Anti-Infectivity against Herpes Simplex Virus and Selected Microbes and Anti-Inflammatory Activities of Compounds Isolated from Eucalyptus globulus Labill.


 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phytochemistry
2.1.1. Plant Material
2.1.2. General Experimental Procedure
2.1.3. Extraction and Isolation
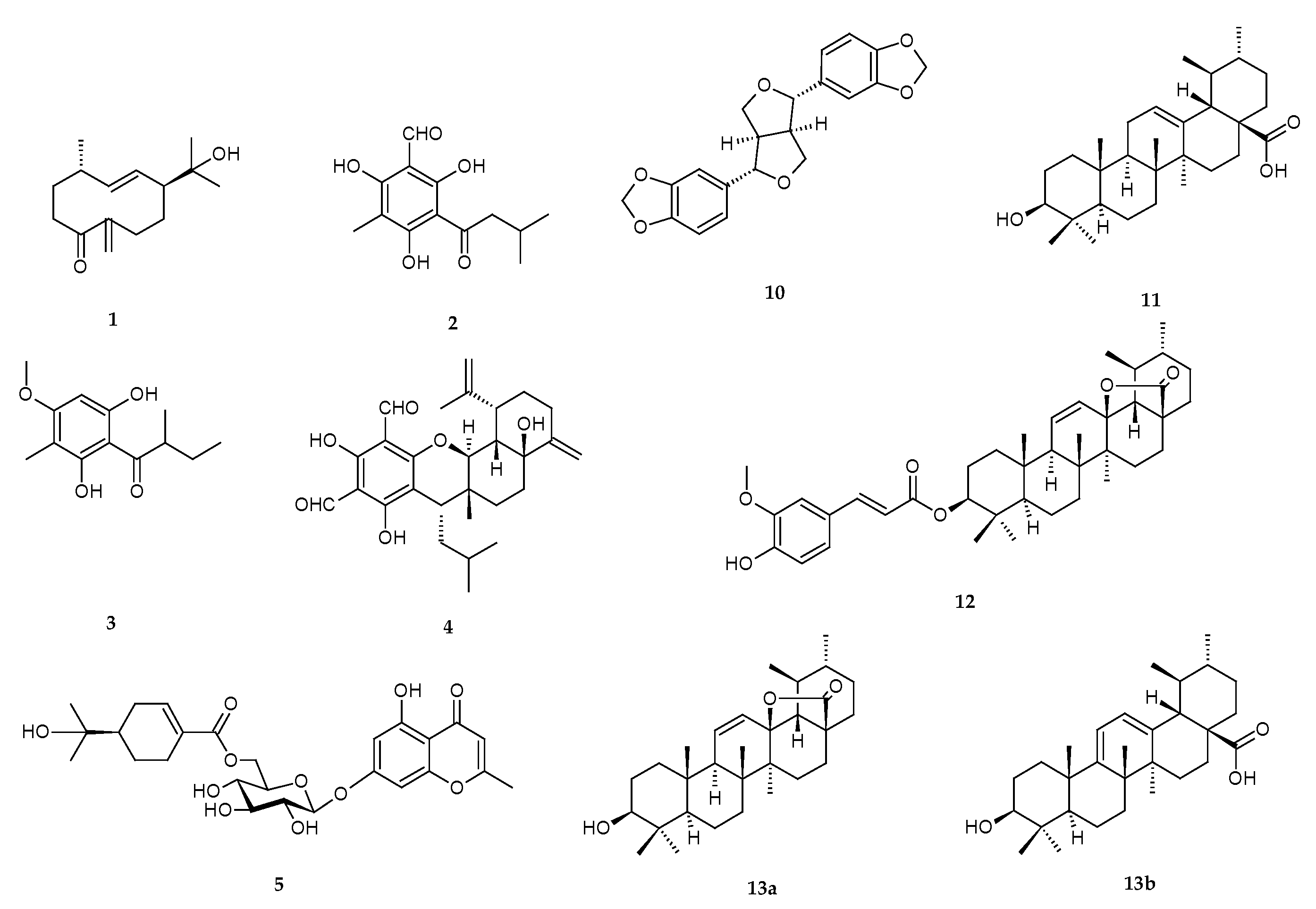
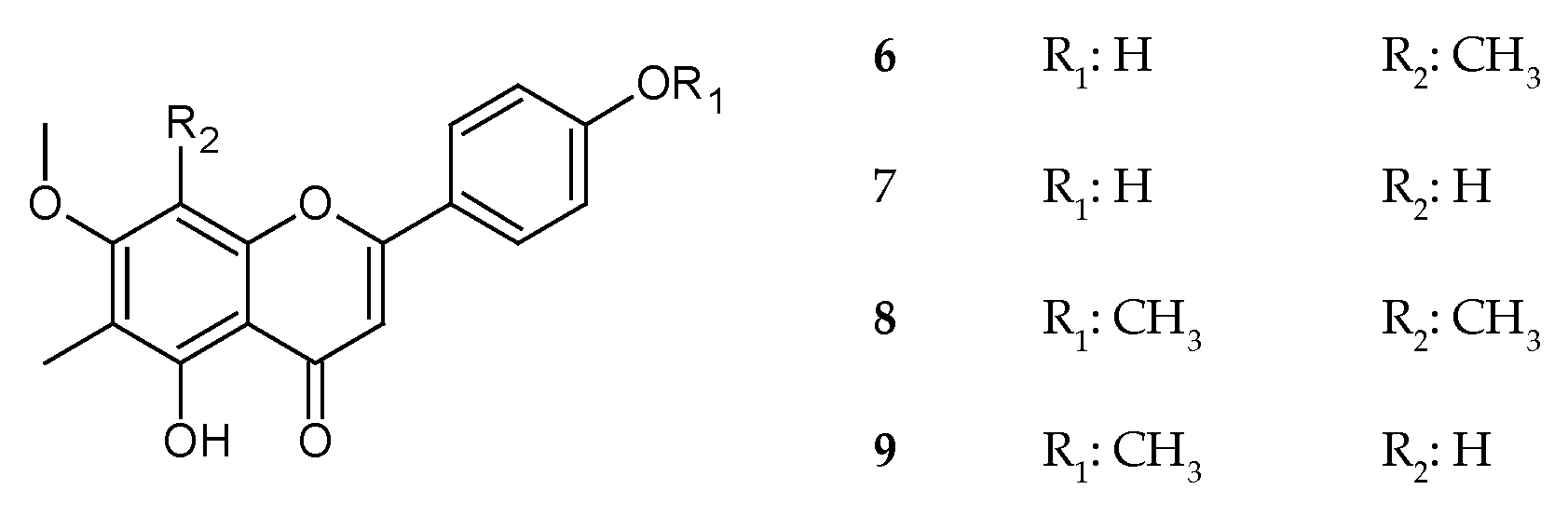
2.1.4. Isolated Compounds
2.2. Antiherpetic Activity
2.2.1. Viral Strains, Cultures, Cell Lines and Reagents
2.2.2. Determination of Cytotoxicity
2.2.3. Anti-HSV-1 Assay
2.2.4. Anti-HSV-2 Assay
2.2.5. Statistical Analysis
2.3. Antibacterial and Antifungal Activity
Microorganisms and Cultivation Media
2.4. Anti-Inflammatory Activity
2.4.1. Cell Culture
2.4.2. Cell Viability Testing
2.4.3. Detection of the Activation of NF-κB/AP-1
2.4.4. Differentiation into Macrophages and Evaluation of Cytokine Secretion
2.4.5. Detection of ROS
2.4.6. Statistical Evaluation
3. Results
3.1. Isolation and Elucidation of Compounds
3.2. Antiherpetic Activity
3.2.1. Anti-HSV-1 Activity
3.2.2. Anti-HSV-2 Activity
3.3. Antibacterial and Antifungal Activity
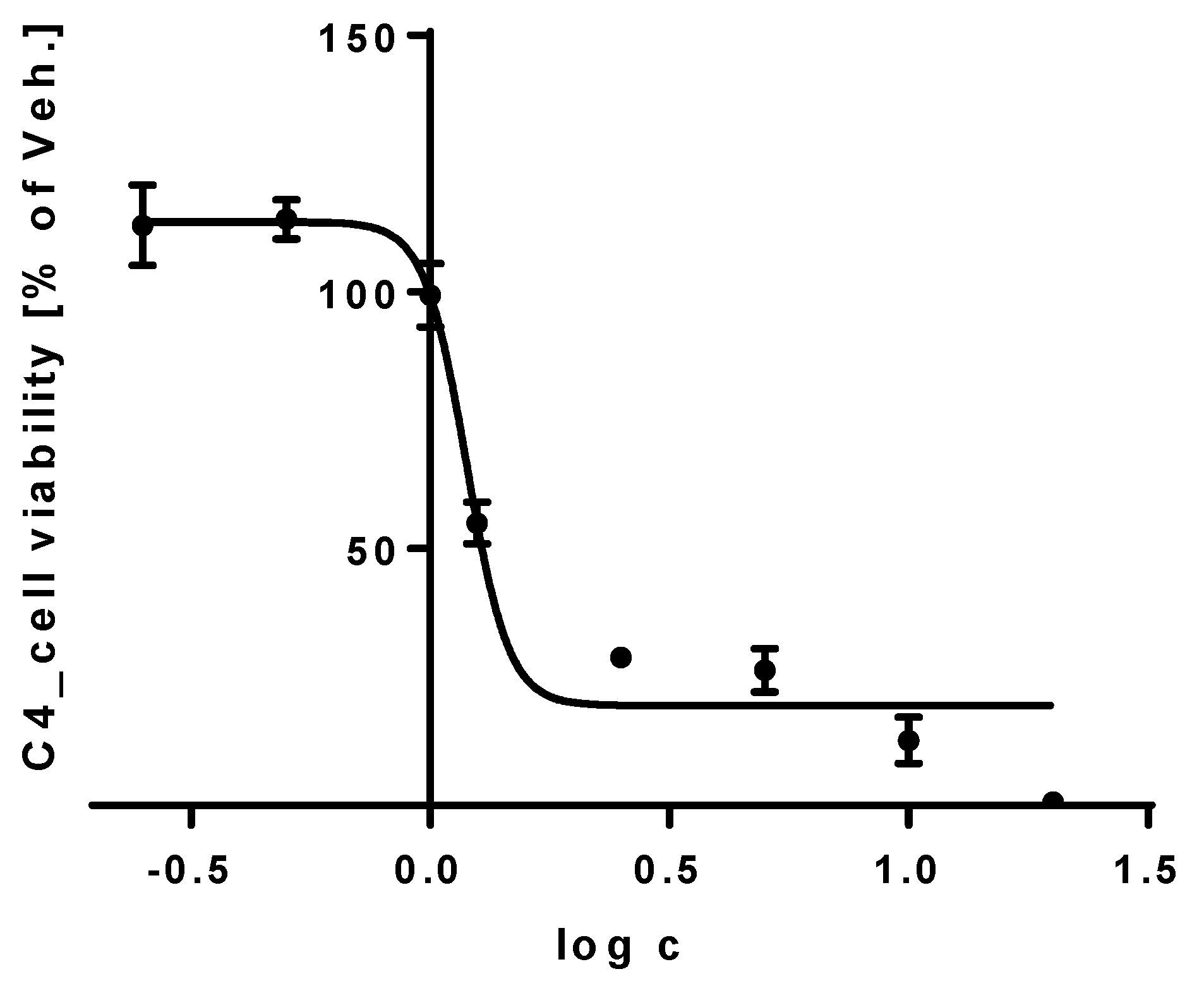
3.4. Cell Viability Assay of THP-1 Cells
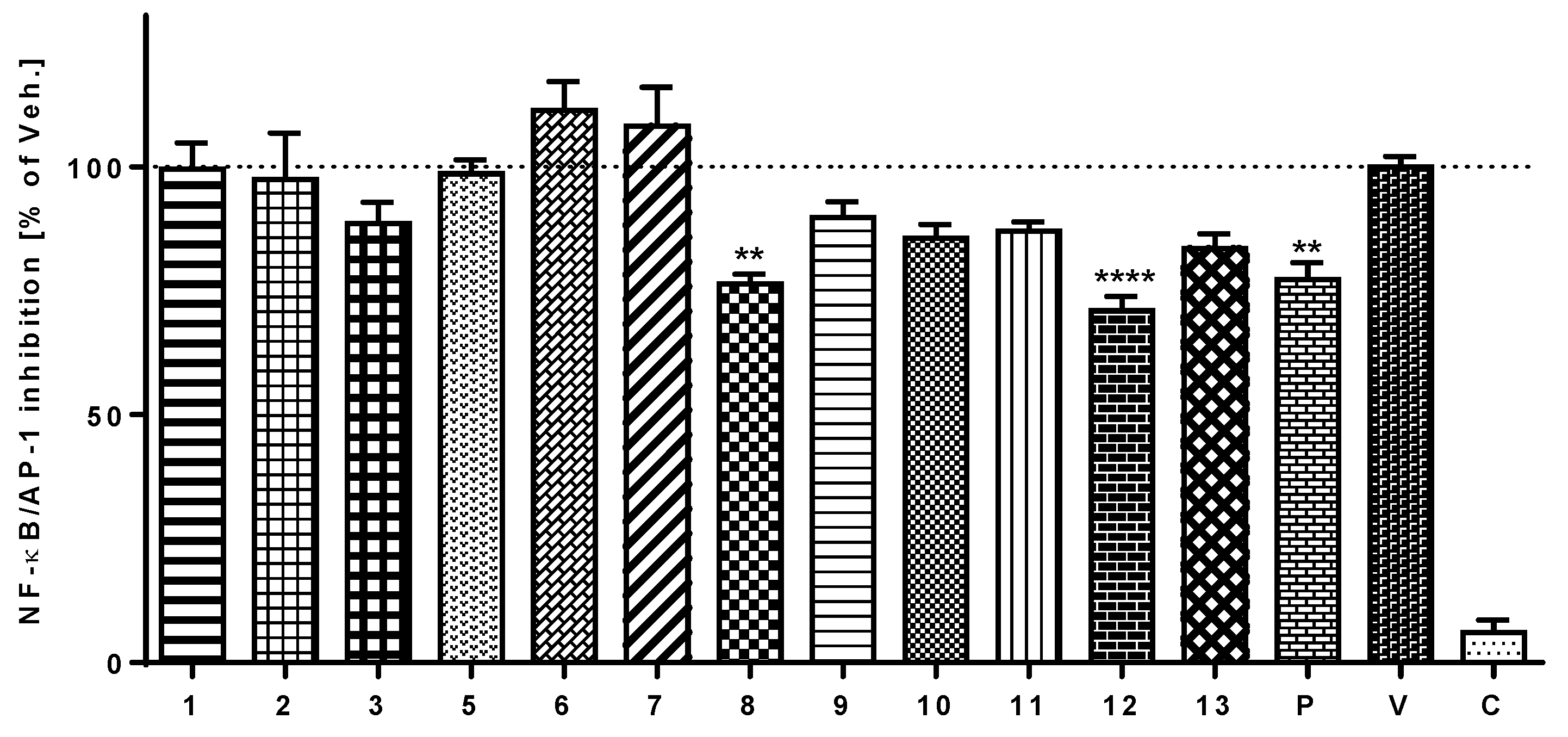
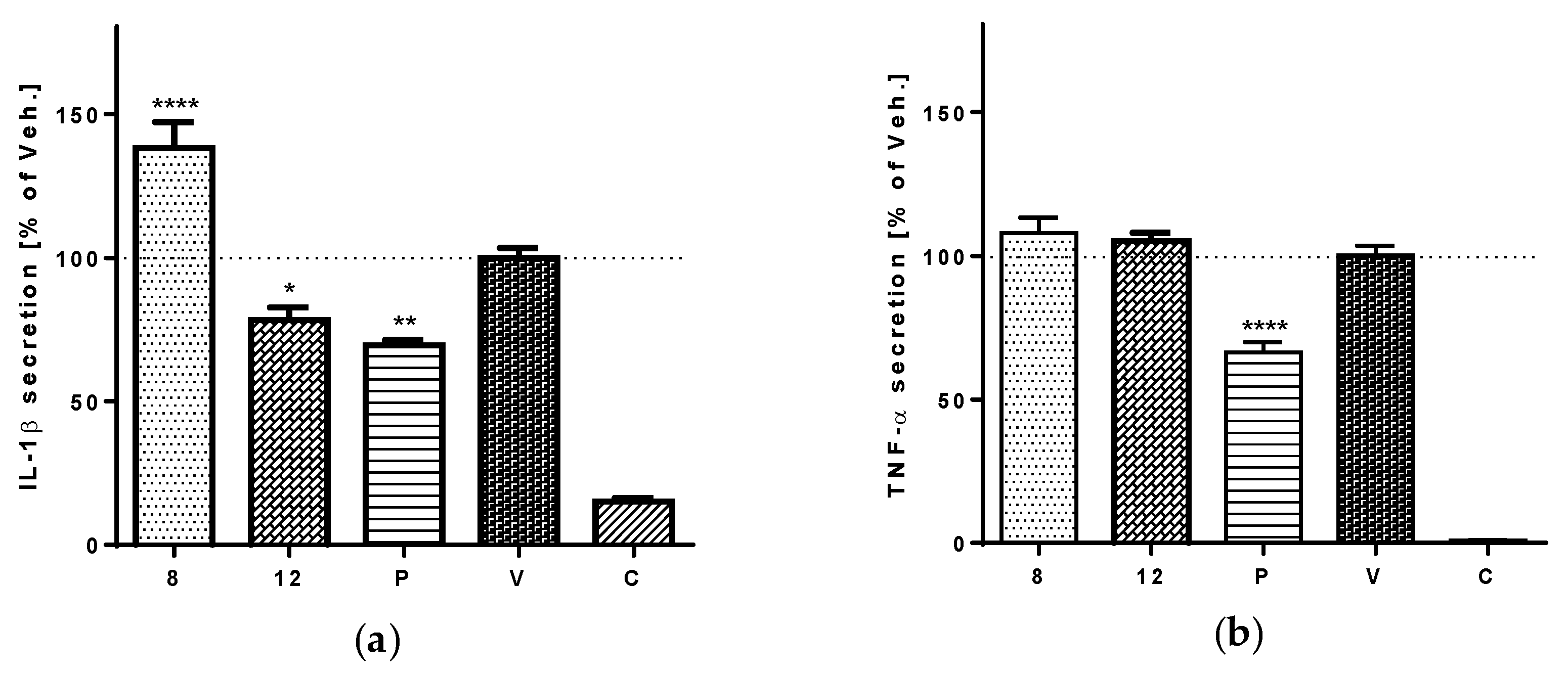
3.5. Anti-Inflammatory Activity
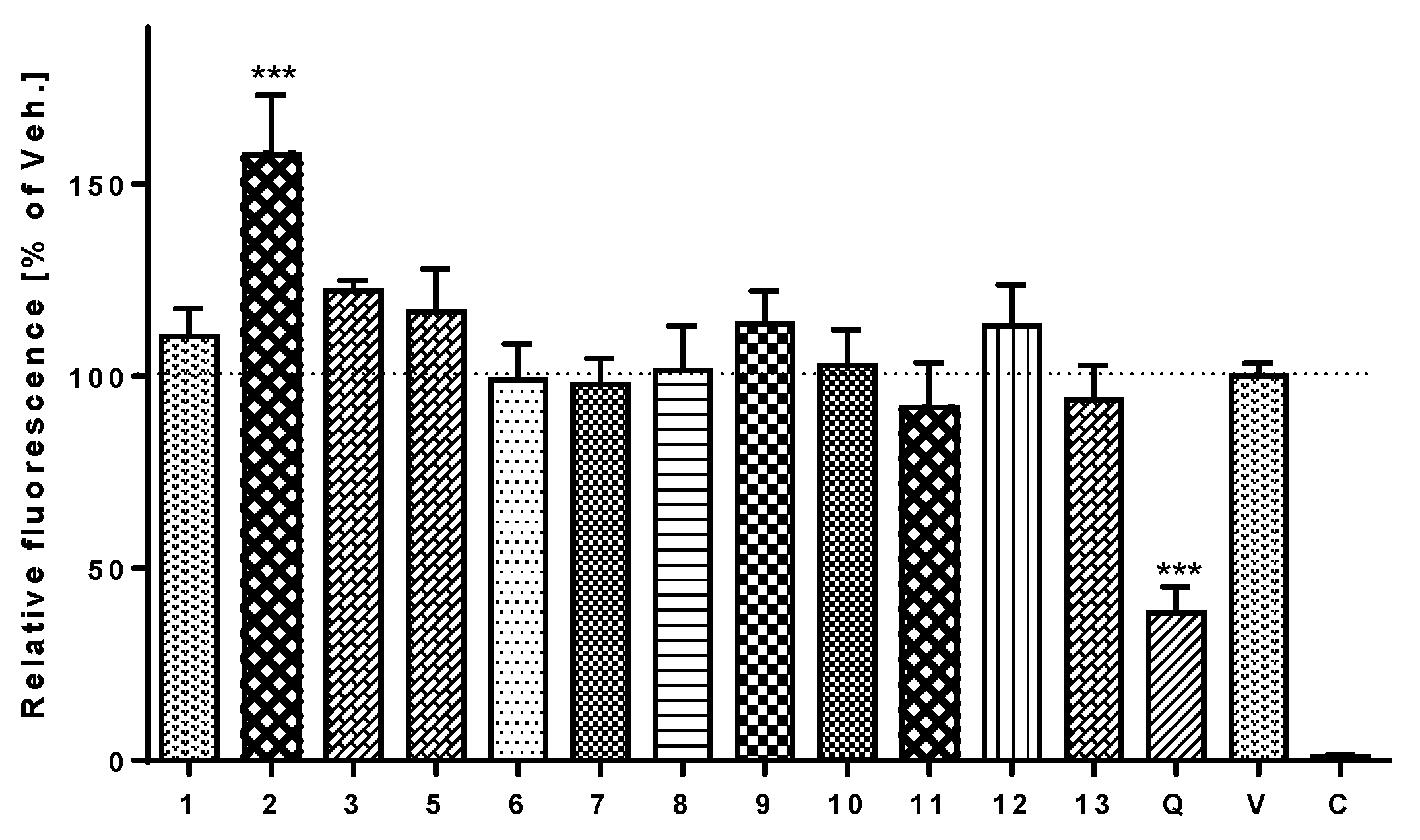
3.6. Antioxidant and Pro-Oxidant Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilles, M.; Zhao, J.; An, M.; Agboola, S. Chemical composition and antimicrobial properties of essential oils of three Australian Eucalyptus species. Food Chem. 2010, 119, 731–737. [Google Scholar] [CrossRef]
