Overview of Trends in the Application of Metagenomic Techniques in the Analysis of Human Enteric Viral Diversity in Africa’s Environmental Regimes

 ,
,  ,
, 

Abstract
:1. Introduction
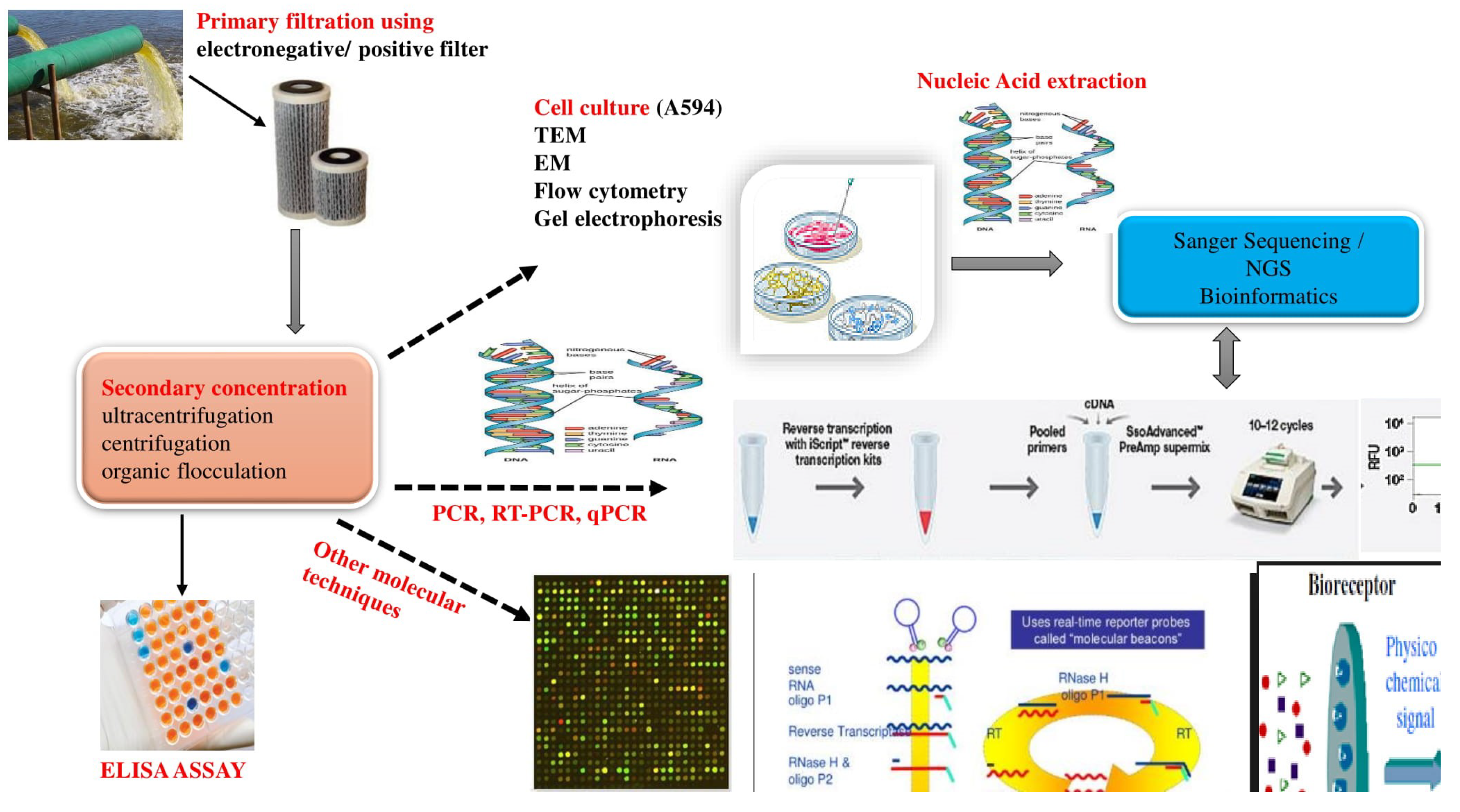
2. Conventional Methods for the Identification of HEV in Environmental Samples
2.1. Culture Based Methods
2.2. Polymerase Chain Reaction Methods (PCR Assays)
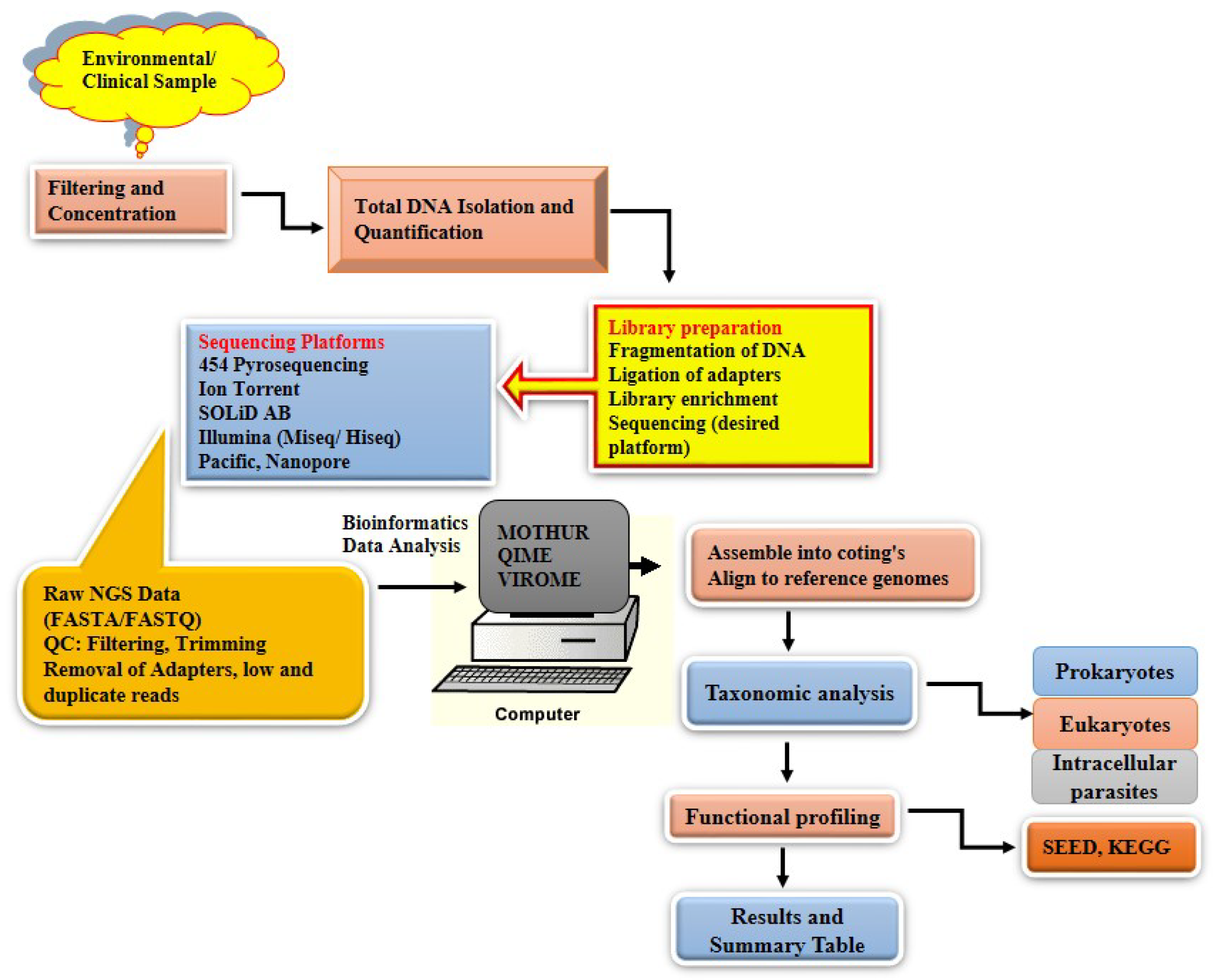
2.3. Viral Metagenomics
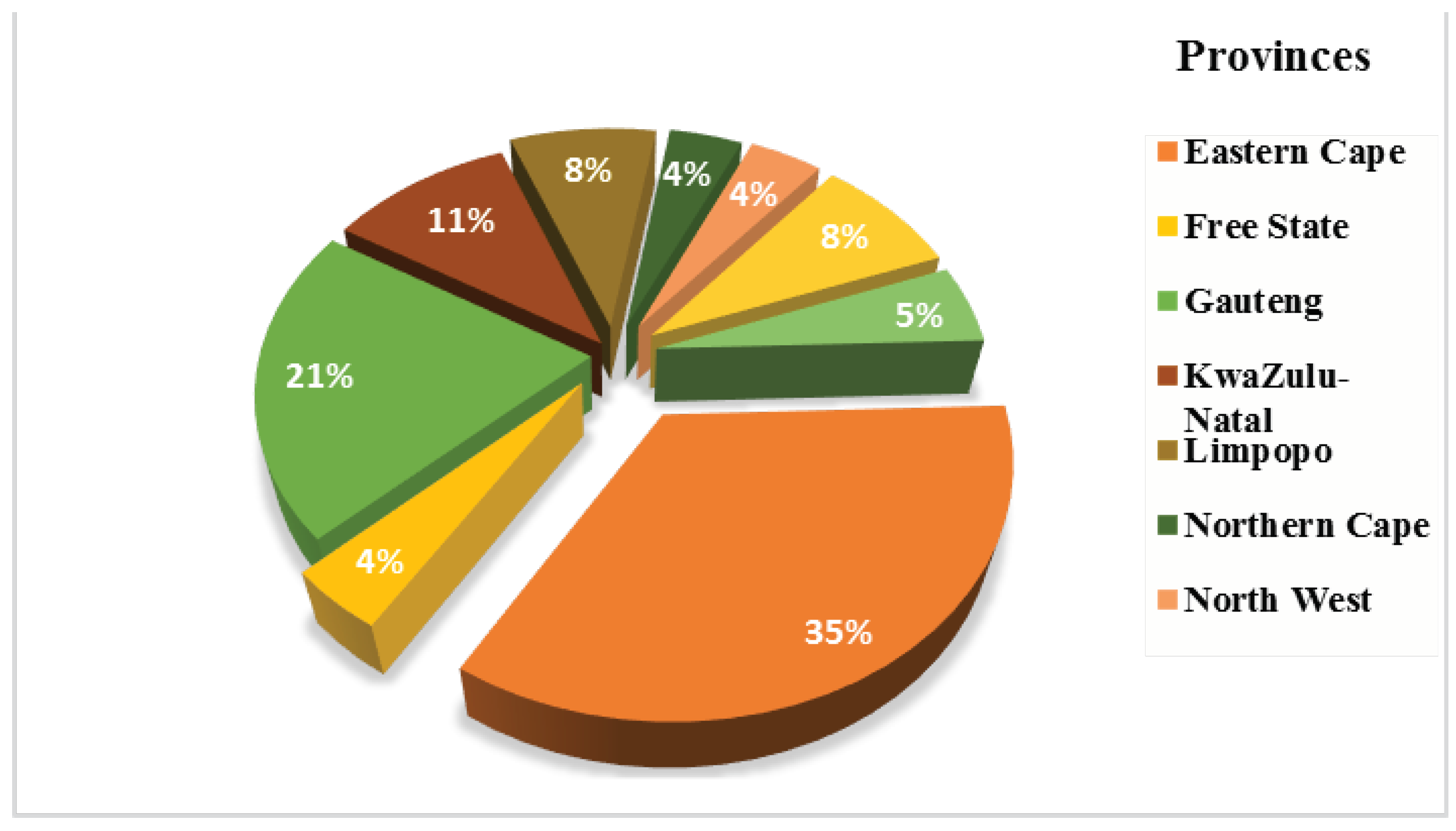
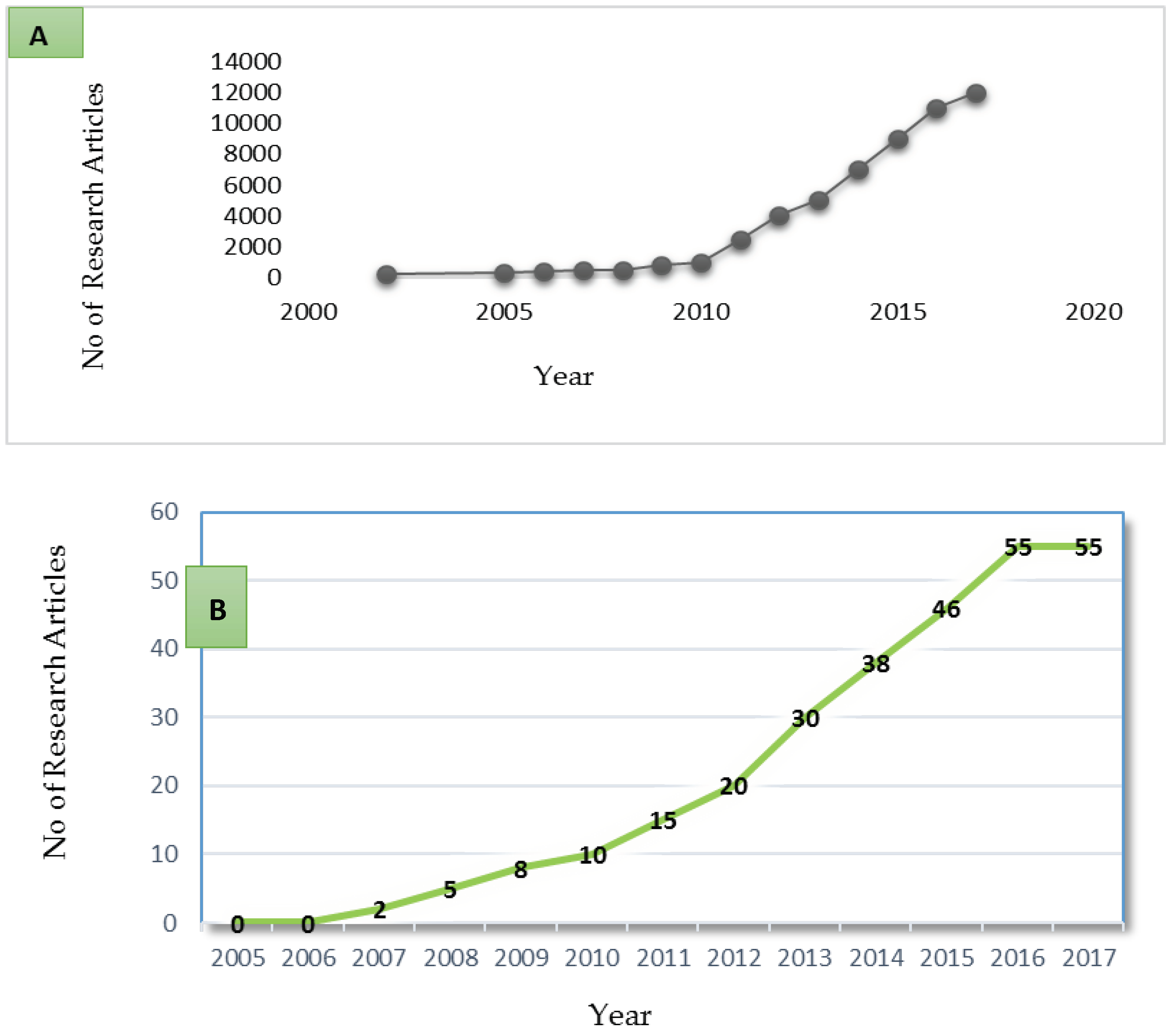
3. Metagenomics and Its Application in Africa
4. Open Research Work and Implications for Environmental Genomes
5. Conclusions and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Thomas, T.; Gilbert, J.; Meyer, F. Metagenomics—A guide from sampling to data analysis. Microbiol. Inf. Exp. 2012, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Bashir, Y.; Pradeep Singh, S.; Kumar Konwar, B. Metagenomics: An application based perspective. Chin. J. Biol. 2014, 2014, 1–7. [Google Scholar] [CrossRef]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol. Mol. Biol. Rev. 2005, 69, 195. [Google Scholar] [CrossRef]
- Alma’abadi, A.D.; Gojobori, T.; Mineta, K. Marine metagenome as a resource for novel enzymes. Genom. Proteom. Bioinform. 2015, 13, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Bajic, V.B. The value and significance of metagenomics of marine environments. Genom. Proteom. Bioinform. 2015, 13, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Flemer, B.; Jackson, S.A.; Lejon, D.P.H.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D.W. Marine metagenomics: New tools for the study and exploitation of marine microbial metabolism. Mar. Drugs 2010, 8, 608–628. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Bharucha, T.; Breuer, J. Encephalitis diagnosis using metagenomics: Application of next generation sequencing for undiagnosed cases. J. Infect. 2018, 76, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.R.; Montoya, V.; Gardy, J.L.; Patrick, D.M.; Tang, P. Metagenomics for pathogen detection in public health. Genome Med. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.F.; Kondov, N.O.; Deng, X.; Van Eenennaam, A.; Neibergs, H.L.; Delwart, E. A metagenomics and case-control study to identify viruses associated with bovine respiratory disease. J. Virol. 2015, 89, 5340–5349. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. The structure and diversity of human, animal and environmental resistomes. Microbiome 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Arenas, M.; Pereira, F.; Oliveira, M.; Pinto, N.; Lopes, A.M.; Gomes, V.; Carracedo, A.; Amorim, A. Forensic genetics and genomics: Much more than just a human affair. PLoS Genet. 2017, 13, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Cunha, M.V.; Inácio, J. Veterinary infection biology: Molecular diagnostics and high-throughput strategies. Vet. Infect. Biol. 2014. [Google Scholar] [CrossRef]
- Blomström, A.L. Viral metagenomics as an emerging and powerful tool in veterinary medicine. Vet. Q. 2011, 31, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Northbrook, H.C.; Berry, J.K. International Conference on Earth Science & Climate Change; The Geological Society: London, UK, 2012. [Google Scholar]
- Lorenz, P.; Eck, J. Outlook: Metagenomics and industrial applications. Nat. Rev. Microbiol. 2005, 3, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.M.; Cotter, P.D.; Hill, C.; Alvarez-Ordóñez, A. Biotechnological applications of functional metagenomics in the food and pharmaceutical industries. Front. Microbiol. 2015, 6, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, J.K.; Taylor, N.D.; Church, G.M. Biosensor-based engineering of biosynthetic pathways. Curr. Opin. Biotechnol. 2016, 42, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Stobbe, A.H.; Roossinck, M.J. Plant virus metagenomics: What we know and why we need to know more. Front. Plant Sci. 2014, 5, 150. [Google Scholar] [CrossRef] [PubMed]
- Duhaime, M.B.; Sullivan, M.B. Ocean viruses: Rigorously evaluating the metagenomic sample-to-sequence pipeline. Virology 2012, 434, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Behjati, S.; Tarpey, P.S. What is next generation sequencing? Arch. Dis. Child. Educ. Pract. Ed. 2013, 98, 236–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.A.; Spacek, D.; Snyder, M.P. High-throughput sequencing technologies. Mol. Cell 2016, 58, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Ambardar, S.; Gupta, R.; Trakroo, D.; Lal, R.; Vakhlu, J. High throughput sequencing: An overview of sequencing chemistry. Indian J. Microbiol. 2016, 56, 394–404. [Google Scholar] [CrossRef] [PubMed]
- van der Walt, A.J.; van Goethem, M.W.; Ramond, J.B.; Makhalanyane, T.P.; Reva, O.; Cowan, D.A. Assembling metagenomes, one community at a time. BMC Genom. 2017, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.P.A.; van Dongen, S.; Abreu-Goodger, C.; Bartonicek, N.; Enright, A.J. Kraken: A set of tools for quality control and analysis of high-throughput sequence data. Methods 2013, 63, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Oulas, A.; Pavloudi, C.; Polymenakou, P.; Pavlopoulos, G.A.; Papanikolaou, N.; Kotoulas, G.; Arvanitidis, C.; Iliopoulos, I. Metagenomics: Tools and insights for analyzing next-generation sequencing data derived from biodiversity studies. Bioinform. Biol. Insights 2015, 9, 75–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.C.; Lai, K.; Lorenc, M.T.; Imelfort, M.; Duran, C.; Edwards, D. Bioinformatics tools and databases for analysis of next-generation sequence data. Brief. Funct. Genom. 2012, 11, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, J.; Yang, Y.; Tang, Y.; Shang, J.; Shen, B. A practical comparison of De Novo genome assembly software tools for next-generation sequencing technologies. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Li, J.; Shen, H.; Zhang, L.; Papasian, C.J.; Deng, H.W. Comparative studies of de novo assembly tools for next-generation sequencing technologies. Bioinformatics 2011, 27, 2031–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thankaswamy-Kosalai, S.; Sen, P.; Nookaew, I. Evaluation and assessment of read-mapping by multiple next-generation sequencing aligners based on genome-wide characteristics. Genomics 2017, 109, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Diniz, W.J.S.; Canduri, F. Bioinformatics: An overview and its applications. Genet. Mol. Res. 2017, 16, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, B.P. Bioinformatics impacts on medicine, microbial genome and agriculture. J. Pharm. Phytochem. 1938, 6, 1938–1942. [Google Scholar]
- Ludwig, W.; Strunk, O.; Westram, R.; Richter, L.; Meier, H.; Yadhukumar, A.; Buchner, A.; Lai, T.; Steppi, S.; Jacob, G.; et al. ARB: A software environment for sequence data. Nucl. Acids Res. 2004, 32, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Ounit, R.; Wanamaker, S.; Close, T.J.; Lonardi, S. CLARK: fast and accurate classification of metagenomic and genomic sequences using discriminative k-mers. BMC Genom. 2015, 16, 236. [Google Scholar] [CrossRef] [PubMed]
- Rosen, G.L.; Reichenberger, E.R.; Rosenfeld, A.M. NBC: The naïve Bayes classification tool webserver for taxonomic classification of metagenomic reads. Bioinformatics 2011, 27, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, D.; Sternberg, M.J.E.; Raczy, C.; Butcher, S.A. k-SLAM: Accurate and ultra-fast taxonomic classification and gene identification for large metagenomic data sets. Nucl. Acids Res. 2017, 45, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.; Salzberg, S. Classification with Interpolated Markov Models. Nat. Methods 2009, 6, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Murray-Rust, P. Bioinformatics and drug discovery. Curr. Opin. Biotechnol. 1994, 5, 648–653. [Google Scholar] [CrossRef]
