Anti-Influenza A Viral Butenolide from Streptomyces sp. Smu03 Inhabiting the Intestine of Elephas maximus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Analytical Instruments
2.2. Cells and Influenza Viral Strains
2.3. Fermentation, Extraction and Isolation
2.4. Structure of (4S)-4-Hydroxy-10-methyl-11-oxo-dodec-2-en-1,4-olide (1)
2.5. Cytotoxic Assay
2.6. Cytopathic Effect (CPE) Reduction Assay
2.7. CPE Reduction Assay with Different Drug Administrations
2.8. Quantitative Real-Time Polymerase Chain Reaction (PCR) Assay
2.9. Fluorescence Confocal Microscopy Assay
2.10. Neuraminidase (NA) Inhibition Assay
2.11. Hemolysis Inhibition Assay
2.12. Polykaryon Formation Inhibition Assay
2.13. In Vivo Protective Efficacy in Mice
2.14. Statistical Analysis
3. Results
3.1. A Butenolide Was Isolated from Streptomyces sp. Smu03 Residing in the Intestine of Elephas maximus
3.2. Butenolide 1 is Active against a Wide Variety of Influenza A Viruses and has Moderate Cellular Toxicity Toward MDCK Cells
3.3. Butenolide 1 Displays the Anti-Influenza A Virus (IAV) Effect in the Early Stage of Infection
3.4. Butenolide 1 is Inactive toward the Enzymatic Activity of NA
3.5. The HA2 Subunit of Hemagglutinin (HA) is the Possible Target of Butenolide 1
3.6. Butenolide 1 is Effective against IAV In Vivo
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kieft, T.L.; Simmons, K.A. Allometry of animal-microbe interactions and global census of animal-associated microbes. Proc. Biol. Sci. 2015, 282, 20150702. [Google Scholar] [CrossRef] [PubMed]
- Mafla, S.; Moraga, R.; Leon, C.G.; Guzman-Fierro, V.G.; Yanez, J.; Smith, C.T.; Mondaca, M.A.; Campos, V.L. Biodegradation of roxarsone by a bacterial community of underground water and its toxic impact. World J. Microbiol. Biotechnol. 2015, 31, 1267–1277. [Google Scholar] [CrossRef] [PubMed]
- Klebe, G.; Schlitzer, M. M2 inhibitors and neuraminidase inhibitors. Pharm. Unserer Zeit 2011, 40, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, R.W.; Aitken, A.; Calder, L.J.; Martin, S.R.; Skehel, J.J.; Wharton, S.A.; Weis, W.; Wiley, D.C. Studies on the structure of the influenza virus haemagglutinin at the PH of membrane fusion. J. Gen. Virol. 1988, 69 Pt 11, 2785–2795. [Google Scholar] [CrossRef] [PubMed]
- Sauter, N.K.; Bednarski, M.D.; Wurzburg, B.A.; Hanson, J.E.; Whitesides, G.M.; Skehel, J.J.; Wiley, D.C. Hemagglutinins from two influenza virus variants bind to sialic acid derivatives with millimolar dissociation constants: A 500-MHz proton nuclear magnetic resonance study. Biochemistry 1989, 28, 8388–8396. [Google Scholar] [CrossRef] [PubMed]