- Hardel, D.K.; Sahoo, L. A review on phytochemical and pharmacological of Eucalyptus globulus: A multipurpose tree. Int. J. Res. Ayurveda Pharm. 2011, 2, 1527–1530. [Google Scholar]
- Yao, L.; Jiang, Y.; D’Arcy, B.; Singanusong, R.; Datta, N.; Caffin, N.; Raymont, K. Quantitative high-performance liquid chromatography analyses of flavonoids in Australian Eucalyptus honeys. J. Agric. Food. Chem. 2004, 52, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Kokubo, R.; Sakaino, M. Antimicrobial activities of eucalyptus leaf extracts and flavonoids from Eucalyptus maculate. Lett. Appl. Microbiol. 2004, 39, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Ghisalberti, E.L. Bioactive acylphloroglucinol derivatives from Eucalyptus species. Phytochemistry 1996, 41, 7–22. [Google Scholar] [CrossRef]
- Takasaki, M.; Konoshima, T.; Etoh, H.; Pal Singh, I.; Tokuda, H.; Nishino, H. Cancer chemopreventive activity of euglobal-G1 from leaves of Eucalyptus grandis. Cancer Lett. 2000, 155, 61–65. [Google Scholar] [CrossRef]
- Cheng, S.S.; Huang, C.G.; Chen, Y.J.; Yu, J.J.; Chen, W.J.; Chang, S.T. Chemical compositions and larvicidal activities of leaf essential oils from two eucalyptus species. Bioresour. Technol. 2009, 100, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Lima, F.J.B.; Brito, T.S.; Freire, W.B.; Costa, R.C.; Linhares, M.I.; Sousa, F.C.; Lahlou, S.; Leal-Cardoso, J.H.; Santos, A.A.; Magalhães, P.J. The essential oil of Eucalyptus tereticornis, and its constituents α- and β-pinene, potentiate acetylcholine-induced contraction in isolated rat trachea. Fitoterapia 2010, 81, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.K.; Malik, A. Antimicrobial potential and chemical composition of Eucalyptus globulus oil in liquid and vapour phase against food spoilage microorganisms. Food Chem. 2011, 126, 228–235. [Google Scholar] [CrossRef]