- Katara, P. Role of bioinformatics and pharmacogenomics in drug discovery and development process. Netw. Model. Anal. Health Inf. Bioinform. 2013, 2, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Moffat, J.G.; Rudolph, J.; Bailey, D. Phenotypic screening in cancer drug discovery—Past, present and future. Nat. Rev. Drug Dis. 2014, 13, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Rajamanickam, E. Application of bioinformatics in agriculture. In ICT for Agricultural Development in Changing Climate; Singh, K.M., Meena, M.S., Eds.; Narendra Publishing House: New Delhi, India, 2012; pp. 163–179. [Google Scholar]
- McArthur, A.G.; Wright, G.D. Bioinformatics of antimicrobial resistance in the age of molecular epidemiology. Curr. Opin. Microbiol. 2015, 27, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, J.; Chen, H.; Bond, P.L.; Yuan, Z. Metagenomic analysis reveals wastewater treatment plants as hotspots of antibiotic resistance genes and mobile genetic elements. Water Res. 2017, 123, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S. Role of bioinformatics in climate change studies. J. Sci. 2015, 1, 1–8. [Google Scholar]
- Weinbauer, M.G.; Rassoulzadegan, F. Are viruses driving microbial diversification and diversity? Environ. Microbiol. 2004, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, F.; Prangishvili, D.; Lindell, D. Roles of viruses in the environment. Environ. Microbiol. 2009, 11, 2771–2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttle, C.A. Viruses in the sea. Nature 2005, 437, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, D. Expression of animal virus genomes. Bacteriol. Rev. 1971, 35, 235–241. [Google Scholar] [PubMed]
- Altintas, Z.; Gittens, M.; Pocock, J.; Tothill, I.E. Biosensors for waterborne viruses: Detection and removal. Biochimie 2015, 115, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Fong, T.-T.; Lipp, E.K. Enteric viruses of humans and animals in aquatic environments: Health risks, detection, and potential water quality assessment tools. Microbiol. Mol. Biol. Rev. 2005, 69, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A. Human enteric viruses in the water environment: A minireview. Int. Microbiol. 1998, 1, 191–196. [Google Scholar] [PubMed]
- Gall, A.M.; Mariñas, B.J.; Lu, Y.; Shisler, J.L. Waterborne viruses: A barrier to safe drinking water. PLoS Pathog. 2015, 11, e1004867. [Google Scholar] [CrossRef] [PubMed]
- Caul, E.O. Viral gastroenteritis: Small round structured viruses, caliciviruses and astroviruses. Part II. The epidemiological perspective. J. Clin. Pathol. 1996, 49, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Gibson, K.E.; Schwab, K.J. Detection of bacterial indicators and human and bovine enteric viruses in surface water and groundwater sources potentially impacted by animal and human wastes in Lower Yakima Valley, Washington. Appl. Environ. Microbiol. 2011, 77, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Chigor, V.N.; Sibanda, T.; Okoh, A.I. Assessment of the risks for human health of adenoviruses, hepatitis a virus, rotaviruses and enteroviruses in the buffalo river and three source water dams in the eastern cape. Food Environ. Virol. 2014, 6, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Osuolale, O.; Okoh, A. Human enteric bacteria and viruses in five wastewater treatment plants in the Eastern Cape, South Africa. J. Infect. Public Health 2017, 10, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Osuolale, O.; Okoh, A. Incidence of human adenoviruses and hepatitis a virus in the final effluent of selected wastewater treatment plants in Eastern Cape Province, South Africa. Virol. J. 2015, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Okoh, A.I.; Sibanda, T.; Gusha, S.S. Inadequately treated wastewater as a source of human enteric viruses in the environment. Int. J. Environ. Res. Public Health 2010, 7, 2620–2637. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.B.; de Mendes, A.C.O.; Guimarães, F.R.; Fumian, T.M.; Leite, J.P.G.; Gaspar, A.M.C.; Miagostovich, M.P. Detection of enteric viruses in recreational waters of an urban lagoon in the city of Rio de Janeiro, Brazil. Mem. Inst. Oswaldo Cruz 2012, 107, 778–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajko-Nenow, P.; Waters, A.; Keaveney, S.; Flannery, J.; Tuite, G.; Coughlan, S.; O’Flaherty, V.; Doré, W. Norovirus genotypes present in oysters and in effluent from a wastewater treatment plant during the seasonal peak of infections in ireland in 2010. Appl. Environ. Microbiol. 2013, 79, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- Kukkula, M.; Maunula, L.; Silvennoinen, E.; von Bonsdorff, C. Outbreak of viral gastroenteritis due to drinking water contaminated by norwalk-like viruses. J. Infect. Dis. 1999, 180, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.C.; O’Brien, S.J. Economic cost of campylobacter, norovirus and rotavirus disease in the United Kingdom. PLoS ONE 2016, 11, e0138526. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.J. Enterically infecting viruses: Pathogenicity, transmission and significance for food and waterborne infection. J. Appl. Microbiol. 2005, 98, 1354–1380. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, J.; Bell, D.; Simmons, G.C.; Rivera-Aban, M.; Wolf, S.; Greening, G.E. Gastroenteritis outbreak caused by waterborne norovirus at a New Zealand Ski Resort. Appl. Environ. Microbiol. 2007, 73, 7853–7857. [Google Scholar] [CrossRef] [PubMed]
- Hamza, I.A.; Jurzik, L.; Überla, K.; Wilhelm, M. Methods to detect infectious human enteric viruses in environmental water samples. Int. J. Hyg. Environ. Health 2011, 214, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Guix, S.; Sano, D.; Pintó, R.M. New tools for the study and direct surveillance of viral pathogens in water. Curr. Opin. Biotechnol. 2008, 19, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Van Heerden, J.; Ehlers, M.M.; Heim, A.; Grabow, W.O.K. Prevalence, quantification and typing of adenoviruses detected in river and treated drinking water in South Africa. J. Appl. Microbiol. 2005, 99, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chigor, V.N.; Okoh, A.I. Quantitative RT-PCR detection of hepatitis A virus, rotaviruses and enteroviruses in the Buffalo River and source water dams in the Eastern Cape Province of South Africa. Int. J. Environ. Res. Public Health 2012, 9, 4017–4032. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, M.M.; Grabow, W.O.K.; Pavlov, D.N. Detection of enteroviruses in untreated and treated drinking water supplies in South Africa. Water Res. 2005, 39, 2253–2258. [Google Scholar] [CrossRef] [PubMed]
- van Heerden, J.; Ehlers, M.M.; van Zyl, W.B.; Grabow, W.O.K. Prevalence of human adenoviruses in raw and treated water. Water Sci. Technol. 2004, 50, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, D.N.; Van Zyl, W.B.; Van Heerden, J.; Grabow, W.O.K.; Ehlers, M.M. Prevalence of vaccine-derived polioviruses in sewage and river water in South Africa. Water Res. 2005, 39, 3309–3319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibanda, T.; Okoh, A.I. Real-time PCR quantitative assessment of hepatitis A virus, rotaviruses and enteroviruses in the Tyume River located in the Eastern Cape Province, South Africa. Water SA 2013, 39, 295–304. [Google Scholar] [CrossRef]
- Taylor, M.B.; Schildhauer, C.I.; Parker, S.; Grabow, W.O.K.; Jiang, X.; Estes, M.K.; Cubitt, W.D. Two successive outbreaks of SRSV-associated gastroenteritis in South Africa. J. Med. Virol. 1993, 41, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.B.; Cox, N.; Vrey, M.A.; Grabow, W.O.K. The occurrence of hepatitis A and astroviruses in selected river and dam waters in South Africa. Water Res. 2001, 35, 2653–2660. [Google Scholar] [CrossRef]
