Bacteriophages of Myxococcus xanthus, a Social Bacterium


Abstract
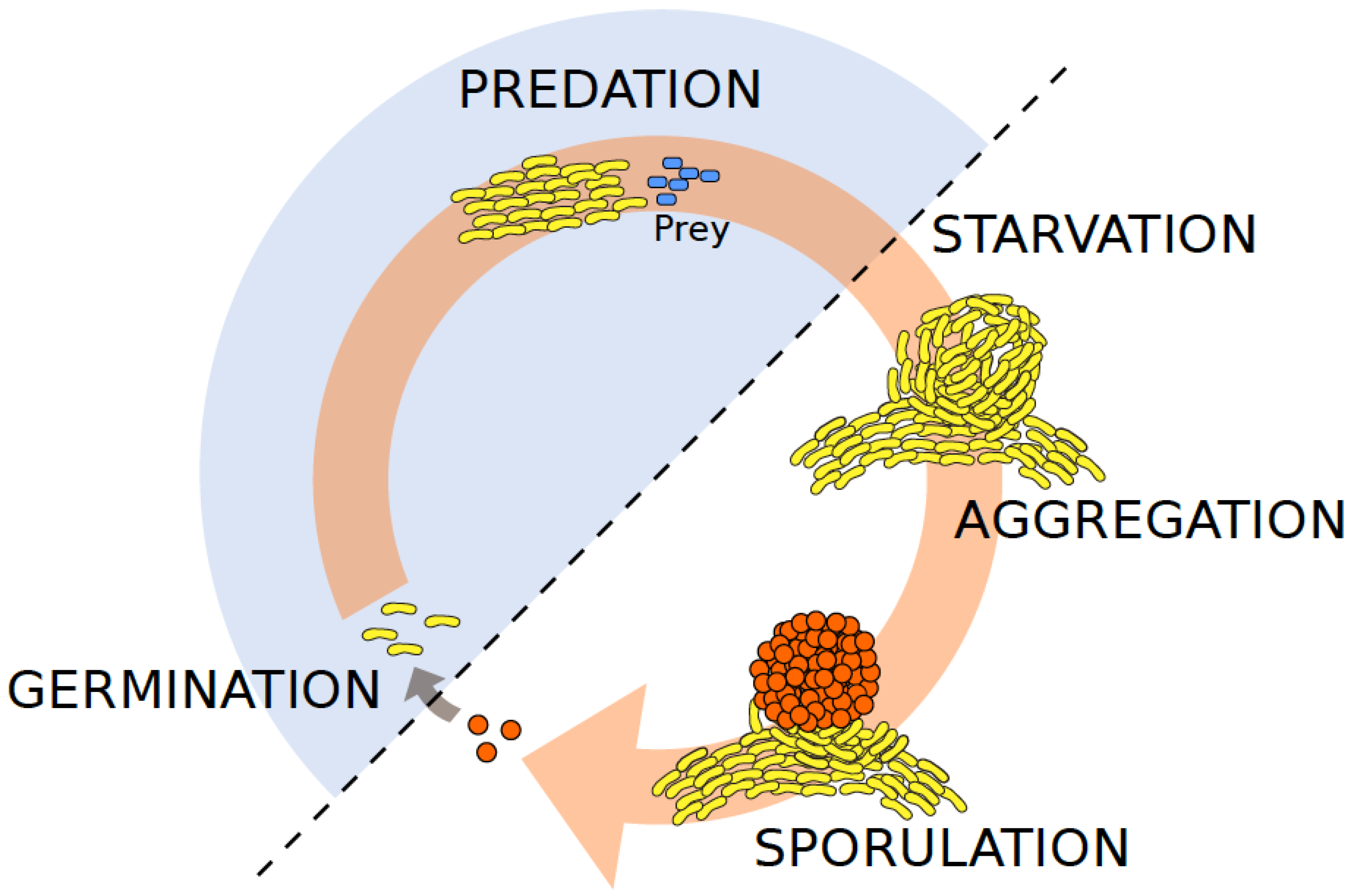
:1. Introduction
2. Isolation Procedures of Myxophages from Nature
3. Nomenclature of Myxophages
- (1)
- A phage found to infect a certain myxobacterial species is assigned letters composed of this initial host’s genus and species, respectively; e.g., phages found to infect M. xanthus are assigned Mx, and those of M. virescens should be named Mv. In some cases, ambiguity will arise due to identical initials. In such cases more letters should be used, e.g., Cfu and Cfe for Cystobacter fuscus and Cystobacter ferrugineus, respectively. If multiple host species are known, the myxophage name may refer to the original source host (if applicable), or arbitrarily to either of the host species if directly derived from environmental samples (such as soil).
