HSV-Induced Systemic Inflammation as an Animal Model for Behçet’s Disease and Therapeutic Applications



Abstract
:1. Introduction
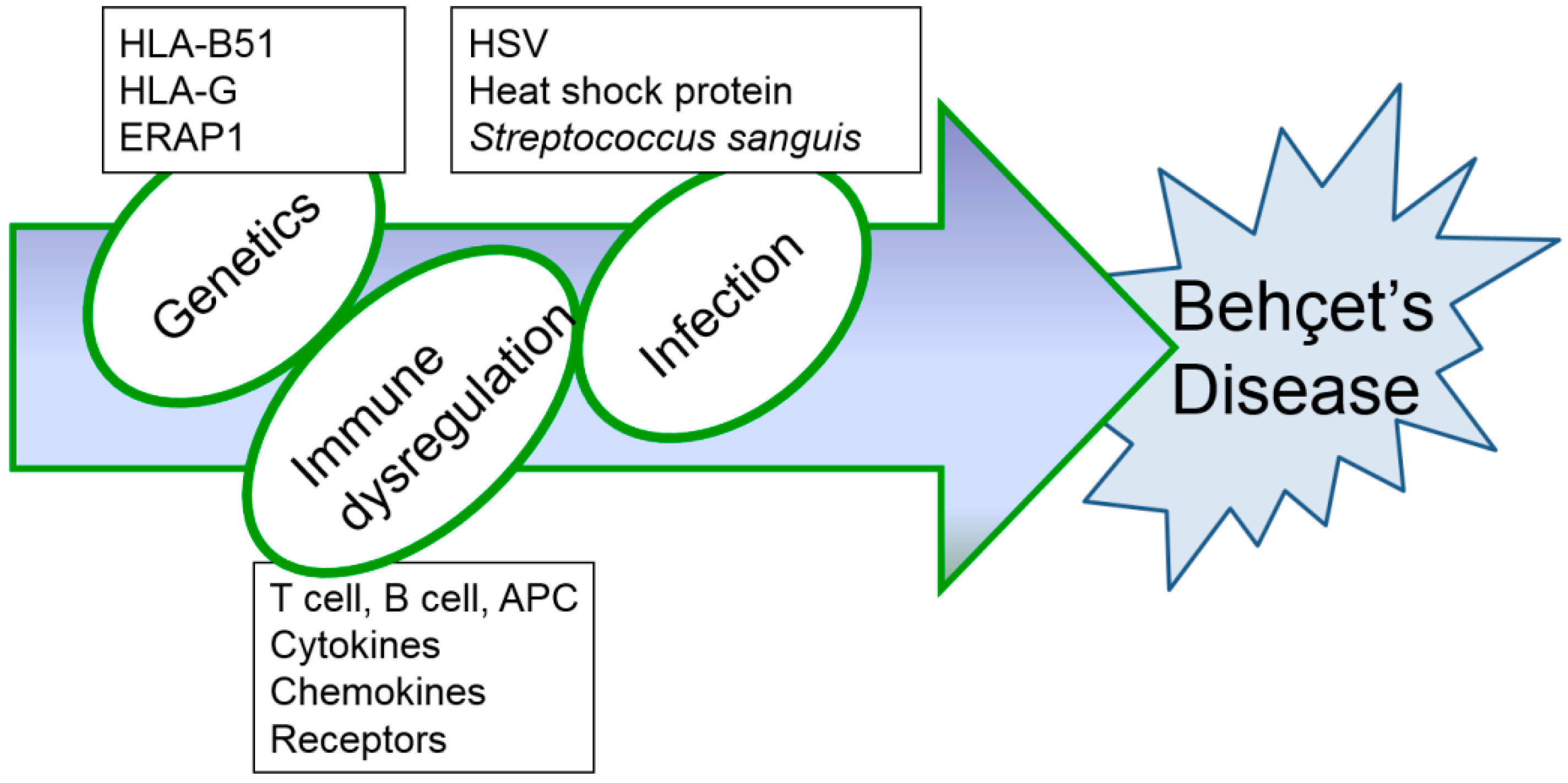
2. Etiopathogenesis of Behçet’s Disease
2.1. Clinical Significance of HSV
2.2. Genetic Susceptibility
2.3. Immuological Dysregulation
3. Therapeutic Applications of Inflammatory Mice Model of Behçet’s Disease Induced by HSV
3.1. Natural Products
3.1.1. TNFα Inhibition with Derivatives of Gentiana Macrophylla Radix
3.1.2. Herba Taraxaci
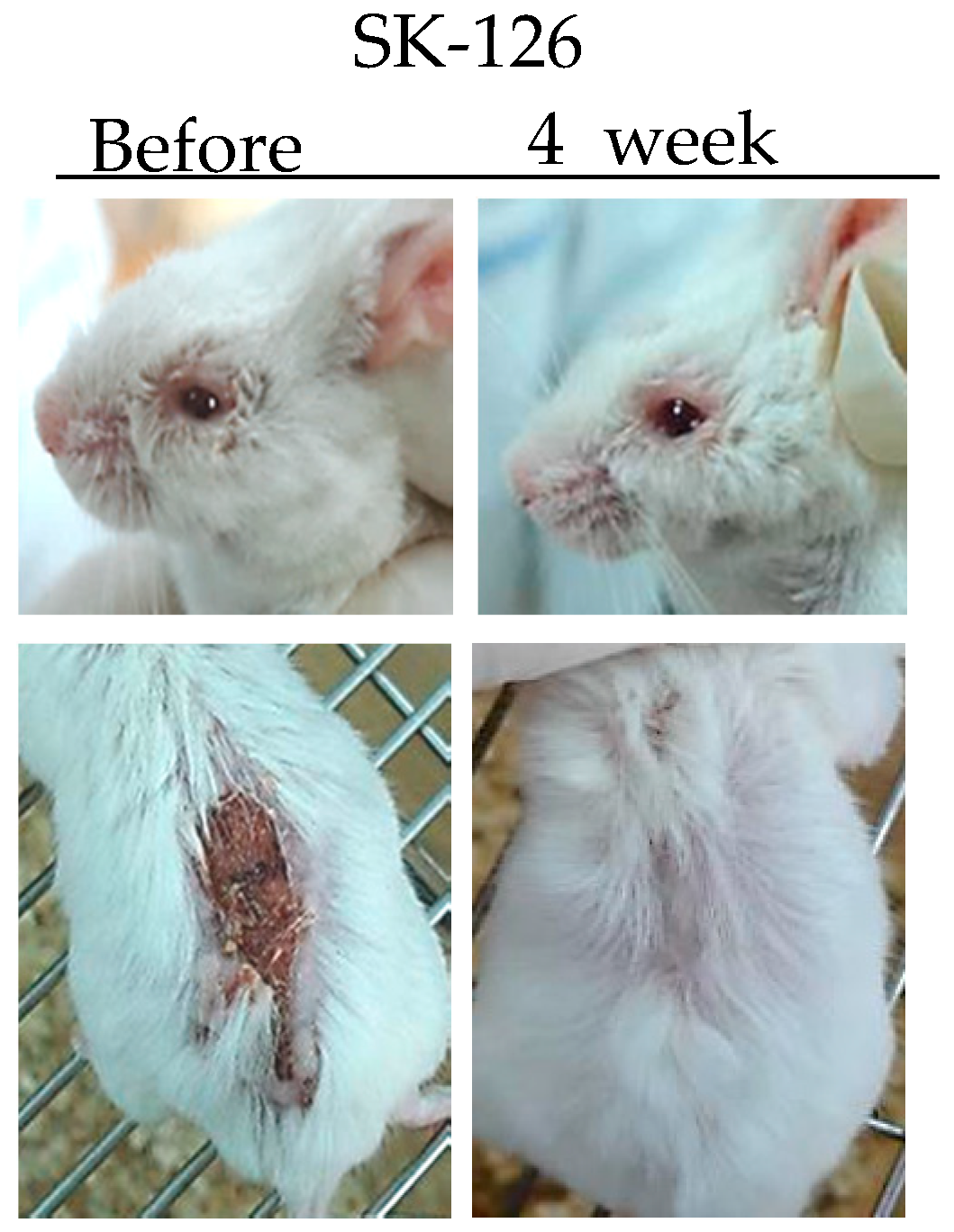
3.1.3. Chitosan
3.2. DNA Vector
3.2.1. Interleukin-4
3.2.2. C-C Chemokine Ligand 21
3.3. RNA and siRNA
3.3.1. Polyinosinic:Polycytidylic Acid (Poly I:C)
3.3.2. TNFα siRNA
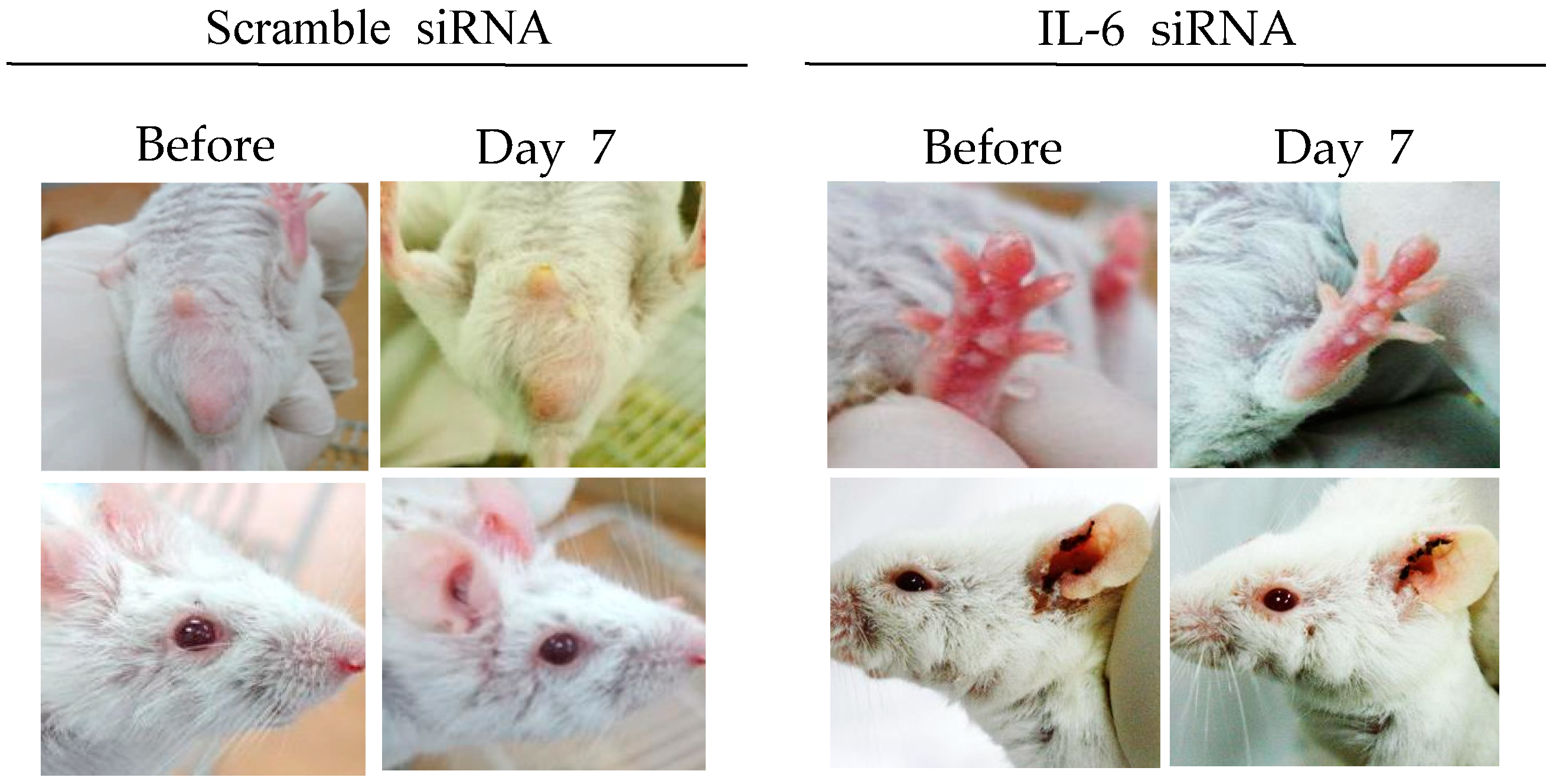
3.3.3. Interleukin-6 siRNA
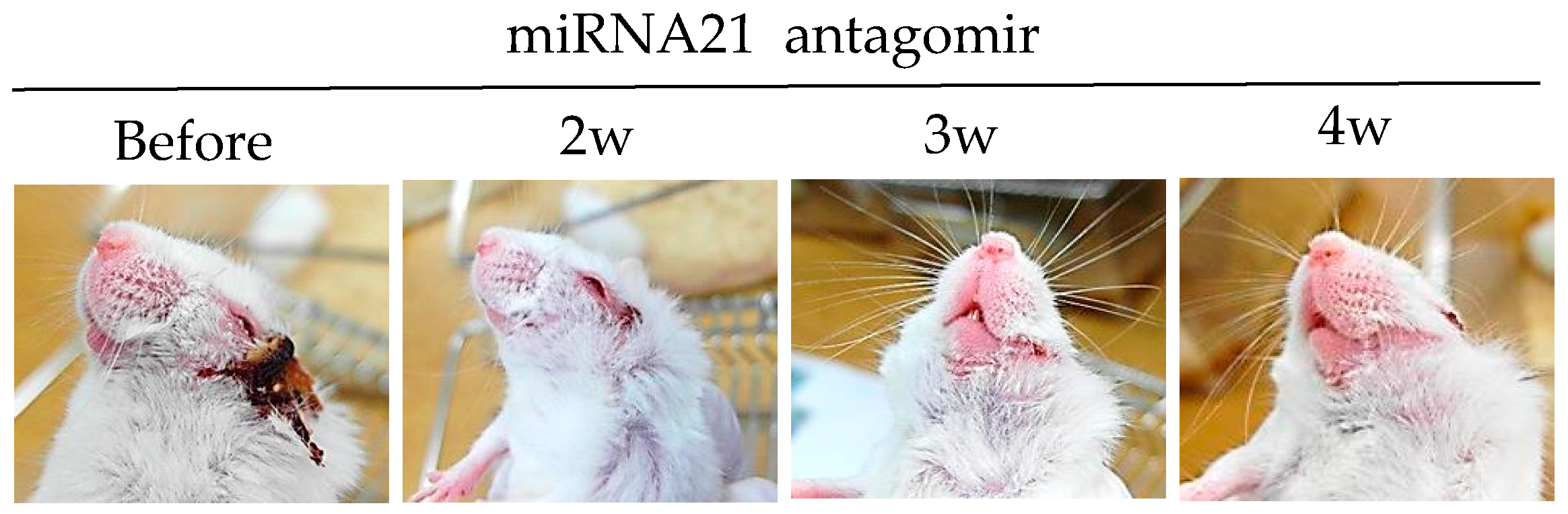
3.3.4. miRNA-21 Antagomir
3.4. Protein Complex
3.4.1. IL-2/IL-2 Antibody Complex
3.4.2. IL-15/IL-15Rα Complex
3.5. Vitamin & Galactose Derivatives
3.5.1. Vitamin D3
3.5.2. N-Acetylgalactosamine (GalNAc), a Mannose Receptor Ligand
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sohn, S.; Lee, E.S.; Bang, D. Learning from HSV-infected mice as a model of Behcet’s disease. Clin. Exp. Rheumatol. 2012, 30, S96–S103. [Google Scholar] [PubMed]
- Greco, A.; De Virgilio, A.; Ralli, M.; Ciofalo, A.; Mancini, P.; Attanasio, G.; de Vincentiis, M.; Lambiase, A. Behçet’s disease: New insights into pathophysiology, clinical features and treatment options. Autoimmun. Rev. 2018, 17, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Eglin, R.P.; Lehner, T.; Subak-Sharpe, J.H. Detection of RNA complementary to herpes-simplex virus in mononuclear cells from patients with Behcet’s syndrome and recurrent oral ulcers. Lancet 1982, 2, 1356–1361. [Google Scholar] [PubMed]