- Mans, J.; de Villiers, J.C.; du Plessis, N.M.; Avenant, T.; Taylor, M.B. Emerging norovirus GII.4 2008 variant detected in hospitalised paediatric patients in South Africa. J. Clin. Virol. 2010, 49, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mans, J.; Murray, T.Y.; Taylor, M.B. Novel norovirus recombinants detected in South Africa. Virol. J. 2014, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Sánchez, G.; Abbaszadegan, M.; Carducci, A.; Guix, S.; Le Guyader, F.S.; Netshikweta, R.; Pintó, R.M.; van der Poel, W.H.M.; Rutjes, S.; et al. Analytical methods for virus detection in water and food. Food Anal. Methods 2011, 4, 4–12. [Google Scholar] [CrossRef]
- Cashdollar, J.L.; Wymer, L. Methods for primary concentration of viruses from water samples: A review and meta-analysis of recent studies. J. Appl. Microbiol. 2013, 115, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wallis, C.; Melnick, J.L. Concentration of enteroviruses on membrane filters. J. Virol. 1967, 1, 472–477. [Google Scholar] [PubMed]
- Wallis, C.; Melnick, J.L.; Fields, J.E. Detection of viruses in large volumes of natural waters by concentration on insoluble polyelectrolytes. Water Res. 1970, 4, 787–796. [Google Scholar] [CrossRef]
- Ikner, L.A.; Gerba, C.P.; Bright, K.R. Concentration and recovery of viruses from water: A comprehensive review. Food Environ. Virol. 2012, 4, 41–67. [Google Scholar] [CrossRef] [PubMed]
- Ikner, L.A.; Soto-Beltran, M.; Bright, K.R. New method using a positively charged microporous filter and ultrafiltration for concentration of viruses from tap water. Appl. Environ. Microbiol. 2011, 77, 3500–3506. [Google Scholar] [CrossRef] [PubMed]
- Cashdollar, J.L.; Brinkman, N.E.; Griffin, S.M.; Mcminn, B.R.; Rhodes, E.R.; Varughese, E.A.; Grimm, A.C.; Parshionikar, S.U.; Wymer, L.; Shay Fout, G. Development and evaluation of EPA method 1615 for detection of enterovirus and norovirus in water. Appl. Environ. Microbiol. 2013, 79, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Cashdollar, J.L.; Dahling, D.R. Evaluation of a method to re-use electropositive cartridge filters for concentrating viruses from tap and river water. J. Virol. Methods 2006, 132, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Wyn-Jones, A.P.; Carducci, A.; Cook, N.; D’Agostino, M.; Divizia, M.; Fleischer, J.; Gantzer, C.; Gawler, A.; Girones, R.; Höller, C.; et al. Surveillance of adenoviruses and noroviruses in European recreational waters. Water Res. 2011, 45, 1025–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fout, G.S.; Cashdollar, J.L.; Varughese, E.A.; Parshionikar, S.U.; Grimm, A.C. EPA method 1615. Measurement of enterovirus and norovirus occurrence in water by culture and RT-qPCR. I. collection of virus samples. J. Vis. Exp. 2016, 107, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Adefisoye, M.A.; Nwodo, U.U.; Green, E.; Okoh, A.I. Quantitative PCR detection and characterisation of human adenovirus, rotavirus and hepatitis a virus in discharged effluents of two wastewater treatment facilities in the Eastern Cape, South Africa. Food Environ. Virol. 2016, 8, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Hill, W.F.; Akin, E.W.; Benton, W.H. Detection of viruses in water: A review of methods and application*. Water Res. Pergam. Press 1971, 5, 967–995. [Google Scholar] [CrossRef]
- Shuval, H.; Cymbalista, B.; Goldblum, N. The phase separation method for the concentration and detection of viruses in water. Water Res. 1969, 3, 225–240. [Google Scholar] [CrossRef]
- Sobsey, M.D.; Jones, B.L. Concentration of poliovirus from tap water using positively charged microporous filters. Appl. Environ. Microbiol. 1979, 37, 588–595. [Google Scholar] [PubMed]
- Karim, M.R.; Rhodes, E.R.; Brinkman, N.; Wymer, L.; Fout, G.S. New electropositive filter for concentrating enteroviruses and noroviruses from large volumes of water. Appl. Environ. Microbiol. 2009, 75, 2393–2399. [Google Scholar] [CrossRef] [PubMed]
- Albinana-Gimenez, N.; Clemente-Casares, P.; Calgua, B.; Huguet, J.M.; Courtois, S.; Girones, R. Comparison of methods for concentrating human adenoviruses, polyomavirus JC and noroviruses in source waters and drinking water using quantitative PCR. J. Virol. Methods 2009, 158, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Kiulia, N.M.; Netshikweta, R.; Page, N.A.; Van Zyl, W.B.; Kiraithe, M.M.; Nyachieo, A.; Mwenda, J.M.; Taylor, M.B. The detection of enteric viruses in selected urban and rural river water and sewage in Kenya, with special reference to rotaviruses. J. Appl. Microbiol. 2010, 109, 818–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin, D.; Fuchs, F.; Crance, J.M.; Deloince, R.; Aymard, M.; Bartoli, M. Validation of an ultrafiltration process to concentrate viruses from large volumes of water. Environ. Technol. 1993, 14, 397–400. [Google Scholar] [CrossRef]
- Cai, L.; Yang, Y.; Jiao, N.; Zhang, R. Evaluation of tangential flow filtration for the concentration and separation of bacteria and viruses in contrasting marine environments. PLoS ONE 2015, 10, e0136741. [Google Scholar] [CrossRef] [PubMed]
- Prata, C.; Ribeiro, A.; Cunha, Â.; Gomes, N.C.M.; Almeida, A. Ultracentrifugation as a direct method to concentrate viruses in environmental waters: Virus-like particle enumeration as a new approach to determine the efficiency of recovery. J. Environ. Monit. 2012, 14, 64. [Google Scholar] [CrossRef] [PubMed]
- Hryniszyn, A.; Skonieczna, M.; Wiszniowski, J. Methods for detection of viruses in water and wastewater. Adv. Microbiol. 2013, 3, 442–449. [Google Scholar] [CrossRef]
- Hematian, A.; Sadeghifard, N.; Mohebi, R.; Taherikalani, M.; Nasrolahi, A.; Amraei, M.; Ghafourian, S. Traditional and modern cell culture in virus diagnosis. Osong Public Health Res. Perspect. 2016, 7, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Lee, S.H.; Han, E.; Kim, S.J. Use of cell culture-PCR assay based on combination of A549 and BGMK cell lines and molecular identification as a tool to monitor infectious adenoviruses and enteroviruses in river water. Appl. Environ. Microbiol. 2004, 70, 6695–6705. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, K. A integrated cell culture/PCR for detection of enteric viruses in environmental samples. Methods Mol. Biol. 2004, 268, 69–78. [Google Scholar] [PubMed]
- Chapron, C.D.; Ballester, N.A.; Fontaine, J.H.; Frades, C.N.; Margolin, A.B. Detection of astroviruses, enteroviruses, and adenovirus types 40 and 41 in surface waters collected and evaluated by the information collection rule and an integrated cell culture-nested PCR procedure. Appl. Environ. Microbiol. 2000, 66, 2520–2525. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, K.A.; Gerba, C.P.; Pepper, I.A.N.L. Detection of infectious enteroviruses by an integrated cell culture-PCR procedure. Appl. Environ. Microbiol. 1996, 62, 1424–1427. [Google Scholar] [PubMed]
- Lee, S.H.; Lee, C.; Lee, K.W.; Cho, H.B.; Kim, S.J. The simultaneous detection of both enteroviruses and adenoviruses in environmental water samples including tap water with an integrated cell culture-multiplex-nested PCR procedure. J. Appl. Microbiol. 2005, 98, 1020–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.K.; Jeong, Y.S. Comparison of total culturable virus assay and multiplex integrated cell culture-PCR for reliability of waterborne virus detection. Appl. Environ. Microbiol. 2004, 70, 3632–3636. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, R.A.; Pepper, I.L.; Gerba, C.P. Application of PCR-based methods to assess the infectivity of enteric viruses in environmental samples. Appl. Environ. Microbiol. 2009, 75, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Bettarel, Y.; Sime-ngando, T.; Amblard, C.; Laveran, H. A comparison of methods for counting viruses in aquatic systems a comparison of methods for counting viruses in aquatic systems. Appl. Environ. Microbiol. 2000, 66, 2283–2289. [Google Scholar] [CrossRef] [PubMed]
- Budinoff, C.R.; Loar, S.N.; LeCleir, G.R.; Wilhelm, S.W.; Buchan, A. A protocol for enumeration of aquatic viruses by epifluorescence microscopy using AnodiscTM 13 membranes. BMC Microbiol. 2011, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Jian-Wen, H.; Jiang, S. Quantification of enterococci and human adenoviruses in environmental samples by quantification of enterococci and human adenoviruses in environmental samples by real-time PCR. Appl. Environ. Microbiol. 2005, 71, 2250–2255. [Google Scholar]