- (2)
- The letters are immediately followed by a distinct serotype category expressed as a running number. This will both help to account for existing phage names and also allow for an unlimited number of novel categories. As before, the first phage describing a novel serotype group is called the prototype-phage and is adopted as representative of that novel serogroup.
- (3)
- For non-prototype phages, the name is finally capped off by a running number separated by a dash from the main group in order of first description, e.g., novel phage isolates that are serologically classified as “Mx1-like” are labeled Mx1-1, Mx1-2, etc.
4. Characterization of the Main Groups of Myxophages
4.1. The Mx1-Like Group
4.2. The Mx4-Like Group
4.3. The Mx8-Like Group
4.4. The Mx9-Like Group
4.5. Prophage Mx Alpha
4.6. Misclassified Phages
4.7. Phage Remnants and Bacteriocins
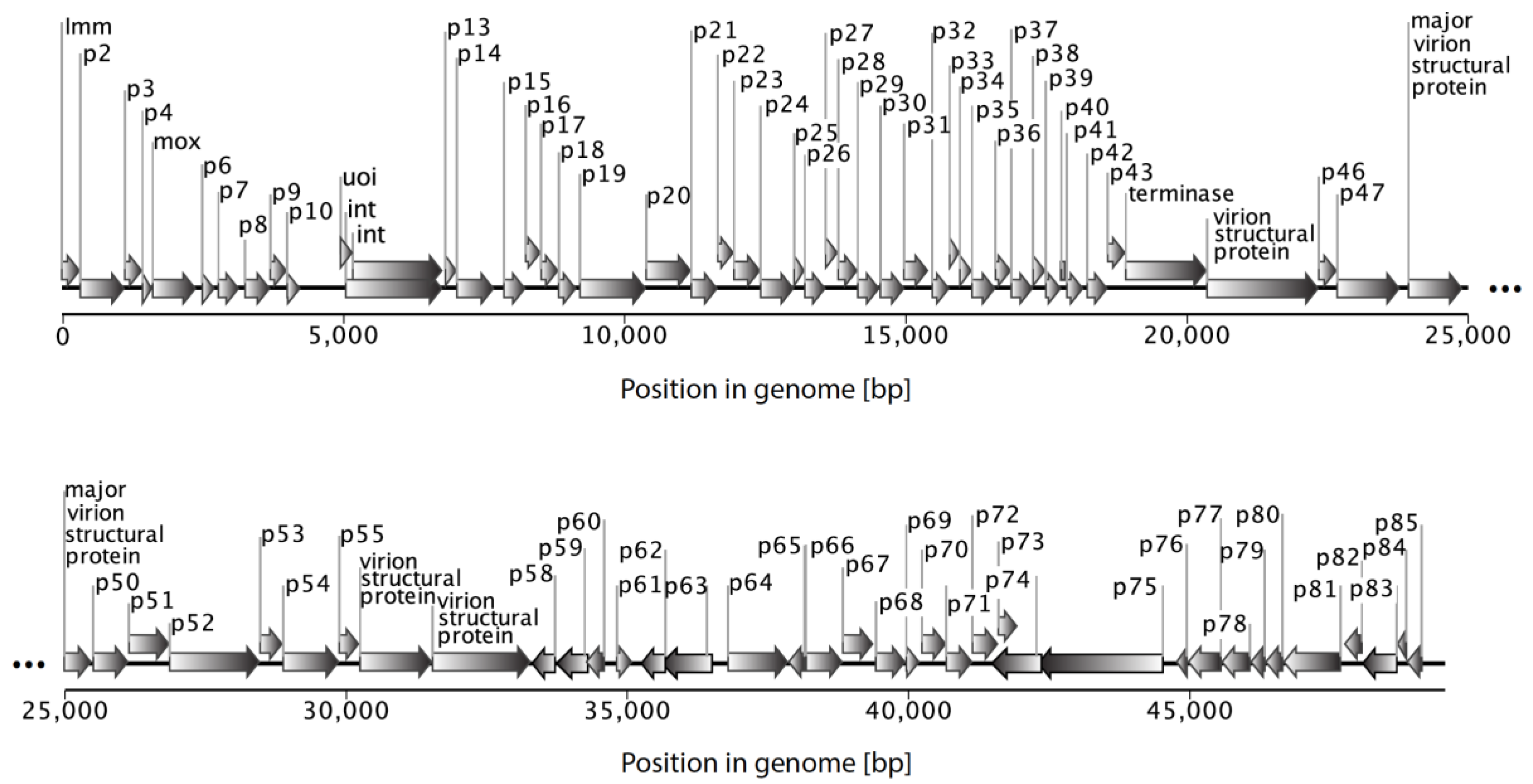
5. Genomic Features of the Main Myxophage Groups
6. Concluding Remarks and Future Research Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colombo, S.; Arioli, S.; Neri, E.; Della Scala, G.; Gargari, G.; Mora, D. Viromes as genetic reservoir for the microbial communities in aquatic environments: A focus on antimicrobial-resistance genes. Front. Microbiol. 2017, 8, 1095. [Google Scholar] [CrossRef] [PubMed]
- Roux, S.; Hallam, S.J.; Woyke, T.; Sullivan, M.B. Viral dark matter and virus-host interactions resolved from publicly available microbial genomes. eLife 2015, 4, e08490. [Google Scholar] [CrossRef] [PubMed]
- Scarpellini, E.; Ianiro, G.; Attili, F.; Bassanelli, C.; de Santis, A.; Gasbarrini, A. The human gut microbiota and virome: Potential therapeutic implications. Dig. Liver Dis. 2015, 47, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Wolf, Y.I. Evolutionary genomics of defense systems in Archaea and Bacteria. Annu. Rev. Microbiol. 2017, 71, 233–261. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Phage evolution and ecology. Adv. Appl. Microbiol. 2009, 67, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Ofir, G.; Sorek, R. Contemporary phage biology: From classic models to new insights. Cell 2018, 172, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Editorial. Microbiology by numbers. Nat. Rev. Microbiol. 2011, 9, 628. [Google Scholar]
- Erez, Z.; Steinberger-Levy, I.; Shamir, M.; Doron, S.; Stokar-Avihail, A.; Peleg, Y.; Melamed, S.; Leavitt, A.; Savidor, A.; Albeck, S.; et al. Communication between viruses guides lysis-lysogeny decisions. Nature 2017, 541, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Broussard, G.W.; Oldfield, L.M.; Villanueva, V.M.; Lunt, B.L.; Shine, E.E.; Hatfull, G.F. Integration-dependent bacteriophage immunity provides insights into the evolution of genetic switches. Mol. Cell 2013, 49, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.M.; Wu, Z.H.; Zhao, J.Y.; Li, Y.Z. Fruiting and non-fruiting myxobacteria: A phylogenetic perspective of cultured and uncultured members of this group. Mol. Phylogenet. Evol. 2007, 44, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Shimkets, L.; Woese, C.R. A phylogenetic analysis of the myxobacteria: Basis for their classification. Proc. Natl. Acad. Sci. USA 1992, 89, 9459–9463. [Google Scholar] [CrossRef] [PubMed]