- Okba, M.M.; El Gedaily, R.A.; Ashour, R.M. UPLC-PDA-ESI-qTOF-MS profiling and potent anti-HSV-II activity of Eucalyptus sideroxylon leaves. J. Chromatogr. B 2017, 1068–1069, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Abu-Jafar, A.; Huleihel, M. Antiviral activity of Eucalyptus camaldulensis leaves ethanolic extract on herpes viruses infection. Int. J. Clin. Virol. 2017, 1, 1–9. [Google Scholar] [CrossRef]
- Drew, W.L. Herpesviruses. In Sherris Medical Microbiology, 4th ed.; Ryan, K.J., Ray, C.G., Eds.; McGraw-Hill Companies, Inc.: New York, NY, USA, 2004; pp. 555–562. ISBN 10 0838585299. [Google Scholar]
- Astani, A.; Reichling, J.; Schnitzler, P. Comparative study on the antiviral activity of selected monoterpenes derived from essential oils. Phytother. Res. 2010, 24, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Minami, M.; Kita, M.; Nakaya, T.; Yamamoto, T.; Kuriyama, H.; Imanishi, J. The inhibitory effect of essential oils on herpes simplex virus type-1 replication in vitro. Microbiol. Immunol. 2003, 47, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Gavanji, S.; Sayedipour, S.S.; Larki, B.; Bakhtari, A. Antiviral activity of some plant oils against herpes simplex virus type 1 in Vero cell culture. JACME 2015, 5, 62–68. [Google Scholar] [CrossRef]
- Brezáni, V.; Šmejkal, K. Secondary metabolites isolated from the genus Eucalyptus. Curr. Top. Med. Chem. 2013, 7, 65–75. [Google Scholar]
- Burleson, F.G.; Chamberts, T.M.; Wiedbrauk, D.L. Virology: A Laboratory Manual; Academic Press: San Diego, CA, USA, 1992; p. 250. ISBN 10 0121447308. [Google Scholar]
