubMed]
- Rubin, G.; Yandell, M.D.; Wortman, J.R.; Miklos, G.; Nelson, C.R.; Hariharan, I.; Fortini, M.E.; Li, P.; Apweiler, R.; Fleischmann, W.; et al. Comparative genomics of the eukaryotes. Science 2000, 287, 2204–2215. [Google Scholar] [CrossRef] [PubMed]
- Swevers, L.; Liu, J.; Smagghe, G. Defense mechanisms against viral infection in Drosophila: RNAi and non-RNAi. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Panayidou, S.; Ioannidou, E.; Apidianakis, Y. Human pathogenic bacteria, fungi, and viruses in Drosophila: Disease modeling, lessons, and shortcomings. Virulence 2014, 5, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Chotkowski, H.L.; Ciota, A.T.; Jia, Y.; Puig-Basagoiti, F.; Kramer, L.D.; Shi, P.-Y.; Glaser, R.L. West Nile virus infection of Drosophila melanogaster induces a protective RNAi response. Virology 2008, 377, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Filone, C.M.; Hanna, S.L.; Caino, M.C.; Bambina, S.; Doms, R.W.; Cherry, S. Rift Valley fever virus infection of human cells and insect hosts is promoted by protein Kinase C Epsilon. PLoS ONE 2010, 5, e15483. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, I.; Jansen, S.; Lorenzen, S.; Martin, R.; Huber, K.; Heitmann, A.; Watson, M.; Brennan, B.; Elliott, R.; Sall, A.; et al. RNA interference restricts Rift Valley fever virus in multiple insect systems. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Mudiganti, U.; Hernandez, R.; Ferreira, D.; Brown, D.T. Sindbis virus infection of two model insect cell systems—A comparative study. Virus Res. 2006, 122, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Hanley, K.A. RNA interference modulates replication of dengue virus in Drosophila melanogaster cells. BMC Microbiol. 2010, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.E.; Veronesi, E.; Maurin, G.; Ftaich, N.; Guiguen, F.; Rixon, F.; Ratinier, M.; Mertens, P.; Carpenter, S.; Palmarini, M.; et al. Drosophila melanogaster as a model organism for bluetongue virus replication and tropism. J. Virol. 2012, 86, 9015–9024. [Google Scholar] [CrossRef] [PubMed]
- Karlikow, M.; Goic, B.; Saleh, M.C. RNAi and antiviral defense in Drosophila: Setting up a systemic immune response. Dev. Comp. Immunol. 2014, 42, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Huszar, T.; Imler, J.L. Drosophila viruses and the study of antiviral host-defense. Adv. Virus Res. 2008, 72, 227–265. [Google Scholar] [PubMed]
- Lamiable, O.; Imler, J.-L. Induced antiviral innate immunity in Drosophila. Curr. Opin. Microbiol. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejarano, F.; Bortolamiol-Becet, D.; Dai, Q.; Sun, K.; Saj, A.; Chou, Y.T.; Raleigh, D.R.; Kim, K.; Ni, J.Q.; Duan, H.; et al. A genome-wide transgenic resource for conditional expression of Drosophila microRNAs. Development 2012, 139, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Schertel, C.; Rutishauser, T.; Forstemann, K.; Basler, K. Functional characterization of Drosophila microRNAs by a novel in vivo library. Genetics 2012, 192, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Fulga, T.A.; McNeill, E.M.; Binari, R.; Yelick, J.; Blanche, A.; Booker, M.; Steinkraus, B.R.; Schnall-Levin, M.; Zhao, Y.; DeLuca, T.; et al. A transgenic resource for conditional competitive inhibition of conserved Drosophila microRNAs. Nat. Commun. 2015, 6, 7279–7279. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Song, S.; Weng, R.; Verma, P.; Kugler, J.M.; Buescher, M.; Rouam, S.; Cohen, S. Systematic study of Drosophila microRNA functions using a collection of targeted knockout mutations. Dev. Cell 2014, 31, 784–800. [Google Scholar] [CrossRef] [PubMed]