- Bettarel, Y.; Bouvier, T.; Bouvier, C.; Carré, C.; Desnues, A.; Domaizon, I.; Jacquet, S.; Robin, A.; Sime-Ngando, T. Ecological traits of planktonic viruses and prokaryotes along a full-salinity gradient. FEMS Microbiol. Ecol. 2011, 76, 360–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CPD, B. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar]
- Paul, J.H.; Jiang, S.C.; Rose, J.B. Concentration of viruses and dissolved DNA from aquatic environments by vortex flow filtration. Appl. Environ. Microbiol. 1991, 57, 2197–2204. [Google Scholar] [PubMed]
- Staggemeier, R.; Bortoluzzi, M.; da Heck, T.M.S.; Spilki, F.R.; de Almeida, S.E.M. Quantitative vs. conventional pcr for detection of human adenoviruses in water and sediment samples. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Farkas, K.; Hassard, F.; McDonald, J.E.; Malham, S.K.; Jones, D.L. Evaluation of molecular methods for the detection and quantification of pathogen-derived nucleic acids in sediment. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Schlindwein, A.D.; Rigotto, C.; Simões, C.M.; Barardi, C.R.M. Detection of enteric viruses in sewage sludge and treated wastewater effluent. Water Sci. Technol. 2010, 61, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Pintó, R.M.; Costafreda, M.I.; Bosch, A. Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl. Environ. Microbiol. 2009, 75, 7350–7355. [Google Scholar] [CrossRef] [PubMed]
- Abbazadegan, M.; Huber, M.S.; Gerba, C.P.; Pepper, I.L. Detection of enteroviruses in groundwater using the polymerase chain reaction. Appl. Environ. Microbiol. 1993, 59, 1318–1324. [Google Scholar]
- Iqbal, S.S.; Mayo, M.W.; Bruno, J.G.; Bronk, B.V.; Batt, C.A.; Chambers, J.P. A review of molecular recognition technologies for detection of biological threat agents. Biosens. Bioelectron. 2000, 15, 549–578. [Google Scholar] [CrossRef]
- Elnifro, E.M.; Ashshi, A.M.; Cooper, R.J.; Klapper, P.E. Multiplex PCR: Optimization and application in diagnostic virology. Clin. Microbiol. Rev. 2000, 13, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Mothershed, E.A.; Whitney, A.M. Nucleic acid-based methods for the detection of bacterial pathogens: Present and future considerations for the clinical laboratory. Clin. Chim. Acta 2006, 363, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Girones, R.; Ferrús, M.A.; Alonso, J.L.; Rodriguez-Manzano, J.; Calgua, B.; de Abreu Corrêa, A.; Hundesa, A.; Carratala, A.; Bofill-Mas, S. Molecular detection of pathogens in water—The pros and cons of molecular techniques. Water Res. 2010, 44, 4325–4339. [Google Scholar] [CrossRef] [PubMed]
- Costafreda, M.I.; Bosch, A.; Pintó, R.M. Development, evaluation, and standardization of a real-time TaqMan reverse transcription-PCR assay for quantification of hepatitis A virus in clinical and shellfish samples. Appl. Environ. Microbiol. 2006, 72, 3846–3855. [Google Scholar] [CrossRef] [PubMed]
- Said, R.; Wolfaardt, M.; Taylor, M.B. Molecular characterisation of hepatitis A virus strains from water sources in South Africa. Water Sci. Technol. 2014, 69, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.B.; Litaker, R.W.; Noble, R.T. Rapid one-step quantitative reverse transcriptase PCR assay with competitive internal positive control for detection of enteroviruses in environmental samples. Appl. Environ. Microbiol. 2006, 72, 3960–3967. [Google Scholar] [CrossRef] [PubMed]
- Monpoeho, S.; Maul, A.; Mignotte-Cadiergues, B.; Schwartzbrod, L.; Billaudel, S.; Ferré, V. Best viral elution method available for quantification of enteroviruses in sludge by both cell culture and reverse transcription-PCR. Appl. Environ. Microbiol. 2001, 67, 2484–2488. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Kang, G.; Hill, V.R. Broadly reactive TaqMan®assay for real-time RT-PCR detection of rotavirus in clinical and environmental samples. J. Virol. Methods 2009, 155, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Victoria, M.; Guimarães, F.R.; Fumian, T.M.; Ferreira, F.F.M.; Vieira, C.B.; Leite, J.P.G.; Miagostovich, M.P.; Shubo, T. One year monitoring of norovirus in a sewage treatment plant in Rio de Janeiro, Brazil. J. Water Health 2010, 8, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Hill, V.R.; Lu, X.; Sobsey, M.D.; Erdman, D.D. Quantitative real-time PCR assays for detection of human adenoviruses and identification of serotypes 40 and 41. Appl. Environ. Microbiol. 2005, 71, 3131–3136. [Google Scholar] [CrossRef] [PubMed]
- Allard, A.; Albinsson, B.; Wadell, G.; Albinsson, B.O. Rapid typing of human adenoviruses by a general PCR combined with restriction endonuclease analysis rapid typing of human adenoviruses by a general PCR combined with restriction endonuclease analysis. J. Clin. Microbiol. 2001, 39, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.; O’Leary, J.J.; O’Sullivan, N. Real-time reverse transcription PCR detection of norovirus, sapovirus and astrovirus as causative agents of acute viral gastroenteritis. J. Virol. Methods 2007, 146, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Wang, Q.; Katayama, K.; Saif, L.J. Comprehensive review of human sapoviruses. Clin. Microbiol. Rev. 2015, 28, 32–53. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.M.; Ao, Y.Y.; Liu, N.; Li, L.L.; Duan, Z.J. Salivirus in children and its association with childhood acute gastroenteritis: A paired case-control study. PLoS ONE 2015, 10, e0130977. [Google Scholar] [CrossRef] [PubMed]
- Haramoto, E.; Kitajima, M.; Otagiri, M. Development of a reverse transcription-quantitative PCR assay for detection of salivirus/klassevirus. Appl. Environ. Microbiol. 2013, 79, 3529–3532. [Google Scholar] [CrossRef] [PubMed]
- Benschop, K.; Thomas, X.; Serpenti, C.; Molenkamp, R.; Wolthers, K. High prevalence of human parechovirus (HPeV) genotypes in the Amsterdam region and identification of specific HPeV variants by direct genotyping of stool samples. J. Clin. Microbiol. 2008, 46, 3965–3970. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.C.Y.; Lo, K.L.; Que, T.L.; Lee, R.A.; Chan, K.H.; Yuen, K.Y.; Woo, P.C.Y.; Lau, S.K.P. Epidemiology of human parechovirus, Aichi virus and salivirus in fecal samples from hospitalized children with gastroenteritis in Hong Kong. Virol. J. 2014, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Markoulatos, P.; Siafakas, N.; Moncany, M. Multiplex polymerase chain reaction: A practical approach. J. Clin. Lab. Anal. 2002, 16, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Castillo, F.; Loera-Muro, A.; Jacques, M.; Garneau, P.; Avelar-González, F.; Harel, J.; Guerrero-Barrera, A. Waterborne pathogens: Detection methods and challenges. Pathogens 2015, 4, 307–334. [Google Scholar] [CrossRef] [PubMed]
- Haramoto, E.; Katayama, H.; Oguma, K.; Ohgaki, S. Quantitative analysis of human enteric adenoviruses in aquatic environments. J. Appl. Microbiol. 2007, 103, 2153–2159. [Google Scholar] [CrossRef] [PubMed]
- Albinana-Gimenez, N.; Miagostovich, M.P.; Calgua, B.; Huguet, J.M.; Matia, L.; Girones, R. Analysis of adenoviruses and polyomaviruses quantified by qPCR as indicators of water quality in source and drinking-water treatment plants. Water Res. 2009, 43, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Olaniran, A.O.; Naidoo, S.; Pillay, B. Surveillance of invasive bacterial pathogens and human enteric viruses in wastewater final effluents and receiving water bodies—A case study from Durban, South Africa. Clean Soil Air Water 2012, 40, 681–691. [Google Scholar] [CrossRef]
- Soto-Beltran, M.; Ikner, L.A.; Bright, K.R. Effectiveness of poliovirus concentration and recovery from treated wastewater by two electropositive filter methods. Food Environ. Virol. 2013, 5, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Cashdollar, J.L.; Brinkman, N.E.; Griffin, S.M.; McMinn, B.R.; Rhodes, E.R.; Varughese, E.A.; Grimm, A.C.; Parshionikar, S.U.; Wymer, L.; Fout, G.S. Development and Evaluation of EPA Method 1615 for Detection of Enterovirus and Norovirus in Water. Appl. Environ. Microbiol. [CrossRef] [PubMed]