- Hassan, S.T.S.; Berchová-Bímová, K.; Petráš, J.; Hassan, K.T.S. Cucurbitacin B interacts synergistically with antibiotics against Staphylococcus aureus clinical isolates and exhibits antiviral activity against HSV-1. S. Afr. J. Bot. 2017, 108, 90–94. [Google Scholar] [CrossRef]
- Hassan, S.T.S.; Švajdlenka, E.; Berchová-Bímová, K. Hibiscus sabdariffa L. and its bioactive constituents exhibit antiviral activity against HSV-2 and anti-enzymatic properties against urease by an ESI-MS based assay. Molecules 2017, 22, 722. [Google Scholar] [CrossRef] [PubMed]
- Borenfreund, E.; Puerner, J.A. Toxicity determined in vitro by morphological alterations and neutral red absorption. Toxicol. Lett. 1985, 24, 119–124. [Google Scholar] [CrossRef]
- Silva, I.T.; Costa, G.M.; Stoco, P.H.; Schenkel, E.P.; Reginatto, F.H.; Simões, C.M.O. In vitro antiherpes effects of C-glycosyl flavonoid-enriched fraction of Cecropia glazivoii Sneth. Lett. Appl. Microbiol. 2010, 51, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Tan, G.T.; Santarsiero, B.D.; Mesecar, A.D.; Hung, N.V.; Cuong, N.M.; Doel Soejarto, D.; Pezzuto, J.M.; Fong, H.H. New sesquiterpenes from Litsea verticillata. J. Nat. Prod. 2003, 66, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Asami, T.; Kawano, T.; Yoneyama, K.; Crow, W.D.; Paton, D.M.; Takahashi, N. Photosynthetic inhibitors in Eucalyptus grandis. Phytochemistry 1988, 27, 1943–1946. [Google Scholar] [CrossRef]
- Wang, W.; Zeng, Y.H.; Osman, K.; Shinde, K.; Rahman, M.; Gibbons, S.; Mu, Q. Norlignans, acylphloroglucinols, and a dimeric xanthone from Hypericum chinense. J. Nat. Prod. 2010, 73, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Xue, J.J.; Fan, C.Q.; Miao, Z.H.; Ding, J.; Yue, J.M. Eucalyptals A–C with a new skeleton isolated from Eucalyptus globulus. Org. Lett. 2007, 9, 5549–5552. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Koreishi, M.; Tokuda, H.; Nishino, H.; Yoshida, T. Cypellocarpins A–C, phenol glycosides esterified with oleuropeic acid, from Eucalyptus cypellocarpa. J. Nat. Prod. 2000, 63, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Hakki, Z.; Cao, B.; Heskes, A.M.; Goodger, J.Q.; Woodrow, I.E.; Williams, S.J. Synthesis of the monoterpenoid esters cypellocarpin C and cuniloside B and evidence for their widespread occurrence in Eucalyptus. Carbohydr. Res. 2010, 345, 2079–2084. [Google Scholar] [CrossRef] [PubMed]
- Junio, H.A.; Sy-Cordero, A.A.; Ettefagh, K.A.; Burns, J.T.; Micko, K.T.; Graf, T.N.; Richter, S.J.; Cannon, R.E.; Oberlies, N.H.; Cech, N.B. Synergy-directed fractionation of botanical medicines: A case study with Goldenseal (Hydrastis canadensis). J. Nat. Prod. 2011, 74, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Pagola, S.; Tracanna, M.I.; Amani, S.M.; Gonzáles, A.M.; Raschi, A.B.; Romano, E.; Benavente, A.M.; Stephens, P.W. Sideroxylin from Miconia ioneura: Monohydrate crystal structure from high resolution X-ray powder diffraction. Nat. Prod. Commun. 2008, 3, 759–764. [Google Scholar]
- Wollenweber, E.; Kohortst, G. Epicuticular leaf flavonoids from Eucalyptus species and from Kalmia latifolia. Z. Naturforsch. C Biosci. C 1981, 36, 913–915. [Google Scholar]
- Huq, F.; Misra, L.N. An alkenol and C-methylated flavones from Callistemon lanceolatus leaves. Planta Med. 1997, 63, 369–370. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.I.; Freire, C.S.; Pascoal Neto, C.; Silvestre, A.J.; Silva, A.M. Chemical composition of the epicuticular wax from the fruits of Eucalyptus globulus. Phytochem. Anal. 2005, 16, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fujimoto, Y. Triterpene esters from Eucalyptus tereticornis. Phytochemistry 1993, 33, 151–153. [Google Scholar] [CrossRef]