- Evans, A.D.; Pallis, C.A.; Spillane, J.D. Involvement of the nervous system in Behcet’s syndrome; report of three cases and isolation of virus. Lancet 1957, 273, 349–353. [Google Scholar] [PubMed]
- Sezer, F.N. The isolation of a virus as the cause of behçet’s disease. Am. J. Ophthalmol. 1953, 36, 301–315. [Google Scholar] [PubMed]
- Studd, M.; McCance, D.J.; Lehner, T. Detection of HSV-1 DNA in patients with Behcet’s syndrome and in patients with recurrent oral ulcers by the polymerase chain reaction. J. Med. Microbiol. 1991, 34, 39–43. [Google Scholar] [PubMed]
- Sohn, S.; Lee, E.S.; Bang, D.; Lee, S. Behcet’s disease-like symptoms induced by the herpes simplex virus in ICR mice. Eur. J. Dermatol. 1998, 8, 21–23. [Google Scholar] [PubMed]
- Lee, S.; Bang, D.; Cho, Y.H.; Lee, E.S.; Sohn, S. Polymerase chain reaction reveals herpes simplex virus DNA in saliva of patients with Behcet’s disease. Arch. Dermatol. Res. 1996, 288, 179–183. [Google Scholar] [PubMed]
- Tojo, M.; Zheng, X.; Yanagihori, H.; Oyama, N.; Takahashi, K.; Nakamura, K.; Kaneko, F. Detection of herpes virus genomes in skin lesions from patients with Behcet’s disease and other related inflammatory diseases. Acta Derm. Venereol. 2003, 83, 124–127. [Google Scholar] [PubMed]
- Kim, J.E.; Nam, J.H.; Cho, J.Y.; Kim, K.S.; Hwang, D.Y. Annual tendency of research papers used ICR mice as experimental animals in biomedical research fields. Lab. Anim. Res. 2017, 33, 171–178. [Google Scholar] [PubMed]
- Zheng, Z.; Sohn, S.; Ahn, K.J.; Bang, D.; Cho, S.B. Serum reactivity against herpes simplex virus type 1 UL48 protein in Behcet’s disease patients and a Behcet’s disease-like mouse model. Acta Derm. Venereol. 