- Dawid, W. Biology and global distribution of myxobacteria in soils. FEMS Microbiol. Rev. 2000, 24, 403–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkhoff, T.; Fischer, D.; Vollmers, J.; Voget, S.; Beardsley, C.; Thole, S.; Mussmann, M.; Kunze, B.; Wagner-Dobler, I.; Daniel, R.; et al. Biogeography and phylogenetic diversity of a cluster of exclusively marine myxobacteria. ISME J. 2012, 6, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z.; Hu, W.; Zhang, Y.Q.; Qiu, Z.; Zhang, Y.; Wu, B.H. A simple method to isolate salt-tolerant myxobacteria from marine samples. J. Microbiol. Methods 2002, 50, 205–209. [Google Scholar] [CrossRef]
- Keane, R.; Berleman, J. The predatory life cycle of Myxococcus xanthus. Microbiology 2016, 162, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Geitler, L. Über Polyangium parasiticum n. sp. eine submerse, parasitische Myxobacteriacee. Archiv für Protistenkunde 1925, 50, 67–88. [Google Scholar]
- Zhou, X.W.; Li, S.G.; Li, W.; Jiang, D.M.; Han, K.; Wu, Z.H.; Li, Y.Z. Myxobacterial community is a predominant and highly diverse bacterial group in soil niches. Environ. Microbiol. Rep. 2014, 6, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-W.; Rettberg, L.A.; Treuner-Lange, A.; Iwasa, J.; Søgaard-Andersen, L.; Jensen, G.J. Architecture of the type IVa pilus machine. Science 2016, 351, aad2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faure, L.M.; Fiche, J.B.; Espinosa, L.; Ducret, A.; Anantharaman, V.; Luciano, J.; Lhospice, S.; Islam, S.T.; Treguier, J.; Sotes, M.; et al. The mechanism of force transmission at bacterial focal adhesion complexes. Nature 2016, 539, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Velicer, G.J.; Vos, M. Sociobiology of the myxobacteria. Annu. Rev. Microbiol. 2009, 63, 599–623. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E. Preface. In Myxobacteria: Development and Cell Interactions, 1st ed.; Rosenberg, E., Ed.; Springer: New York, NY, USA, 1984; pp. vii–viii. [Google Scholar]
- Fiegna, F.; Yu, Y.T.; Kadam, S.V.; Velicer, G.J. Evolution of an obligate social cheater to a superior cooperator. Nature 2006, 441, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Velicer, G.J.; Kroos, L.; Lenski, R.E. Loss of social behaviors by Myxococcus xanthus during evolution in an unstructured habitat. Proc. Natl. Acad. Sci. USA 1998, 95, 12376–12380. [Google Scholar] [CrossRef] [PubMed]