- Suttle, C.A.; Fuhrman, J.A. Enumeration of virus particles in aquatic or sediment samples by epifluorescence microscopy. Man. Aquat. Viral Ecol. 2010, 145–153. [Google Scholar] [CrossRef]
- Vives-Rego, J.; Lebaron, P. Caron nebe-von current and future applications of £ ow cytometry in aquatic microbiology. FEMS Microbiol. Rev. 2000, 24, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Beuret, C. Simultaneous detection of enteric viruses by multiplex real-time RT-PCR. J. Virol. Methods 2004, 115, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, D.R.; Schwab, K.J.; Cole, R.N.; Halden, R.U. Detection of norovirus capsid protein in authentic standards and in stool extracts by matrix-assisted laser desorption ionization and nanospray mass spectrometry. Appl. Environ. Microbiol. 2006, 72, 2749–2755. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Osborn, A.M. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol. Ecol. 2009, 67, 6–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watzinger, F.; Ebner, K.; Lion, T. Detection and monitoring of virus infections by real-time PCR. Mol. Asp. Med. 2006, 27, 254–298. [Google Scholar] [CrossRef] [PubMed]
- Tekere, M.; Lötter, A.; Olivier, J.; Jonker, N.; Venter, S. Memory tekere metagenomic analysis of bacterial diversity of siloam hot water spring, Limpopo, South Africa. Afr. J. Biotechnol. 2011, 10, 18005–18012. [Google Scholar]
- Kim, J.M.; Kim, S.Y.; Park, Y.B.; Kim, H.J.; Min, B.S.; Cho, J.C.; Yang, J.M.; Cho, Y.H.; Ko, G.P. Simultaneous detection of major enteric viruses using a combimatrix microarray. J. Microbiol. 2012, 50, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Coscoy, L.; Zylberberg, M.; Avila, P.C.; Boushey, H.A.; Ganem, D.; DeRisi, J.L. Microarray-based detection and genotyping of viral pathogens. Proc. Natl. Acad. Sci. USA 2002, 99, 15687–15692. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.A.; De Los Dolores Soto-Del Río, M.; Gutiérrez, R.M.; Chiu, C.Y.; Greninger, A.L.; Contreras, J.F.; López, S.; Arias, C.F.; Isa, P. DNA microarray for detection of gastrointestinal viruses. J. Clin. Microbiol. 2015, 53, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Sano, D.; Tazawa, M.; Inaba, M.; Kadoya, S.; Watanabe, R.; Miura, T.; Kitajima, M.; Okabe, S. Selection of cellular genetic markers for the detection of infectious poliovirus. J. Appl. Microbiol. 2018, 124, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-Y.; Lauder, H.; Cruwys, H.; Falletta, P.; Beaudette, L.A. Development and application of an oligonucleotide microarray and real-time quantitative PCR for detection of wastewater bacterial pathogens. Sci. Total Environ. 2008, 398, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Sidoti, F.; Bergallo, M.; Costa, C.; Cavallo, R. Alternative molecular tests for virological diagnosis. Mol. Biotechnol. 2013, 53, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Compton, J. Nucleic acid sequence-based amplification. Nature 1991, 350, 91–92. [Google Scholar] [CrossRef] [PubMed]
- Starkey, W.G.; Millar, R.M.; Jenkins, M.E.; Ireland, J.H.; Muir, K.F.; Richards, R.H. Detection of piscine nodaviruses by real-time nucleic acid sequence based amplification (NASBA). Dis. Aquat. Organ. 2004, 59, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutjes, S.A.; Van Den Berg, H.H.J.L.; Lodder, W.J.; De Roda Husman, A.M. Real-time detection of noroviruses in surface water by use of a broadly reactive nucleic acid sequence-based amplification assay. Appl. Environ. Microbiol. 2006, 72, 5349–5358. [Google Scholar] [CrossRef] [PubMed]
- Saeedinia, A.; Shamsara, M.; Zeinoddini, M.; Sadeghi, V. Evaluation of nucleic acid sequence based amplification (NASBA) and reverse transcription polymerase chain reaction for detection of coxsackievirus B3 in cell culture and animal tissue samples. Iranian J. Biotechnol. 2008, 6, 222–228. [Google Scholar]
- Jean, J.; Blais, B.; Darveau, A.; Fliss, I. Simultaneous detection and identification of hepatitis A virus and rotavirus by multiplex nucleic acid sequence-based amplification (NASBA) and microtiter plate hybridization system. J. Virol. Methods 2002, 105, 123–132. [Google Scholar] [CrossRef]
- Fakruddin, M.D.; Mazumdar, R.M.; Chowdhury, A.; Mannan, K.S. Bin nucleic acid sequence based amplification (NASBA)—Prospect and applications. Microbiology 2012, 2, 106–121. [Google Scholar]
- Law, J.W.F.; Mutalib, N.S.A.; Chan, K.G.; Lee, L.H. Rapid metho ds for the detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2014, 5, 770. [Google Scholar] [PubMed]
- Kittigul, L.; Khamoun, P.; Sujirarat, D.; Utrarachkij, F.; Chitpirom, K.; Chaichantanakit, N.; Vathanophas, K. An improved method for concentrating rotavirus from water samples. Mem. Inst. Oswaldo Cruz 2001, 96, 815–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koedrith, P.; Thasiphu, T.; Weon, J.I.L.; Boonprasert, R.; Tuitemwong, K.; Tuitemwong, P. Recent trends in rapid environmental monitoring of pathogens and toxicants: Potential of nanoparticle-based biosensor and applications. Sci. World J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Shruthi, G.S.; Amitha, C.V.; Mathew, B.B. Biosensors: A modern day achievement. J. Instrum. Technol. 2014, 2, 26–39. [Google Scholar]
- Meng, J.; Doyle, M.P. Introduction. Microbiological food safety. Microbes Infect. 2002, 4, 395–397. [Google Scholar] [CrossRef]
- Connelly, J.T.; Baeumner, A.J. Biosensors for the detection of waterborne pathogens. Anal. Bioanal. Chem. 2012, 402, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Lazcka, O.; Del Campo, F.J.; Muñoz, F.X. Pathogen detection: A perspective of traditional methods and biosensors. Biosens. Bioelectron. 2007, 22, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Metzker, M.L. Sequencing technologies the next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Salamon, P.; Andresen, B.; Mahaffy, J.M.; Segall, A.M.; Mead, D.; Azam, F.; Rohwer, F. Genomic analysis of uncultured marine viral communities. Proc. Natl. Acad. Sci. USA 2002, 99, 14250–14255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-S.; Whon, T.W.; Bae, J.-W. Comparative viral metagenomics of environmental samples from Korea. Genom. Inf. 2013, 11, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Kramna, L.; Autio, R.; Hyöty, H.; Nykter, M.; Cinek, O. Vipie: Web pipeline for parallel characterization of viral populations from multiple NGS samples. BMC Genom. 2017, 18, 378. [Google Scholar] [CrossRef] [PubMed]
- Rastrojo, A.; Alcamí, A. Aquatic viral metagenomics: Lights and shadows. Virus Res. 2016, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bikel, S.; Valdez-Lara, A.; Cornejo-Granados, F.; Rico, K.; Canizales-Quinteros, S.; Soberón, X.; Del Pozo-Yauner, L.; Ochoa-Leyva, A. Combining metagenomics, metatranscriptomics and viromics to explore novel microbial interactions: Towards a systems-level understanding of human microbiome. Comput. Struct. Biotechnol. J. 2015, 13, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.F.; Milos, P.M. The properties and applications of single-molecule DNA sequencing. Genome Biol. 2011, 12, 217. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, T.J. An introduction to the analysis of shotgun metagenomic data. Front. Plant Sci. 2014, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.; Mahony, J.; Nauta, A.; Van Sinderen, D. Metagenomic approaches to assess bacteriophages in various environmental niches. Viruses 2017, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.; Nakyazze, J.; Wu, H.; Kiwanuka, N.; Cunningham, W.; Kaneene, J.B.; Xagoraraki, I. Viral diversity and abundance in polluted waters in Kampala, Uganda. Water Res. 2017, 127, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, D.M.; Mushegian, A.R.; Dolja, V.V.; Koonin, E.V. New dimensions of the virus world discovered through metagenomics. Trends Microbiol. 2010, 18, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.; Park, S.Y.; Park, M.; Lee, S.; Jo, Y.; Cho, W.K.; Lee, T.K. Metagenomic characterization of viral communities in Goseong Bay, Korea. Ocean Sci. J. 2016, 51, 599–612. [Google Scholar] [CrossRef]