- Belinda, L.W.-C.; Wei, W.X.; Hanh, B.T.H.; Lei, L.X.; Bow, H.; Ling, D.J. SARM: A novel Toll-like receptor adaptor, is functionally conserved from arthropod to human. Mol. Immunol. 2008, 45, 1732–1742. [Google Scholar] [CrossRef] [PubMed]
- Guillem, Y.; Bastian, F.; Maria-Dolors, P.; Xavier, B. The microRNA toolkit of insects. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]


{kind=link}
| Ae. aegypti | Ae. albopictus | Culex | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Whole Insect | Insect Parts | Cell | Whole Insect | Insect Parts | Cell | Whole Insect | Insect Parts | Cells | |
| BTV | 0.5 dpi [73] | ||||||||
| CHIKV | saliva, 10 dpi * [66] | 1 dpi [74] | saliva, 10 dpi * [66] | ||||||
| DENV | 2, 4, 9 dpi [64] | 3 dpi [75] | 7 dpi * [65] | midgut, 1 dpi [68] | |||||
| WNV | 14 dpi [76] | ||||||||
| ZIKV | 2, 7, 14 dpi [67] | ||||||||
| Aae-ZIKV | Aae-DENV | Aal-DENV | Aal-BTV | Aal-CHIKV | Aal-DENV * | Aae-CHIKV ^ | Aal-CHIKV ^ | Aae-ZIKV | Aae-DENV | Aal-DENV | Aal-BTV | Aal-CHIKV | Aal-DENV * | Aae-CHIKV ^ | Aal-CHIKV ^ | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bantam/bantam-3p | ↓ | ↓ | ↑ | ↓ | miR-1 | ↓ | ||||||||||||
| miR-263a/miR-263a-5p | ↓ | ↓ | ↑ | ↑ | miR-1-5p | ↑ | ||||||||||||
| miR-34/miR-34-5p | ↑ | ↓ | ↑ | ↓ | miR-1174 | ↓ | ||||||||||||
| let-7 | ↓ | ↓ | ↑ | miR-1175-3p | ↓ | |||||||||||||
| miR-1000/miR-1000-5p | ↓ | ↓ | ↑ | miR-12 | ↑ | |||||||||||||
| miR-275/miR-275-3p | ↓ | ↑ | ↑ | miR-124-5p | ↓ | |||||||||||||
| miR-276/miR-276-3p | ↓ | ↑ | ↑ | miR-13 | ↑ | |||||||||||||
| miR-281/miR-281-5p | ↓ | ↓ | ↑ | miR-15b | ↑ | |||||||||||||
| miR-2941 | ↓ | ↓ | ↓ | miR-1767 | ↑ | |||||||||||||
| miR-305/miR-305-5p | ↓ | ↓ | ↑ | miR-1889 | ↑ | |||||||||||||
| miR-315 | ↓ | ↑ | ↑ | miR-190 | ↑ | |||||||||||||
| miR-317 | ↓ | ↓ | ↑ | miR-193-5p | ↑ | |||||||||||||
| miR-8/miR-8-3p | ↓ | ↓ | ↓ | miR-252 | ↑ | |||||||||||||
| miR-927 | ↓ | ↑ | ↑ | miR-275-5p | ↑ | |||||||||||||
| miR-957 | ↓ | ↑ | ↑ | miR-276-1 | ↑ | |||||||||||||
| miR-989 | ↑ | ↓ | ↓ | miR-277-5p | ↓ | |||||||||||||
| miR-999 | ↓ | ↑ | ↑ | miR-279 | ↑ | |||||||||||||
| miR-308/miR-308-5p | ↓ | ↑ | ↑ | miR-281a-3p | ↓ | |||||||||||||
| miR-10 | ↑ | ↑ | miR-281a-5p | ↓ | ||||||||||||||
| miR-100 | ↑ | ↑ | miR-281b-3p | ↓ | ||||||||||||||
| miR-125 | ↑ | ↑ | miR-281c-5p | ↓ | ||||||||||||||
| miR-14 | ↑ | ↑ | miR-283 | ↓ | ||||||||||||||
| miR-184 | ↓ | ↓ | miR-285 | ↑ | ||||||||||||||
| miR-1889-5p | ↓ | ↓ | miR-286a | ↓ | ||||||||||||||
| miR-1890 | ↓ | ↑ | miR-286b | ↓ | ||||||||||||||
| miR-210 | ↓ | ↑ | miR-2944b-5p | ↓ | ||||||||||||||
| miR-263b/miR-263b-5p | ↓ | ↑ | miR-2a | ↑ | ||||||||||||||
| miR-276-5p | ↓ | ↑ | miR-2c | ↑ | ||||||||||||||
| mir-277 | ↑ | ↑ | miR-2c-3p | ↓ | ||||||||||||||