- Velicer, G.J.; Kroos, L.; Lenski, R.E. Developmental cheating in the social bacterium Myxococcus xanthus. Nature 2000, 404, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Velicer, G.J.; Yu, Y.-T.N. Evolution of novel cooperative swarming in the bacterium Myxococcus xanthus. Nature 2003, 425, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.T.; Yuan, X.; Velicer, G.J. Adaptive evolution of an sRNA that controls Myxococcus development. Science 2010, 328, 993. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, S.A.; Toups, M.A.; Velicer, G.J. Natural variation in developmental life-history traits of the bacterium Myxococcus xanthus. FEMS Microbiol. Ecol. 2010, 73, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, S.A.; Velicer, G.J. Social complementation and growth advantages promote socially defective bacterial isolates. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140036. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.; Velicer, G.J. Natural variation of gliding motility in a centimetre-scale population of Myxococcus xanthus. FEMS Microbiol. Ecol. 2008, 64, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, S.A.; Velicer, G.J. Endemic social diversity within natural kin groups of a cooperative bacterium. Proc. Natl. Acad. Sci. USA 2011, 108, 10823–10830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, M.; Velicer, G.J. Genetic population structure of the soil bacterium Myxococcus xanthus at the centimeter scale. Appl. Environ. Microbiol. 2006, 72, 3615–3625. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.; Velicer, G.J. Isolation by distance in the spore-forming soil bacterium Myxococcus xanthus. Curr. Biol. 2008, 18, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Wielgoss, S.; Didelot, X.; Chaudhuri, R.R.; Liu, X.; Weedall, G.D.; Velicer, G.J.; Vos, M. A barrier to homologous recombination between sympatric strains of the cooperative soil bacterium Myxococcus xanthus. ISME J. 2016, 10, 2468–2477. [Google Scholar] [CrossRef] [PubMed]
- Burchard, R.P.; Dworkin, M. A bacteriophage for Myxococcus xanthus: Isolation, characterization and relation of infectivity to host morphogenesis. J. Bacteriol. 1966, 91, 1305–1313. [Google Scholar] [PubMed]
- Murphy, K.A.; Garza, A.G. Genetic tools for studying Myxococcus xanthus biology. In Myxobacteria: Multicellularity and Differentiation, 3rd ed.; Whitworth, D.E., Ed.; ASM Press: Washington, DC, USA, 2008; pp. 491–501. [Google Scholar]
- Hodgkin, J.; Kaiser, D. Genetics of gliding motility in Myxococcus xanthus (Myxobacterales): Two gene systems control movement. Mol. Gen. Genet. 1979, 171, 177–191. [Google Scholar] [CrossRef]
- Wall, D.; Kolenbrander, P.E.; Kaiser, D. The Myxococcus xanthus pilQ (sglA) gene encodes a secretin homolog required for Type IV pilus biogenesis, social motility and development. J. Bacteriol. 1999, 181, 24–33. [Google Scholar] [PubMed]
- Hartzell, P.L. Genetic tools for dissecting motility and development of Myxococcus xanthus. In Modern Microbial Genetics, 2nd ed.; Streips, U.N., Yasbin, R.E., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 2002; pp. 289–322. [Google Scholar]