- Heather, J.M.; Chain, B. The sequence of sequencers: The history of sequencing DNA. Genomics 2016, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S. DNA sequencing with chain-terminating. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chiodini, R.; Badr, A.; Zhang, G. The impact of next-generation sequencing on genomics. J. Genet. Genom. 2011, 38, 95–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogorzaly, L.; Walczak, C.; Galloux, M.; Etienne, S.; Gassilloud, B.; Cauchie, H.M. Human adenovirus diversity in water samples using a next-generation amplicon sequencing approach. Food Environ. Virol. 2015, 7, 112–121. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, E.L.; Auger, H.; Jaszczyszyn, Y.; Thermes, C. Ten years of next-generation sequencing technology. Trends Genet. 2014, 30, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K. Single-molecule DNA sequencing technologies for future genomics research. Trends Biotechnol. 2008, 26, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Djikeng, A.; Kuzmickas, R.; Anderson, N.G.; Spiro, D.J. Metagenomic analysis of RNA viruses in a fresh water lake. PLoS ONE 2009, 4, e7264. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Nilsson, C.; Lim, Y.W.; Ruan, Y.; Breitbart, M. Metagenomic analysis of viruses in reclaimed water. Environ. Microbiol. 2009, 11, 2806–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooks, D.J.; Smith, D.L.; Mcdonald, J.E.; Woodward, M.J.; Mccarthy, A.J.; Allison, H.E. 454-pyrosequencing: A molecular battiscope for freshwater viral ecology. Genes 2010, 1, 210–226. [Google Scholar] [CrossRef] [PubMed]
- Neill, J.D.; Bayles, D.O.; Ridpath, J.F. Simultaneous rapid sequencing of multiple RNA virus genomes. J. Virol. Methods 2014, 201, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Lawrence, J.R.; Sanschagrin, S.; Waiser, M.J.; Korber, D.R.; Greer, C.W. Next-generation sequencing of microbial communities in the athabasca river and its tributaries in relation to oil sands mining activities. Appl. Environ. Microbiol. 2012, 78, 7626–7637. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Li, S.; Hu, N.; He, Y.; Pong, R.; Lin, D.; Lu, L.; Law, M. Comparison of next-generation sequencing systems. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hjelmsø, M.H.; Hellmér, M.; Fernandez-Cassi, X.; Timoneda, N.; Lukjancenko, O.; Seidel, M.; Elsässer, D.; Aarestrup, F.M.; Löfström, C.; Bofill-Mas, S.; et al. Evaluation of methods for the concentration and extraction of viruses from sewage in the context of metagenomic sequencing. PLoS ONE 2017, 12, e0170199. [Google Scholar] [CrossRef] [PubMed]
- Iaconelli, M.; Valdazo-González, B.; Equestre, M.; Ciccaglione, A.R.; Marcantonio, C.; Della Libera, S.; La Rosa, G. Molecular characterization of human adenoviruses in urban wastewaters using next generation and Sanger sequencing. Water Res. 2017, 121, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Bayley, H. Nanopore sequencing: From imagination to reality. Clin. Chem. 2015, 61, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Bleidorn, C. Third generation sequencing: Technology and its potential impact on evolutionary biodiversity research. Syst. Biodivers. 2016, 14, 1–8. [Google Scholar] [CrossRef]
- Fichot, E.B.; Norman, R.S. Microbial phylogenetic profiling with the Pacific Biosciences sequencing platform. Microbiome 2013, 1, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheval, J.; Sauvage, V.; Frangeul, L.; Dacheux, L.; Guigon, G.; Dumey, N.; Pariente, K.; Rousseaux, C.; Dorange, F.; Berthet, N.; et al. Evaluation of high-throughput sequencing for identifying known and unknown viruses in biological samples. J. Clin. Microbiol. 2011, 49, 3268–3275. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, Z.Q.; Zhang, Q.; Jin, M.; Yu, J.M.; Li, J.S.; Liu, N.; Cui, S.X.; Kong, X.Y.; Wang, H.; et al. Simultaneous detection of seven enteric viruses associated with acute gastroenteritis by a multiplexed luminex-based assay. J. Clin. Microbiol. 2012, 50, 2384–2389. [Google Scholar] [CrossRef] [PubMed]
- Barba, M.; Czosnek, H.; Hadidi, A. Historical perspective, development and applications of next-generation sequencing in plant virology. Viruses 2013, 6, 106–136. [Google Scholar] [CrossRef] [PubMed]
- Rosario, K.; Breitbart, M. Exploring the viral world through metagenomics. Curr. Opin. Virol. 2011, 1, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Aw, T.G.; Teal, T.K.; Rose, J.B. Metagenomic investigation of viral communities in ballast water. Environ. Sci. Technol. 2015, 49, 8396–8407. [Google Scholar] [CrossRef] [PubMed]
- Angly, F.E.; Felts, B.; Breitbart, M.; Salamon, P.; Edwards, R.A.; Carlson, C.; Chan, A.M.; Haynes, M.; Kelley, S.; Liu, H.; et al. The marine viromes of four oceanic regions. PLoS Biol. 2006, 4, e368. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Zhang, R.; He, Y.; Feng, X.; Jiao, N. Metagenomic analysis of Virioplankton of the subtropical Jiulong river estuary, China. Viruses 2016, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, H.; Zhang, R.; Angly, F.E.; Nakamura, S.; Hong, P.Y.; Yasunaga, T.; Kamagata, Y.; Liu, W.T. Metagenomic analysis of DNA viruses in a wastewater treatment plant in tropical climate. Environ. Microbiol. 2012, 14, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Bibby, K.; Viau, E.; Peccia, J. Viral metagenome analysis to guide human pathogen monitoring in environmental samples. Lett. Appl. Microbiol. 2011, 52, 386–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whon, T.W.; Kim, M.-S.; Roh, S.W.; Shin, N.-R.; Lee, H.-W.; Bae, J.-W. Metagenomic characterization of airborne viral DNA diversity in the near-surface atmosphere. J. Virol. 2012, 86, 8221–8231. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, B.; Freeborough, M.J.; Maree, H.J.; Celton, J.M.; Rees, D.J.G.; Burger, J.T. Deep sequencing analysis of viruses infecting grapevines: Virome of a vineyard. Virology 2010, 400, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M. Environmental viral metagenomics analyses in aquaculture: Applications in epidemiology and disease control. Front. Microbiol. 2016, 7, 1986. [Google Scholar] [CrossRef] [PubMed]
- Breitbart, M.; Hewson, I.; Felts, B.; Mahaffy, J.M.; Nulton, J.; Salamon, P.; Rohwer, F. Metagenomic analyses of an uncultured viral community from human feces metagenomic analyses of an uncultured viral community from human feces. J. Bacteriol. 2003, 185, 6220–6223. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.S.; Kapoor, A.; Lukashov, V.V.; Simmonds, P.; Hecht, F.; Delwart, E. New DNA viruses identified in patients with acute viral infection syndrome. J. Virol. 2005, 79, 8230–8236. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.F.; Suedmeyer, W.K.; Wheeler, E.; Gulland, F.; Breitbart, M. Novel anellovirus discovered from a mortality event of captive California sea lions. J. Gen. Virol. 2009, 90, 1256–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramganesh, S.; Maredza, A.T.; Tekere, M. Microbial Exploration in Extreme Conditions: Metagenomic Analysis and Future Perspectives. In Metagenomics—Methods, Applications and Perspectives; Benedetti, C., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2014; pp. 157–181. [Google Scholar]
- Tekere, M.; Lötter, A.; Olivier, J.; Venter, S. Bacterial diversity in some South African Thermal Springs: A metagenomic analysis. Proc. World Geotherm. Congr. 2015, 19–25. [Google Scholar]
- Selvarajan, R.; Sibanda, T.; Tekere, M. Thermophilic bacterial communities inhabiting the microbial mats of “indifferent” and chalybeate (iron-rich) thermal springs: Diversity and biotechnological analysis. MicrobiologyOpen 2018. [Google Scholar] [CrossRef] [PubMed]