| miR-281-2-5p | ↓ | ↑ | miR-307 | ↑ | ||||||||||||||
| miR-2940/miR-2940-5p | ↓ | ↓ | miR-309a | ↓ | ||||||||||||||
| miR-2940-3p | ↑ | ↓ | miR-33-5p | ↓ | ||||||||||||||
| miR-2944b-3p | ↓ | ↓ | miR-3368-5p | ↓ | ||||||||||||||
| miR-2945-3p | ↓ | ↑ | miR-34-3p | ↑ | ||||||||||||||
| miR-2946 | ↓ | ↓ | miR-3722-5p | ↓ | ||||||||||||||
| miR-2b | ↓ | ↑ | miR-3811e-5p | ↑ | ||||||||||||||
| miR-306/miR-306-5p | ↓ | ↑ | miR-4275-5p | ↓ | ||||||||||||||
| miR-308-3p | ↓ | ↓ | miR-4448 | ↓ | ||||||||||||||
| miR-375 | ↓ | ↓ | miR-4728-5p | ↑ | ||||||||||||||
| miR-71/miR-71-5p | ↑ | ↑ | miR-5108-5p | ↓ | ||||||||||||||
| miR-980/miR-980-3p | ↓ | ↑ | miR-5119-5p | ↑ | ||||||||||||||
| miR-9a | ↑ | ↑ | miR-6134 | ↑ | ||||||||||||||
| miR-9c-5p | ↓ | ↓ | miR-622 | ↑ | ||||||||||||||
| miR-133 | ↑ | ↑ | miR-79c-3p | ↓ | ||||||||||||||
| miR-1891 | ↑ | ↑ | miR-8-5p | ↓ | ||||||||||||||
| mir-92b | ↑ | ↑ | miR-87-5p | ↑ | ||||||||||||||
| miR-927a | ↑ | |||||||||||||||||
| miR-92a | ↑ | |||||||||||||||||
| miR-932 | ↑ | |||||||||||||||||
| miR-932-5p | ↓ | |||||||||||||||||
| miR-970 | ↑ | |||||||||||||||||
| miR-988-5p | ↑ | |||||||||||||||||
| miR-993 | ↑ | |||||||||||||||||
| miR-996 | ↑ | |||||||||||||||||
| miR-998 | ↑ | |||||||||||||||||
| miR-9b | ↓ | |||||||||||||||||
| miR-iab | ↓ | |||||||||||||||||
| virus | miRNA | miRNA Regulation during Viral Infection | virus Accumulation upon Mimic Treatment | virus Accumulation upon Inhibitor Treatment | Target/Function | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ae. aegypti | Ae. albopictus | 1 dpi | 2 dpi | 3 dpi | 4 dpi | 5 dpi | 1 dpi | 2 dpi | 3 dpi | 4 dpi | 5 dpi | |||
| CHIKV | miR-12[66] | ↑s10 dpi | ≈ s 10 dpi | ↑# | ↓ # | targets MCM6 & MCT1 [107] | ||||||||
| miR-125[66] | ↑ s10 dpi | ↑ s10 dpi | ↑ # | ↓ # | ↑ # | |||||||||
| miR-184[66] | ↓ s10 dpi | ≈ s 10 dpi | ↓ | ↓ # | ||||||||||
| miR-375[66] | ↓ s10 dpi | ⊗s 10 dpi | ↓ # | enhances DENV infection [108], up-regulates cactus, down-regulates REL1 [108] | ||||||||||
| miR-2940[66] | ≈ s10 dpi | ≈ s10 dpi | ↑ # | ↓ # | up-regulates metalloprotease m41 ftsh [95], arginine methyltransferase [96], & DNA methyltransferase [97] | |||||||||
| WNV | miR-2940-5p[98] | ↓3 dpi & 5 dpi | ↑ | ↑ | ↑ | ↑ | ↓ | ↓ | ||||||
| DENV | miR-252[109] | ↑1 dpi & 3 dpi | ↓ | ↓ | ↑ | ↑ | targets DENV-2 E protein [109] | |||||||
| miR-281[94] | ↑ 4 dpi↑wm & m 4 dpi & 7 dpi | ↑ | ↓ | ↓ * | targets 5′-UTR of DENV2 to enhance viral replication [94] | |||||||||
| miR-4728m[68] | ↑ wm1 dpi | ↑ | ↑ ** | ↓ | ||||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monsanto-Hearne, V.; Johnson, K.N. miRNAs in Insects Infected by Animal and Plant Viruses. Viruses 2018, 10, 354. https://doi.org/10.3390/v10070354
Monsanto-Hearne V, Johnson KN. miRNAs in Insects Infected by Animal and Plant Viruses. Viruses. 2018; 10(7):354. https://doi.org/10.3390/v10070354
Chicago/Turabian StyleMonsanto-Hearne, Verna, and Karyn N. Johnson. 2018. "miRNAs in Insects Infected by Animal and Plant Viruses" Viruses 10, no. 7: 354. https://doi.org/10.3390/v10070354
APA StyleMonsanto-Hearne, V., & Johnson, K. N. (2018). miRNAs in Insects Infected by Animal and Plant Viruses. Viruses, 10(7), 354. https://doi.org/10.3390/v10070354