- Martin, S.; Sodergren, E.; Masuda, T.; Kaiser, D. Isolation of transducing phages for Myxococcus. Virology 1978, 53, 44–53. [Google Scholar] [CrossRef]
- Singh, B.N. Myxobacteria in soils and composts; their distribution, number and lytic action on bacteria. J. Gen. Microbiol. 1947, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.L.; Burchard, R.P.; Morris, D.W.; Parish, J.H.; Stow, N.D.; Tsopanakis, C. Phage and defective phage of strains of Myxococcus. Arch. Microbiol. 1976, 108, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.K.; Virrankoski-Castrodeza, V.; Parish, J.H.; Grimm, K. Isolation and characterization of new bacteriophages for Myxococcus xanthus. Arch. Microbiol. 1980, 180, 175–180. [Google Scholar] [CrossRef]
- Tsopanakis, C.; Parish, J.H. Bacteriophage MX-1: Properties of the phage and its structural proteins. J. Gen. Virol. 1976, 30, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Paez-Espino, D.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Thomas, A.D.; Huntemann, M.; Mikhailova, N.; Rubin, E.; Ivanova, N.N.; Kyrpides, N.C. Uncovering Earth’s virome. Nature 2016, 536, 425–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, J.M.; Geisselsoder, J.; Zusman, D.R. Isolation of bacteriophage MX4, a generalized transducing phage for Myxococcus xanthus. J. Mol. Biol. 1978, 119, 167–178. [Google Scholar] [CrossRef]
- Orndorff, P.; Stellwag, E.; Starich, T.; Dworkin, M.; Zissler, J. Genetic and physical characterization of lysogeny by bacteriophage MX8 in Myxococcus xanthus. J. Bacteriol. 1983, 154, 772–779. [Google Scholar] [PubMed]
- Geisselsoder, J.; Campos, J.M.; Zusman, D.R. Physical characterization of bacteriophage MX4, a generalized transducing phage for Myxococcus xanthus. J. Mol. Biol. 1978, 119, 179–189. [Google Scholar] [CrossRef]
- Ackermann, H.-W.; Abedon, S.T. Bacteriophage Names 2000. The Bacteriophage Ecology Group. Available online: http://www.phage.org/names/2000/ (accessed on 18 May 2018).
- Azuaga, M.J.; Muñoz, J.; Gonzáles, F.; Arias, J.M. Isolation and characterization of bacteriophages from Myxococcus virescens. Microbios 1990, 61, 83–88. [Google Scholar]
- Voelz, H.; Burchard, R.P. Fine structure of bacteriophage-infected Myxococcus xanthus. I. The lytic cycle in vegetative cells. Virology 1971, 43, 243–250. [Google Scholar] [CrossRef]
- Salmi, D.; Magrini, V.; Hartzell, P.L.; Youderian, P. Genetic determinants of immunity and integration of temperate Myxococcus xanthus phage Mx8. J. Bacteriol. 1998, 180, 614–621. [Google Scholar] [PubMed]
- Julien, B. Characterization of the integrase gene and attachment site for the Myxococcus xanthus bacteriophage Mx9. J. Bacteriol. 2003, 185, 6325–6330. [Google Scholar] [CrossRef] [PubMed]
- Vasse, M.; Wielgoss, S. Public database of Myxophages. Github. Avaliable online: https://github.com/marievasse/Myxophages (accessed on 17 July 2018).