- Selvarajan, R.; Sibanda, T.; Tekere, M.; Nyoni, H.; Meddows-Taylor, S. Diversity analysis and bioresource characterization of halophilic bacteria isolated from a South African saltpan. Molecules 2017, 22, 657. [Google Scholar] [CrossRef] [PubMed]
- Abia, A.L.K.; Alisoltani, A.; Keshri, J.; Ubomba-Jaswa, E. Metagenomic analysis of the bacterial communities and their functional profiles in water and sediments of the Apies River, South Africa, as a function of land use. Sci. Total Environ. 2018, 616–617, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Ramganesh, S.; Timothy, S.; Venkatachalam, S.; Kamika, I.; Nel, W.A.J. Industrial wastewaters harbor a unique diversity of bacterial communities revealed by high-throughput amplicon analysis. Ann. Microbiol. 2018. [Google Scholar] [CrossRef]
- Atoni, E.; Wang, Y.; Karungu, S.; Waruhiu, C.; Zohaib, A.; Obanda, V.; Agwanda, B.; Mutua, M.; Xia, H.; Yuan, Z. Metagenomic virome analysis of Culex mosquitoes from Kenya and China. Viruses 2018, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Vikram, S.; Guerrero, L.D.; Makhalanyane, T.P.; Le, P.T.; Seely, M.; Cowan, D.A. Metagenomic analysis provides insights into functional capacity in a hyperarid desert soil niche community. Environ. Microbiol. 2016, 18, 1875–1888. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.E.; Radosevich, M.; Smith, D.W.; Wommack, K.E. Incidence of lysogeny within temperate and extreme soil environments. Environ. Microbiol. 2007, 9, 2563–2574. [Google Scholar] [CrossRef] [PubMed]
- Le, P.T.; Makhalanyane, T.P.; Guerrero, L.D.; Vikram, S.; Van De Peer, Y.; Cowan, D.A. Comparative metagenomic analysis reveals mechanisms for stress response in hypoliths from extreme hyperarid deserts. Genome Biol. Evol. 2016, 8, 2737–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zablocki, O.; Adriaenssens, E.M.; Cowan, D. Diversity and ecology of viruses in hyperarid desert soils. Appl. Environ. Microbiol. 2015, 82, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Segobola, J.; Adriaenssens, E.; Tsekoa, T.; Rashamuse, K.; Cowan, D. Exploring viral diversity in a unique South African soil habitat. Sci. Rep. 2018, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Zablocki, O. Diversity of dsDNA viruses in a South African hot spring assessed by metagenomics and microscopy. Viruses 2017, 9, 348. [Google Scholar] [CrossRef] [PubMed]
- Vos, H.J.; Knox, C.M. The recovery and molecular identification of HAdV-D17 in raw sewage and mussel samples collected in the Eastern Cape province of South Africa. South. Afr. J. Infect. Dis. 2017, 33, 4–7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Collective Names | Adverse Effects | References |
|---|---|---|---|---|
| Picornaviridae | Enterovirus (ssRNA) Hepatovirus, Hepevirus, Sapovirus (ssRNA) | Poliovirus, Echovirus, Coxsackievirus A, B Hepatitis A, E Sapporo-like virus | Meningitis, Paralysis, Myocarditis, respiratory infections, gastroenteritis Infectious Hepatitis | [51,52,62,64,65,66,67] |
| Reoviridae | Rotaviridae (dsRNA) | Human rotavirus | Gastroenteritis | [51,52,62,64,65,66,67] |
| Adenoviridae | Mastadenovirus (dsDNA) | Human Adenovirus | Conjunctivitis Gastroenteritis Respiratory diseases | [51,52,62,64,65,66,67] |
| Caliciviridae | Calicivirus (ssRNA) Polyomavirus (dsDNA) | Human calicivirus Norwalk virus Polyomavirus | Gastroenteritis, Fever Progressive Multifocal leukoencephalopathy, Urinary tract diseases | [51,52,62,64,65,66,67] |
| Astroviridae | Mamastrovirus Parovirus | Human astrovirus Human parvovirus | Gastroenteritis | [51,52,62,64,65,66,67] |
| Coronoviridae | Coronavirus (ssRNA) | Human coronavirus | Gastroenteritis Respiratory diseases | [51,52,62,64,65,66,67] |
| Circovirus | Torovirus (ssDNA) | Human Torovirus | Gastroenteritis | [51,52,62,64,65,66,67] |
| Virus | Technique | Advantages | Disadvantages | References |
|---|---|---|---|---|
| Enterovirus | Membrane adsorption technique | Simple, speed, sensitive | Low efficiency of virus adsorption Easy clogging of membrane filter | [89] |
| Enterovirus | Aqueous polymer two phase separation | Simple and cost effective Requires small sample volumes | Limited serotypes identified Inhibitory action of salts | [82,89,90] |
| Poliovirus, Herpesvirus, Echovirus | Adsorption to precipitable salts, iron oxide, and polyelectrolytes | Requires large sample volume, simple, time effective | Specific to certain viruses and water samples | [81,82] |
| Poliovirus | Soluble alginate filter | Simple, Non-cytotoxic | Clogging of filters, Pre-filtration required, time consuming | [81,82,89] |
| Poliovirus | Continuous-flow ultracentrifugation | Opportunity for diversity | Expensive instrumentation, time consuming | [81,82,89] |
| Bacteriophage | Forced-flow electrophoresis and electro-osmosis | Small sample volumes, less processing time | Small sample volume | [82,89] |
| Enterovirus | Hydro extraction | Good recoveries | Small sample volume | [82,89] |
| Poliovirus | Gauze sampler | Large sample volume, cost effective | Low efficiency Minimal recovery of viruses | [82,89] |
| Poliovirus, Norovirus, Enterovirus | Electropositive Filtration | Large sample volume, pre-conditioning step not required, Cost effective | Not effective for selected environmental samples including marine water and sediments, expensive | [82,83,91,92] |
| Poliovirus, Echovirus, Reovirus, Coxsackievirus | Electronegative Filtration | Varieties of adsorbent materials, available, High recoveries | Conditioning of large volumes of water is difficult, Acidification protocol may lead to the formation of precipitates, Filter clogs easily, Expensive | [80,82] |
| Poliovirus, Enterovirus, Rotavirus | Glass wool | Less expensive, Pre-conditioning of water sample is not required | Not suitable for large sample volume | [70,71,93,94] |
| Poliovirus, Echovirus, Hepatitis A | Ultrafiltration (Tangential flow, Dead-end flow, Vortex) | No pre-conditioning steps required | Expensive, retreatment of fibres important | [82,95,96] |
| Calcivirus, Hepatitis A | Ultracentrifugation | Less time consuming, Large volumes of water are concentrated to millilitres | Clarification step required, Loss of viruses through the use of membrane filters, expensive | [82,89,97] |
| HEV | Primers and Labelled TaqMan Probes | Target Region | References |
|---|---|---|---|
| Hepatitis A virus | HAV68 (F): 5′-TCA CCG CCG TTT GCC TAG-3′ HAV240 (R): 5′-GGA GAG CCC TGG AAG AAA G3′ HAV150 (P): 5′-FAM-CCT GAA CCT GCA GGA ATT AA-MGBNFQ-3′ | capsid gene VP1/P2B | [69,73,116,122,123] |
| Enterovirus | EV1 (F): 5′-CCCTGAATGCGGCTAAT-3′ EV1 (R): 5′-TGTCACCATA AGCAGCCA-3′ EV-BHQ (P): 5′-FAM- ACGGACACCCAAAGTAGTCGGTTC-MGBNFQ-3 | 5′ Non-coding region | [57,58,69,73,124,125] |
| Rotavirus | JVK (F): 5′-CAGTGGTTGATGCTCAAGATGGA-3′ JVK (R): 5′-TCATTGTAATCATATTGAATACCCA-3′ JVK (P): 5′-FAM-ACAACTGCAGCTTCAAAAGAAGWGT-MGBNFQ-3′ | NSP3 gene | [69,73,126] |
| Noroviruses GI GII | JV13I (F) 5′-TCA TCA TCA CCA TAG AAI GAG-3′ JV12Y (R) 5′-ATA CCA CTA TGA TGC AGA YTA-3′ JV13I (F) 5′-TCA TCA TCA CCA TAG AAI GAG-3′ G1 (R) 5′-TCN GAA ATG GAT GTT GG-3′ JV12Y (F) 5′-ATA CCA CTA TGA TGC AGA YTA-3′ Noro11(R) 5′-AGC CAG TGG GCG ATG GAA TTC-3′ | Polymerase region | [73,127] |
| Adenoviruses | JTVX(F) 5′-GGACGCCTCGGAGTACCTGAG-3′ JTVX(R) 5′-ACIGTGGGGTTTCTGAACTTGTT-3′ JTVX(P):5′-FAM-CTGGTGCAGTTCGCCCGTGCCA-MGBFQ-3′ | Hexon gene | [58,128,129] |
| Astrovirus | HAst.(F): TCAACGTGTCCGTAAMATTGTCA HAstV. (R):TGCWGGTTTTGGTCCTGTGA HAstV.probe1(FAM): CAACTCAGGAAACAGG HAstV.probe2 (FAM): CAACTCAGGAAACAAG | ORF 1b-VPg region ssRNA | [130] |
| Sapovirus GI, II and IV | Sapo (F) A: ACCAGGCTCTCGCCACCTA Sapo (F) B: ATTTGGCCCTCGCCACCTA Sapo (R): GCCCTCCATYTCAAACACTAWTTT Sapo.probeA (FAM) CTGTACCACCTATGAACCA Sapo.probeB (FAM) TTGTACCACCTATGAACCA Sapo.probe C (FAM) TGTACCACCTATAAACCA Sapo.probe D (FAM) TGCACCACCTATGAAC | RdRp-VP1 region | [130,131] |