- Sidana, J.; Singh, S.; Arora, S.K.; Foley, W.J.; Singh, I.P. Terpenoidal constituents of Eucalyptus loxophleba ssp. Lissophloia. Pharm. Biol. 2012, 50, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.M.; Yang, X.W. Cypellocarpin C and other compounds from the fruits of Eucalyptus globulus Labill. Biochem. Syst. Ecol. 2006, 34, 543–545. [Google Scholar] [CrossRef]
- Heidary Navid, M.; Laszczyk-Lauer, M.N.; Reichling, J.; Schnitzler, P. Pentacyclic triterpenes in birch bark extract inhibit early step of herpes simplex virus type 1 replication. Phytomedicine 2014, 21, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.; Ha, T.K.Q.; Kang, K.B.; Kim, K.H.; Oh, W.K.; Sung, S.H. C-methylated flavonoid glycosides from Pentarhizidium orientale rhizomes and their inhibitory effects on the H1N1 influenza virus. J. Nat. Prod. 2017, 80, 2818–2824. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, R.; Murata, M.; Homma, S.; Aida, K. Antibacterial compounds from Eucalyptus perriniana. Agric. Biol. Chem. 1990, 54, 231–232. [Google Scholar] [CrossRef]
- Khan, I.; Bahuguna, A.; Kumar, P.; Bajpai, V.K.; Kang, S.C. Antimicrobial potential of carvacrol against uropathogenic Escherichia coli via membrane disruption, depolarization, and reactive oxygen species generation. Front. Microbiol. 2017, 8, 2421. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [PubMed]
- Bolte, M.L.; Crow, W.D.; Tahahashi, N.; Sakurai, A.; Uji-Ie, M.; Yoshida, S. Structure/activity relationship of grandinol: A germination inhibitor in Eucalyptus. Agric. Biol. Chem. 1985, 49, 761–768. [Google Scholar] [CrossRef]