- Burchard, R.P.; Voelz, H. Bacteriophage infection of Myxococcus xanthus during cellular differentiation and vegetative growth. Virology 1972, 48, 555–566. [Google Scholar] [CrossRef]
- Kaiser, D. Genetic systems in bacteria. Methods Enzymol. 1991, 204, 357–372. [Google Scholar] [PubMed]
- Starich, T.; Cordes, P.; Zissler, J. Transposon tagging to detect a latent virus in Myxococcus xanthus. Science 1985, 230, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Starich, T.; Zissler, J. Movement of multiple DNA units between Myxococcus xanthus cells. J. Bacteriol. 1989, 171, 2323–2336. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Vassallo, C.N.; Conklin, A.C.; Pathak, D.T.; Troselj, V.; Wall, D. Sibling rivalry in Myxococcus xanthus is mediated by kin recognition and a polyploid prophage. J. Bacteriol. 2016, 198, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Keseler, I.M.; Shimkets, L.J. Genome size of Myxococcus xanthus determined by pulsed-field gel electrophoresis. J. Bacteriol. 1990, 172, 4206–4213. [Google Scholar] [CrossRef] [PubMed]
- Goldman, B.S.; Nierman, W.C.; Kaiser, D.; Slater, S.C.; Durkin, A.S.; Eisen, J.A.; Ronning, C.M.; Barbazuk, W.B.; Blanchard, M.; Field, C.; et al. Evolution of sensory complexity recorded in a myxobacterial genome. Proc. Natl. Acad. Sci. USA 2006, 103, 15200–15205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassallo, C.N.; Cao, P.; Conklin, A.; Finkelstein, H.; Hayes, C.S.; Wall, D. Infectious polymorphic toxins delivered by outer membrane exchange discriminate kin in myxobacteria. eLife 2017, 6, e29397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anacker, R.L.; Ordal, E.J. Study of a bacteriophage infecting the myxobacterium Chondrococcus columnaris. J. Bacteriol. 1955, 70, 738–741. [Google Scholar] [PubMed]
- Kingsbury, D.T.; Ordal, E.J. Bacteriophage infecting the myxobacterium Chondrococcus columnaris. J. Bacteriol. 1966, 91, 1327–1332. [Google Scholar] [PubMed]
- Declercq, A.M.; Haesebrouck, F.; van den Broeck, W.; Bossier, P.; Decostere, A. Columnaris disease in fish: A review with emphasis on bacterium-host interactions. Vet. Res. 2013, 44, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordal, E.J.; Rucker, R.R. Pathogenic myxobacteria. Proc. Soc. Exp. Biol. Med. 1944, 56, 15–18. [Google Scholar] [CrossRef]
- McCurdy, H.D., Jr.; MacRae, T.H. Xanthacin. A bacteriocin of Myxococcus xanthus fb. Can. J. Microbiol. 1974, 20, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Reichenbach, H. Die wahre Natur der Myxobakterien—“Rhapidosomen”. Arch. Mikrobiol. 1967, 56, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.L.; Morris, D.W.; Parish, J.H. DNA of Myxococcus bacteriophage MX-1: Macromolecular properties and restriction fragments. Arch. Microbiol. 1976, 226, 221–226. [Google Scholar] [CrossRef]
- Morris, D.W.; Brown, N.L.; Parish, J.H. DNA of Myxococcus phage MX-1. Pyrimidine isostichs and the recognition of a minor pyrimidine. Biochim. Biophys. Acta 1976, 442, 174–183. [Google Scholar] [CrossRef]
- Petrov, V.M.; Ratnayaka, S.; Nolan, J.M.; Miller, E.S.; Karam, J.D. Genomes of the T4-related bacteriophages as windows on microbial genome evolution. Virol. J. 2010, 7, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stellwag, E.; Fink, J.M.; Zissler, J. Physical characterization of the genome of the Myxococcus xanthus bacteriophage MX-8. Mol. Gen. Gen. 1985, 199, 123–132. [Google Scholar] [CrossRef]