2015, 95, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Seoudi, N.; Bergmeier, L.A.; Hagi-Pavli, E.; Bibby, D.; Fortune, F. The seroprevalence and salivary shedding of herpesviruses in Behcet’s syndrome and recurrent aphthous stomatitis. J. Oral. Microbiol. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Paludan, S.R.; Bowie, A.G.; Horan, K.A.; Fitzgerald, K.A. Recognition of herpesviruses by the innate immune system. Nat. Rev. Immunol. 2011, 11, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Cho, S.; Choi, M.J.; Sohn, S.; Lee, E.S.; Bang, D. Immunopathogenic role of herpes simplex virus in Behcet’s disease. Genet. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Ohguchi, M.; Hirose, S.; Matsuda, H.; Wakisaka, A.; Aizawa, M. Close association of HLA-Bw51 with Behcet’s disease. Arch. Ophthalmol. 1982, 100, 1455–1458. [Google Scholar] [CrossRef] [PubMed]
- Verity, D.H.; Marr, J.E.; Ohno, S.; Wallace, G.R.; Stanford, M.R. Behcet’s disease, the silk road and HLA-B51: Historical and geographical perspectives. Tissue Antigens. 1999, 54, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Takeno, M.; Kariyone, A.; Yamashita, N.; Takiguchi, M.; Mizushima, Y.; Kaneoka, H.; Sakane, T. Excessive function of peripheral blood neutrophils from patients with Behcet’s disease and from HLA-B51 transgenic mice. Arthritis Rheum. 1995, 38, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Nam, J.H.; Lee, E.S.; Choi, J.S.; Bang, D.; Lee, S. Increased risk of human leukocyte antigen-G gene variants in Behçet’s disease. Clin. Exp. Rheumatol. 2006, 24, S126–S127. [Google Scholar] [PubMed]
- Park, K.S.; Park, J.S.; Nam, J.H.; Bang, D.; Sohn, S.; Lee, E.S. HLA-E*0101 and HLA-G*010101 reduce the risk of Behcet’s disease. Tissue Antigens. 2007, 69, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Kovats, S.; Main, E.K.; Librach, C.; Stubblebine, M.; Fisher, S.J.; DeMars, R. A class I antigen, HLA-G, expressed in human trophoblasts. Science 1990, 248, 220–223. [Google Scholar] [CrossRef] [PubMed]