- Ibewuike, J.C.; Ogungbamila, F.O.; Ogundaini, A.O.; Okeke, I.N.; Bohlin, L. Antiinflammatory and antibacterial activities of C-methylflavonols from Pilostigma thonningii. Phytother. Res. 1997, 11, 281–284. [Google Scholar] [CrossRef]
- Brasier, A.R. The NF-κB regulatory network. Cardiovasc. Toxicol. 2006, 6, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.-K.; Mitchell, P.J.; Brown, N.; Drummond, R.A.; Brown, G.D.; Kaye, P.M.; Bowles, D.J. Regiospecific Methylation of a Dietary Flavonoid Scaffold Selectively Enhances IL-1β Production Following Toll-like Receptor 2 Stimulation in THP-1 Monocytes. J. Biol. Chem. 2013, 288, 21126–21135. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | CC50 (μg/mL) | IC50 (μg/mL) | Selectivity Index CC50/IC50 |
|---|---|---|---|
| 2 | >210 | 1.23 ± 0.13 | >170.7 |
| 6 | >210 | 1.44 ± 0.14 | >145.8 |
| 12 | >210 | 0.96 ± 0.12 | >218.8 |
| Acyclovir | >210 | 1.92 ± 0.23 | >109.4 |
| Compound | CC50 (μg/mL) | EC50 (μg/mL) | Selectivity Index CC50/EC50 |
|---|---|---|---|
| 1 | >210 | 1.25 ± 0.20 | >168.0 |
| 5 | >210 | 0.73 ± 0.11 | >287.7 |
| Acyclovir | >210 | 1.75 ± 0.33 | >120.0 |
| Compound | S. aureus ATCC 29213 | B. cereus ATCC 11778 | E. coli ATCC 25922 | P. aeruginosa ATCC 27853 | C. albicans ATCC 10231 |
|---|---|---|---|---|---|
| 1 | >64 | 64 | >64 | >64 | >64 |
| 2 | 16 | 16 | >64 | >64 | >64 |
| 3 | 32 | 8 | >32 | >32 | >32 |
| 4 | >128 | 32 | >128 | >128 | >128 |
| 5 | >128 | >128 | >128 | >128 | >128 |
| 6 | >64 | 64 | >64 | >64 | >64 |
| 7 | >128 | 128 | >128 | >128 | >128 |
| 8 | >64 | >64 | >64 | >64 | >64 |
| 9 | >64 | >64 | >64 | >64 | >64 |