- Magrini, V.; Creighton, C.; Youderian, P. Site-Specific recombination of temperate Myxococcus xanthus phage Mx8: Genetic elements required for integration. J. Bacteriol. 1999, 181, 4050–4061. [Google Scholar] [PubMed]
- Tojo, N.; Sanmiya, K.; Sugawara, H.; Inouye, S.; Komano, T. Integration of bacteriophage MX8 into the Myxococcus xanthus chromosome causes a structural alteration at the C-Terminal region of the IntP Protein. J. Bacteriol. 1996, 178, 4004–4011. [Google Scholar] [CrossRef] [PubMed]
- Magrini, V.; Storms, M.L.; Youderian, P. Site-specific recombination of temperate Myxococcus xanthus phage Mx8: Regulation of integrase activity by reversible, covalent modification. J. Bacteriol. 1999, 181, 4062–4070. [Google Scholar] [PubMed]
- Magrini, V.; Salmi, D.; Thomas, D.; Herbert, S.K.; Hartzell, P.L.; Youderian, P. Temperate Myxococcus xanthus phage Mx8 encodes a DNA adenine methylase, Mox. J. Bacteriol. 1997, 179, 4254–4263. [Google Scholar] [CrossRef] [PubMed]
- Nan, B.; Chen, J.; Neu, J.C.; Berry, R.M.; Oster, G.; Zusman, D.R. Myxobacteria gliding motility requires cytoskeleton rotation powered by proton motive force. Proc. Natl. Acad. Sci. USA 2011, 108, 2498–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskella, B.; Taylor, T.B.; Bates, J.; Buckling, A. Using experimental evolution to explore natural patterns between bacterial motility and resistance to bacteriophages. ISME J. 2011, 5, 1809–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, T.B.; Buckling, A. Bacterial motility confers fitness advantage in the presence of phages. J. Evol. Biol. 2013, 26, 2154–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, J.M.; Kalos, M.; Zissler, J.F. Isolation of cell surface antigen mutants of Myxococcus xanthus by use of monoclonal antibodies. J. Bacteriol. 1989, 171, 2033–2041. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Zhang, F. Diversity, classification and evolution of CRISPR-Cas systems. Curr. Opin. Microbiol. 2017, 37, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, R.; Kroos, L. The dev operon regulates the timing of sporulation during Myxococcus xanthus development. J. Bacteriol. 2017, 199. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, R.; Wielgoss, S.; Lippert, G.; Velicer, G.J.; Kroos, L. devI is an evolutionarily young negative regulator of Myxococcus xanthus development. J. Bacteriol. 2015, 197, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.A.; Black, W.P.; Yang, X.; Yang, Z. A CRISPR with roles in Myxococcus xanthus development and exopolysaccharide production. J. Bacteriol. 2014, 196, 4036–4043. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Original Name (New Name) 1 | Source | Life Cycle | Transduction Capacity | Host Range among Lab Strains 2 |
|---|---|---|---|---|
| MX-1 (Mx1) [34] | cow dung [34] | virulent [34] | - | Myxococcus xanthus strains:
Myxococcus fulvus M [43], Myxococcus virescens V2 [43] |
| MX4 (Mx4) [45] | farmyard manure [45] | virulent [45] | - | M. xanthus strains: |
| MX4 hrm-1, MX4* [45] | host range mutant of MX4 [45] | - | generalized [45] | M. xanthus FB (Fru+, Mot+) [45] |