- McMaster, M.T.; Librach, C.L.; Zhou, Y.; Lim, K.H.; Janatpour, M.J.; DeMars, R.; Kovats, S.; Damsky, C.; Fisher, S.J. Human placental HLA-G expression is restricted to differentiated cytotrophoblasts. J. Immunol. 1995, 154, 3771–3778. [Google Scholar] [PubMed]
- Michita, R.T.; Zambra, F.M.B.; Fraga, L.R.; Sanseverino, M.T.V.; Callegari-Jacques, S.M.; Vianna, P.; Chies, J.A.B. A tug-of-war between tolerance and rejection–new evidence for 3′UTR HLA-G haplotypes influence in recurrent pregnancy loss. Hum. Immunol. 2016, 77, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Riteau, B.; Rouas-Freiss, N.; Menier, C.; Paul, P.; Dausset, J.; Carosella, E.D. HLA-G2, -G3, and -G4 isoforms expressed as nonmature cell surface glycoproteins inhibit nk and antigen-specific CTL cytolysis. J. Immunol. 2001, 166, 5018–5026. [Google Scholar] [CrossRef] [PubMed]
- Dorling, A.; Monk, N.J.; Lechler, R.I. HLA-G inhibits the transendothelial migration of human NK cells. Eur. J. Immunol. 2000, 30, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Kanai, T.; Fujii, T.; Unno, N.; Yamashita, T.; Hyodo, H.; Miki, A.; Hamai, Y.; Kozuma, S.; Taketani, Y. Human leukocyte antigen-G-expressing cells differently modulate the release of cytokines from mononuclear cells present in the decidua versus peripheral blood. Am. J. Reprod. Immunol. 2001, 45, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, D.R.; Ellis, S.A.; Sargent, I.L. HLA-G suppresses proliferation of CD4(+) T-lymphocytes. J. Reprod. Immunol. 2000, 48, 17–26. [Google Scholar] [CrossRef]