| 10 | >32 | >32 | >32 | >32 | >32 |
| 11 | 128 | 32 | >128 | >128 | >128 |
| 12 | >32 | >32 | >32 | >32 | >32 |
| 13 (a + b) | 128 | 16 | 128 | 128 | 128 |
| Antibiotic | 0.125 a | 0.03 a | 1 a | 16 a | 4 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brezáni, V.; Leláková, V.; Hassan, S.T.S.; Berchová-Bímová, K.; Nový, P.; Klouček, P.; Maršík, P.; Dall’Acqua, S.; Hošek, J.; Šmejkal, K. Anti-Infectivity against Herpes Simplex Virus and Selected Microbes and Anti-Inflammatory Activities of Compounds Isolated from Eucalyptus globulus Labill. Viruses 2018, 10, 360. https://doi.org/10.3390/v10070360
Brezáni V, Leláková V, Hassan STS, Berchová-Bímová K, Nový P, Klouček P, Maršík P, Dall’Acqua S, Hošek J, Šmejkal K. Anti-Infectivity against Herpes Simplex Virus and Selected Microbes and Anti-Inflammatory Activities of Compounds Isolated from Eucalyptus globulus Labill. Viruses. 2018; 10(7):360. https://doi.org/10.3390/v10070360
Chicago/Turabian StyleBrezáni, Viliam, Veronika Leláková, Sherif T. S. Hassan, Kateřina Berchová-Bímová, Pavel Nový, Pavel Klouček, Petr Maršík, Stefano Dall’Acqua, Jan Hošek, and Karel Šmejkal. 2018. "Anti-Infectivity against Herpes Simplex Virus and Selected Microbes and Anti-Inflammatory Activities of Compounds Isolated from Eucalyptus globulus Labill." Viruses 10, no. 7: 360. https://doi.org/10.3390/v10070360
APA StyleBrezáni, V., Leláková, V., Hassan, S. T. S., Berchová-Bímová, K., Nový, P., Klouček, P., Maršík, P., Dall’Acqua, S., Hošek, J., & Šmejkal, K. (2018). Anti-Infectivity against Herpes Simplex Virus and Selected Microbes and Anti-Inflammatory Activities of Compounds Isolated from Eucalyptus globulus Labill. Viruses, 10(7), 360. https://doi.org/10.3390/v10070360