| Salivirus | F: 5′-TCTGCTTGGTGCCAACCTC-3′ R: 5′-CCARGCACACACATGAGRGGATAC-3′ Probe: 5′-FAM- TGCGGGAGTGCTCTMGB- NFQ-3′ | VP1 region or 3CD region | [132,133] |
| Klassevirus | KLA-F; 5′-TCTGCT TGGTGCCAACCTC-3′ KLA-R; 5′-CCARGC ACACACATGAGRGGATAC-3′ KLA-TP; 5′FAM-TGCGGGAGTGCTCT-MGB-NFQ-3′ | VP0/VP3 regions | [133] |
| Human Parechovirus | F: 5′-CCA AAA TTC RTG GGG TTC-3′ R: 5′-AAA CCY CTR TCT AAA TAW GC-3′ | VP1 capsid gene or 3CD region | [134,135] |
| Aichi virus | F: ACA CTC CCA CCT CCAGCC AGT A R: GGA AGA GCT GGG TGT CAA GA | 3CD junction region | [134,135] |
| Technique | Principle | Advantage | Disadvantages | References |
|---|---|---|---|---|
| Cell culture | Cytopathic effects potential for viruses | Direct isolation of a variety of cultivable viruses to high titres | Highly skilled Requires controlled conditions Expensive and time consuming | [51,99,105] |
| Electron microscope transmission electron microscopy | Electron beam used to illuminate viruses. Counting of the viral particles and morphology | Prior knowledge of organism not required DNA provides high resolution image | It requires technical skills and expertise Poor detection limit High concentrations High cost of maintenance and training of the instrument | [97,109,110,143] |
| Flow cytometry | Direct and rapid assays for the determination of cell numbers and morphology | High speed and velocity | Skill generation and refrigeration a pre-requisite, expensive | [112,144] |
| Vortex flow filtration | Counting and quantifying virus-like particles | High recovery Reduces filter clogging | Expensive method | [107,112,144] |
| PCR Assay | Amplification assays based on specific primers and enzyme to generate more copies of DNA | Sequence dependent Cost effective High sensitivity and specificity | Cannot detect new viral species Risk of contamination False positive results | [51,66,106] |
| ICC-PCR | Viral particle is amplified via host cell assays | Less vulnerable to PCR inhibition Identify non-cytopathic viruses | Does not detect non-culturable viruses, Requires multiple cell lines Time consuming, More costly than direct PCR detection | [51,66,105,106] |
| Multiple PCR | Simultaneous amplification of sequences of several pathogenic microorganisms in a reaction mixture | Sequence dependent, Cost and time effective, High sensitivity and specific | Cannot detect new viral species Challenges with optimisation and sensitivity for all targeted species Contamination Non-specific amplification in environmental samples | [51,106,136,145] |
| Nested/Semi Nested PCR | Distinct pair of primers amplifies enormous region of DNA The amplified PCR product is now used as a template for the next round of amplification | Increased sensitivity | Potential risk of contamination and carry-over | [51,106,120,146] |
| RT-PCR (Reverse-transcriptase PCR) | Amplification is achieved by converting DNA to complementary DNA (cDNA) in a reverse transcription procedure | Speed sensitivity Contamination Specificity Repeatability | Sequence knowledge is a perquisite, expensive, Possible reaction inhibition, and there is a need for experts for the interpretation as well as the accuracy of results | [51,66,78,106,137,147,148,149] |
| qRT-PCR (quantitative real-time PCR) | Quantifies and measures amplification of DNA using dyes or fluorescent dyes or probes | Elimination of gel electrophoresis applicable for both culturable and unculturable microorganisms | ||
| Microarray technology | Detection is done by means of radio-labelled probes or fluorescent tags | Known viral sequences | Expensive Reproducibility test results are poor | [150,151,152,153,154] |
| NASBA | Isothermal amplification of RNA | Sensitive, rapid simple Resistant to matrix influence | Can be used only for organisms, which are already known | [106,155,156,157,158,159,160,161] |
| Immunology-based method | Formation of antigen—antibody through recognition and binding | High sensitivity Specificity speed Easy automation and equipment | QC assurance dependent Risk of interferences Expensive | [162,163] |
| Biosensor-based methods | analytical device that identifies analytes via an electrical signal | Detects non-polar molecules High specificity Reaction time is short | Relies on specific antibodies or DNA Probes Necessary chemical inactivation for the recognition sites | [164,165,166,167,168] |
| NGS | Parallel sequencing of multiple small fragments of DNA to determine its sequence using high-throughput instrumentation | Fast and easy to approach for DNA sequencing Large sequencing data per run | Expensive equipment | [169] |
| Platform | Amplification Technique | Chemistry | Read Length | Output and Duration | Advantages | Disadvantages | References |
|---|---|---|---|---|---|---|---|
| Roche 454 | Emulsion PCR | Pyro-sequencing | 400–700 bp | 100–700 Mb 10–23 h | Long read length, short run times | High error rate | [22,23,186,196,198,199,200] |
| AB SOLiD | Emulsion PCR | Ligation | 35 bp | 80–360 Gb between 6–8 days | Low error rate | Short reads Long run time | [22,23,185,186,195,196,199] |
| Ion Torrent (PGM) | Emulsion PCR | Proton detection | 100–400 | 100–64 Gb for 2–7 h | Less sequencing time, reduces costs | Short reads Homopolymer errors | [22,186,192] |
| Illumina Solexa (MiSeq, HiSeq) | Bridge PCR | Reversible terminators | 100–300 | 600 Gb 5 h to 3 days run | High throughput, Cost and time effective, minimal error rate | Short reads Decrease in quality of reads towards the ends | [22,186,192] |
| Pacific Bioscience (SMRT) | Single molecule real time (SMRT) | Fluorescently labelled nucleotides | 4000–5000 nts | 200 Mb–1 Gb generated under few hours | Data generation is monitored in real-time, Accurate | Expensive, high error rates | [22,23,186,195,196] |
| Helicos TM Genetic Analysis System | non-amplified DNA templates | Fluorescently labelled nucleotides | 24–70 bp | 35 Gb for a few hours | Accurate | Expensive, low data output | [23,186,196] |
| Oxford Nanopore (MinION) | Single molecule real time (SMRT) | Reversible terminators | 90 Mbp of data with 16,000 reads | 6 kb–60 kb | Accurate | Expensive, high error rate, low throughput | [22,23,186,195,196] |
| Country | Microbe | NGS Platform | Environment | References |
|---|---|---|---|---|
| South Africa | Bacteria thermophiles | Roche 454 | Hot spring | [149] |
| South Africa | Bacteria | Illumina MiSeq | Surface water Sediments, Industrial wastewater | [215,217] |
| Namibia | Virus | Soil, deserts | [220,221] | |
| Kenya | Mosquito | Illumina | Clinical sample | [219] |
| South Africa | Virus | Illumina | Hot spring | [225] |
| Uganda | Viruses (HEV) | Illumina | Surface water, WWTP | [178] |
| South Africa | Viruses (HADV) | Illumina | Sewage Mussels | [226] |
| South Africa | Viruses (Caudovirales, phages) | Illumina MiSeq | Soil | [224] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osunmakinde, C.O.; Selvarajan, R.; Sibanda, T.; Mamba, B.B.; Msagati, T.A.M. Overview of Trends in the Application of Metagenomic Techniques in the Analysis of Human Enteric Viral Diversity in Africa’s Environmental Regimes. Viruses 2018, 10, 429. https://doi.org/10.3390/v10080429
Osunmakinde CO, Selvarajan R, Sibanda T, Mamba BB, Msagati TAM. Overview of Trends in the Application of Metagenomic Techniques in the Analysis of Human Enteric Viral Diversity in Africa’s Environmental Regimes. Viruses. 2018; 10(8):429. https://doi.org/10.3390/v10080429
Chicago/Turabian StyleOsunmakinde, Cecilia Oluseyi, Ramganesh Selvarajan, Timothy Sibanda, Bhekie B Mamba, and Titus A.M Msagati. 2018. "Overview of Trends in the Application of Metagenomic Techniques in the Analysis of Human Enteric Viral Diversity in Africa’s Environmental Regimes" Viruses 10, no. 8: 429. https://doi.org/10.3390/v10080429
APA StyleOsunmakinde, C. O., Selvarajan, R., Sibanda, T., Mamba, B. B., & Msagati, T. A. M. (2018). Overview of Trends in the Application of Metagenomic Techniques in the Analysis of Human Enteric Viral Diversity in Africa’s Environmental Regimes. Viruses, 10(8), 429. https://doi.org/10.3390/v10080429