- Riteau, B.; Menier, C.; Khalil-Daher, I.; Sedlik, C.; Dausset, J.; Rouas-Freiss, N.; Carosella, E.D. HLA-G inhibits the allogeneic proliferative response. J. Reprod. Immunol. 1999, 43, 203–211. [Google Scholar] [CrossRef]
- Comiskey, M.; Goldstein, C.Y.; De Fazio, S.R.; Mammolenti, M.; Newmark, J.A.; Warner, C.M. Evidence that HLA-G is the functional homolog of mouse Qa-2, the ped gene product. Hum. Immunol. 2003, 64, 999–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.; Choi, B.; Kwon, H.J.; Shim, J.A.; Park, K.S.; Lee, E.S.; Sohn, S. The role of Qa-2, the functional homolog of Hla-G, in a Behcet’s disease-like mouse model induced by the herpes virus simplex. J. Inflamm. 2010, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Frassanito, M.A.; Dammacco, R.; Cafforio, P.; Dammacco, F. Th1 polarization of the immune response in Behcet’s disease: A putative pathogenetic role of interleukin-12. Arthritis Rheum. 1999, 42, 1967–1974. [Google Scholar] [CrossRef]
- Raziuddin, S.; al-Dalaan, A.; Bahabri, S.; Siraj, A.K.; al-Sedairy, S. Divergent cytokine production profile in Behcet’s disease. Altered Th1/Th2 cell cytokine pattern. J. Rheumatol. 1998, 25, 329–333. [Google Scholar] [PubMed]
- Desmedt, M.; Rottiers, P.; Dooms, H.; Fiers, W.; Grooten, J. Macrophages induce cellular immunity by activating Th1 cell responses and suppressing Th2 cell responses. J. Immunol. 1998, 160, 5300–5308. [Google Scholar] [PubMed]
- Sohn, S.; Lee, E.-S.; Kwon, H.J.; Lee, S.I.; Bang, D.; Lee, S. Expression of Th2 cytokines decreases the development of and improves Behçet’s disease–like symptoms induced by herpes simplex virus in mice. J. Infect. Dis. 2001, 183, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Nanke, Y.; Kotake, S.; Goto, M.; Ujihara, H.; Matsubara, M.; Kamatani, N. Decreased percentages of regulatory T cells in peripheral blood of patients with Behcet’s disease before ocular attack: A possible predictive marker of ocular attack. Mod. Rheumatol. 