- Gething, M.J.; Doms, R.W.; York, D.; White, J. Studies on the mechanism of membrane fusion: Site-specific mutagenesis of the hemagglutinin of influenza virus. J. Cell Biol. 1986, 102, 11–23. [Google Scholar] [CrossRef] [PubMed]
- White, J.M.; Wilson, I.A. Anti-peptide antibodies detect steps in a protein conformational change: Low-ph activation of the influenza virus hemagglutinin. J. Cell Biol. 1987, 105, 2887–2896. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Colonno, R.; Krystal, M. Characterization of a hemagglutinin-specific inhibitor of influenza A virus. Virology 1996, 226, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, R.; Zhang, R.; Chan, C.C.; Xi, B.; Zhu, Z.; Yang, J.; Poon, V.K.; Zhou, J.; Chen, M.; et al. Cl-385319 inhibits H5N1 avian influenza A virus infection by blocking viral entry. Eur. J. Pharmacol. 2011, 660, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.L.; Bridges, O.A.; Seiler, P.; Kim, J.K.; Yen, H.L.; Salomon, R.; Govorkova, E.A.; Webster, R.G.; Russell, C.J. The PH of activation of the hemagglutinin protein regulates H5N1 influenza virus pathogenicity and transmissibility in ducks. J. Virol. 2010, 84, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.W.; Lee, H.S.; Rho, J.R.; Kim, T.S.; Mo, S.J.; Shin, J. New lactone-containing metabolites from a marine-derived bacterium of the genus streptomyces. J. Nat. Prod. 2001, 64, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Mukku, V.J.; Speitling, M.; Laatsch, H.; Helmke, E. New butenolides from two marine streptomycetes. J. Nat. Prod. 2000, 63, 1570–1572. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Tan, G.S.; Mullarkey, C.E.; Lee, A.J.; Lam, M.M.; Krammer, F.; Henry, C.; Wilson, P.C.; Ashkar, A.A.; Palese, P.; et al. Epitope specificity plays a critical role in regulating antibody-dependent cell-mediated cytotoxicity against influenza a virus. Proc. Natl. Acad. Sci. USA 2016, 113, 11931–11936. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Li, F.; Wu, Q.; Xie, X.; Wu, W.; Wu, J.; Chen, Q.; Liu, S.; He, J. A “building block” approach to the new influenza a virus entry inhibitors with reduced cellular toxicities. Sci. Rep. 2016, 6, 22790. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.K.; Cheung, N.N.; Yang, F.; Dai, J.; Liu, L.; Chen, Z.; Sze, K.H.; Chen, H.; Yuen, K.Y.; Kao, R.Y. Identification of novel fusion inhibitors of influenza a virus by chemical genetics. J. Virol. 2015, 90, 2690–2701. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Luo, Y.; Yang, G.; Li, F.; Xie, X.; Chen, D.; He, L.; Wang, J.; Ye, C.; Lu, S.; et al. Potent influenza a virus entry inhibitors targeting a conserved region of hemagglutinin. Biochem. Pharmacol. 