| MX4 ts27htf-1hrm-1 [45] | generalized transduction mutant of MX4 [45] | temperate at 35 °C [45] | generalized [45] | M. xanthus FB (Fru+, Mot+) [45] |
| Mx8 (Mx8) [39] | bacterial isolates from soil sample [39] | temperate [39] | generalized [39] | M. xanthus strains: |
| Mx8 cd [46] | cloudy subtype of MX8 [46] | temperate [46] | generalized [46] | M. xanthus strains [46] |
| Mx8 cr [46] | turbid subtype of Mx8 [46] | temperate [46] | generalized [46] | M. xanthus strains [46] |
| Mx8 a [46] | very turbid subtype of Mx8 [46] | temperate [46] | generalized [46] | M. xanthus strains [46] |
| Mx8 c (clp2) [46] | clear plaquing mutant of Mx8 a [46] | temperate [46] | generalized [46] | M. xanthus strains [46] |
| Mx9 (Mx9) [39] | bacterial isolates from soil sample [39] | temperate [39] | generalized [39] | M. xanthus strains: |
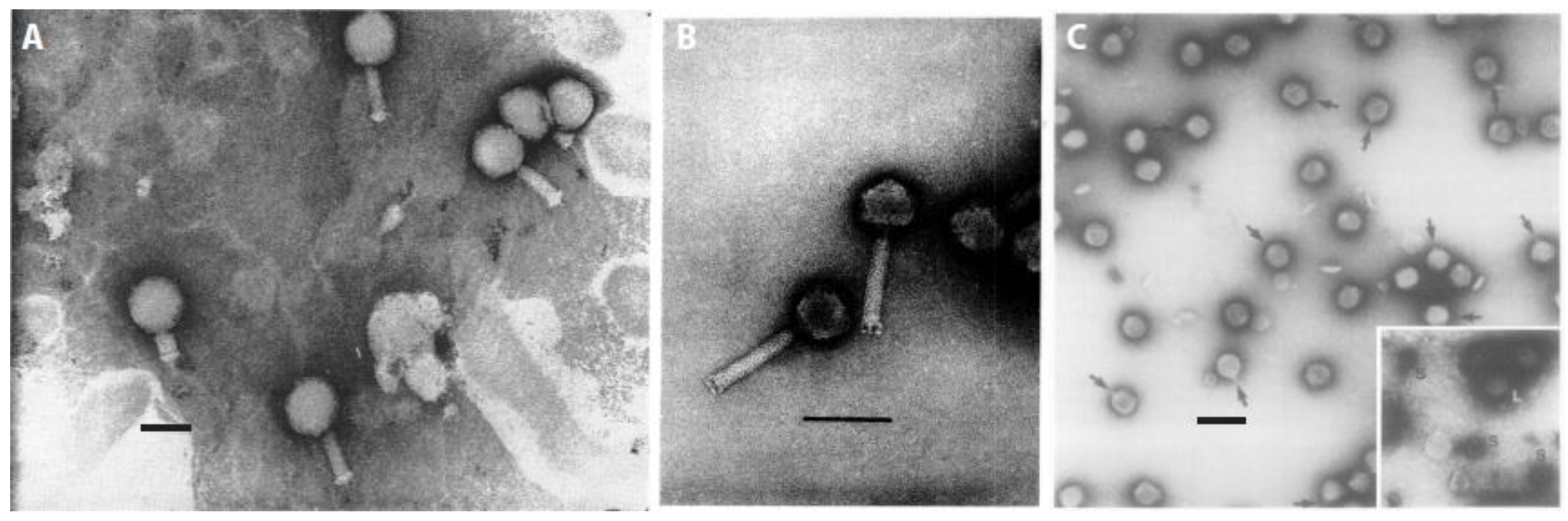
| Original Name (New Name) 1 | Genome 2 | Tail Length [nm] | Tail Structure | Head Diameter [nm] | Head Shape | Plaque Morphology | Similar Morphologies to Enterophages | Morphologically-Related Myxophage Isolates |
|---|---|---|---|---|---|---|---|---|
| MX-1 (Mx1) [34] | dsDNA [34] | 100 [34] | long, contractile [34] | 75–90 [34] | isometric, polyhedral [34] | clear [34] | T-even [50] | From [42]: Mx1l3 |
| From [41]: ϕa, ϕb, ϕm, ϕv, ϕ2 (ϕ = phi) | ||||||||
| From [49]: Mv-1 g1, Mv-1 g2 | ||||||||
| MX4 (Mx4) [45] | dsDNA [47] | 118 [47] | long, contractile [47] | 67 [47] | isometric, icosahedral [47] | clear [45] | T4, λ [45,47] | From [39]: Mx41, Mx43 |
| From [42]: Mx4l2, Mx4l4, Mx4l6, Mx4l7, Mx4l8, Mx4l10, Mx4l11, Mx4l12, Mx4l13, Mx4l14, Mx4k1, Mx4k2, Mx4k3, Mx4k4, Mx4k6, Mx4k10, Mx4k13 | ||||||||
| Mx8 (Mx8) [39] | dsDNA [39] | 10 [39] | short, non-contractile [39] | 60 [39] | isometric, polyhedral [39] | turbid [39] | P22 [39,51] | From [39]: Mx81, Mx82 |
| From [42]: Mx8l1 | ||||||||
| From [49]: Mv-8 g1, Mv-8 g2 | ||||||||
| Mx9 (Mx9) [39] | dsDNA [39,52] | - | very short, non-contractile [39] | 60 [39] | isometric, polyhedral [39] | turbid [39] | P22 [39,51] | - |
| Host Species | Original Phage Name | New Name | Phage Group |
|---|---|---|---|
| Myxococcus xanthus | MX-1 | Mx1 | Mx1-like |
| MX4 | Mx4 | Mx4-like | |
| Mx41 | Mx4-1 | Mx4-like | |
| Mx43 | Mx4-3 | Mx4-like | |
| Mx8 | Mx8 | Mx8-like | |
| Mx81 | Mx8-1 | Mx8-like | |
| Mx82 | Mx8-2 | Mx8-like | |
| Mx9 | Mx9 | Mx9-like | |
| Myxococcus virescens | Mv-1 g1 | Mv1 | Mv1-like |
| Mv-1 g2 | Mv1-1 | Mv1-like | |
| Mv-8 g1 | Mv8 | Mv8-like | |
| Mv-8 g2 | Mv8-1 | Mv8-like |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasse, M.; Wielgoss, S. Bacteriophages of Myxococcus xanthus, a Social Bacterium. Viruses 2018, 10, 374. https://doi.org/10.3390/v10070374
Vasse M, Wielgoss S. Bacteriophages of Myxococcus xanthus, a Social Bacterium. Viruses. 2018; 10(7):374. https://doi.org/10.3390/v10070374
Chicago/Turabian StyleVasse, Marie, and Sébastien Wielgoss. 2018. "Bacteriophages of Myxococcus xanthus, a Social Bacterium" Viruses 10, no. 7: 374. https://doi.org/10.3390/v10070374
APA StyleVasse, M., & Wielgoss, S. (2018). Bacteriophages of Myxococcus xanthus, a Social Bacterium. Viruses, 10(7), 374. https://doi.org/10.3390/v10070374