2008, 18, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Lee, E.S.; Park, S.; Bang, D.; Sohn, S. CD4(+) CD25(+) regulatory T cells ameliorate Behcet’s disease-like symptoms in a mouse model. Cytotherapy 2011, 13, 835–847. [Google Scholar] [CrossRef] [PubMed]
- El Menyawi, M.; Fawzy, M.; Al-Nahas, Z.; Edris, A.; Hussein, H.; Shaker, O.; Elwan, H. Serum tumor necrosis factor alpha (TNF-α) level in patients with Behçet’s disease: Relation to clinical manifestations and disease activity. Egypt. Rheumatol. 2014, 36, 139–143. [Google Scholar] [CrossRef]
- Mohammed, R.H. Etanercept therapy in Behcet’s disease. The tight control strategy in refractory disease. Z. Rheumatol. 2014, 73, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Ugras, M.; Ertem, D.; Celikel, C.; Pehlivanoglu, E. Infliximab as an alternative treatment for Behçet disease when other therapies fail. J. Pediatr. Gastroenterol. Nutr. 2008, 46, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Hwang, Y.; Kwon, H.J.; Lee, E.S.; Park, K.S.; Bang, D.; Lee, S.; Sohn, S. Tumor necrosis factor alpha small interfering RNA decreases herpes simplex virus-induced inflammation in a mouse model. J. Dermatol. Sci. 2008, 52, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.; Kim, J.; Lee, E.S.; Bang, D.; Sohn, S. Synthesized pyridine compound derivatives decreased TNF alpha and adhesion molecules and ameliorated HSV-induced inflammation in a mouse model. Eur. J. Pharmacol. 2011, 657, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schütz, K.; Carle, R.; Schieber, A. Taraxacum—A review on its phytochemical and pharmacological profile. J. Ethnopharmacol. 