2017, 144, 35–51. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Pei, Y.; Chen, S.; Li, S.; Hu, X.; Chen, G.; Lin, B.; Wang, H. Three new butenolides from the fungus Aspergillus sp. CBS-P-2. Mol. (Basel, Switzerland) 2016, 21, 1361. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, Y.; Ma, K.X.; Li, Y.Q.; Huang, R.; Xie, X.S.; Wu, S.H. Two new butenolides produced by an actinomycete Streptomyces sp. Chem. Biodivers. 2014, 11, 929–933. [Google Scholar] [CrossRef] [PubMed]
- Li, D.H.; Zhu, T.J.; Liu, H.B.; Fang, Y.C.; Gu, Q.Q.; Zhu, W.M. Four butenolides are novel cytotoxic compounds isolated from the marine-derived bacterium, streptoverticillium luteoverticillatum 11014. Arch. Pharm. Res. 2006, 29, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, T.A.; Hong, J.; Lee, C.O.; Sim, C.J.; Im, K.S.; Lee, D.S.; Jung, J.H. New cytotoxic metabolites from a marine sponge Homaxinella sp. J. Nat. Prod. 2004, 67, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.; Carlsson, M.; Uvell, H.; Islam, K.; Edlund, K.; Cullman, I.; Altermark, B.; Mei, Y.F.; Elofsson, M.; Willassen, N.P.; et al. Isolation and characterization of anti-adenoviral secondary metabolites from marine actinobacteria. Mar. Drugs 2014, 12, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Serkedjieva, J.; Hay, A.J. In vitro anti-influenza virus activity of a plant preparation from Geranium sanguineum L. Antivir. Res. 1998, 37, 121–130. [Google Scholar] [CrossRef]
- Nobusawa, E. [Structure and function of the hemagglutinin of influenza viruses]. Nihon Rinsho Jpn. J. Clin. Med. 1997, 55, 2562–2569. [Google Scholar]
- Bantia, S.; Ghate, A.A.; Ananth, S.L.; Babu, Y.S.; Air, G.M.; Walsh, G.M. Generation and characterization of a mutant of influenza a virus selected with the neuraminidase inhibitor BCX-140. Antimicrob. Agents Chemother. 1998, 42, 801–807. [Google Scholar] [PubMed]
- Vareckova, E.; Mucha, V.; Wharton, S.A.; Kostolansky, F. Inhibition of fusion activity of influenza A haemagglutinin mediated by HA2-specific monoclonal antibodies. Arch. Virol. 2003, 148, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Epand, R.F.; Leikina, E.; Epand, R.M.; Chernomordik, L.V. The final conformation of the complete ectodomain of the HA2 subunit of influenza hemagglutinin can by itself drive low PH-dependent fusion. J. Biol. Chem. 2011, 286, 13226–13234. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.B. Avian flu to human influenza. Annu. Rev. Med. 2006, 57, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, G.T. Natural products and pharma 2011: Strategic changes spur new opportunities. Nat. Prod. Rep. 2011, 28, 1783–1789. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.B.; Tiwari, V.K. Natural products: An evolving role in future drug discovery. Eur. J. Med. Chem. 2011, 46, 4769–4807. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yao, Z.; Shen, X.; Chen, Z.; Liu, X.; Parquette, J.R.; Liu, S. Oligothiophene compounds inhibit the membrane fusion between H5N1 avian influenza virus and the endosome of host cell. Eur. J. Med. Chem. 2017, 130, 185–194. [Google Scholar] [CrossRef] [PubMed]
- McCullough, C.; Wang, M.; Rong, L.; Caffrey, M. Characterization of influenza hemagglutinin interactions with receptor by NMR. PLoS ONE 2012, 7, e33958. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, M.; Shen, X.; Liu, S. Influenza a virus entry inhibitors targeting the hemagglutinin. Viruses 2013, 5, 352–373. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
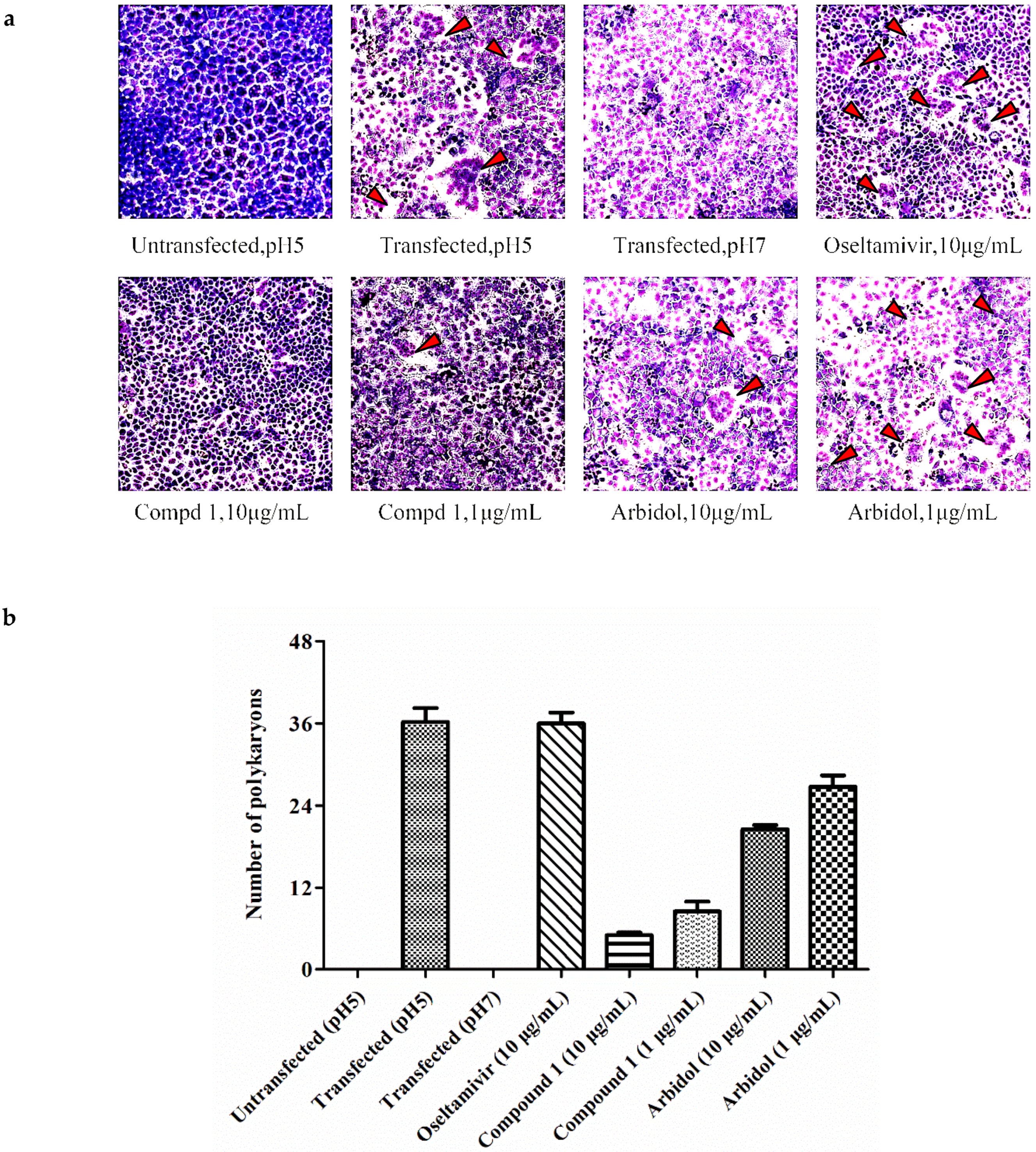{kind=link}
{kind=link}
| Name | IC50 ± SD a | CC50 ± SD b | ||||||
|---|---|---|---|---|---|---|---|---|
| H3N2 c | H1N1 d | H1N1 e | H1N1 f | H3 (690) g | H3 (699) | B | ||
| 1 | 7.59 ± 0.72 | 6.10 ± 1.30 | 0.29 ± 0.06 | 3.60 ± 0.43 | 5.45 ± 1.83 | 4.59 ± 0.43 | 11.69 ± 0.92 | 38.07 ± 1.50 |
| Arbidol | >10 | 5.58 ± 0.69 | 0.45 ± 0.16 | NT h | 6.51 ± 170 | 3.85 ± 0.70 | 24.58 ± 0.28 | 12.97 ± 2.25 |
| Name | IC50 ± SD (µg/mL) | |||
|---|---|---|---|---|
| Pretreatment to Cell | Pretreatment to Virus | During Infection | After Infection | |
| Compound 1 | 0.96 ± 0.10 | 0.29 ± 0.06 | 7.50 ± 0.59 | >25 |
| Arbidol | >25 | 0.45 ± 0.16 | 6.80 ± 1.45 | >25 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Chen, D.; Lu, S.; Yang, G.; Zhang, X.; Chen, Z.; Fan, S.; Wu, S.; He, J. Anti-Influenza A Viral Butenolide from Streptomyces sp. Smu03 Inhabiting the Intestine of Elephas maximus. Viruses 2018, 10, 356. https://doi.org/10.3390/v10070356
Li F, Chen D, Lu S, Yang G, Zhang X, Chen Z, Fan S, Wu S, He J. Anti-Influenza A Viral Butenolide from Streptomyces sp. Smu03 Inhabiting the Intestine of Elephas maximus. Viruses. 2018; 10(7):356. https://doi.org/10.3390/v10070356
Chicago/Turabian StyleLi, Fangfang, Daiwei Chen, Shengsheng Lu, Guang Yang, Xiaoling Zhang, Zhao Chen, Sheng Fan, Shaohua Wu, and Jian He. 2018. "Anti-Influenza A Viral Butenolide from Streptomyces sp. Smu03 Inhabiting the Intestine of Elephas maximus" Viruses 10, no. 7: 356. https://doi.org/10.3390/v10070356
APA StyleLi, F., Chen, D., Lu, S., Yang, G., Zhang, X., Chen, Z., Fan, S., Wu, S., & He, J. (2018). Anti-Influenza A Viral Butenolide from Streptomyces sp. Smu03 Inhabiting the Intestine of Elephas maximus. Viruses, 10(7), 356. https://doi.org/10.3390/v10070356