2006, 107, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Dai, W.; Yu, H.; Wang, Y.; Wang, X.; Peng, S. Antibacterial activity of aqueous and ethanolic extracts of Portulaca oleracea L. And Taraxacum mongolicum Hand.-Mazz against pathogenic bacteria of cow mastitis. Indian J. Anim. Res. 2015, 49, 827–829. [Google Scholar] [CrossRef]
- Sohn, S.; Bang, D.; Lee, S.I.; Kim, Y.A.; Lee, E.S.; Ha, J.Y.; Kim, J.H.; Choi, S.Y.; Lee, S. Combined treatment with colchicine and Herba Taraxaci (Tarazacum mongolicum Hand.-Mazz.) attenuates Behcet’s disease-like symptoms in mice and influences the expressions of cytokines. Int. Immunopharmacol. 2003, 3, 713–721. [Google Scholar] [CrossRef]
- Nakamura, S.; Sugita, M.; Matoba, H.; Tanaka, S.; Isoda, F.; Ohno, S. Insufficient expression of Fas antigen on helper t cells in behcet’s disease. Br. J. Ophthalmol. 1996, 80, 174–176. [Google Scholar] [CrossRef] [PubMed]
- Carlos, P.; Waldo, A.M.; Hazel, P.; Niuris, A. Chitosan: An attractive biocompatible polymer for microencapsulation. Macromol. Biosci. 2003, 3, 511–520. [Google Scholar]
- Sailaja, A.K.; Amareshwar, P.; Chakravarty, P. Chitosan nanoparticles as a drug delivery system. Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 474–484. [Google Scholar]
- Choi, B.C.; Choi, J.Y.; Sohn, S. Oral delivery of DNA vector conjugated with chitosan and its effect on Th1 polarized inflammation. In Advances in the Diagnosis and Treatemnt of Vasculitis; Amexcua-Guerra, L.M., Ed.; InTech: London, UK, 2011; pp. 221–232. [Google Scholar]
- Choi, B.; Jo, D.H.; Anower, A.K.; Islam, S.M.; Sohn, S. Chitosan as an immunomodulating adjuvant on T-cells and antigen-presenting cells in herpes simplex virus type 1 infection. Mediators Inflamm. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Gratchev, A.; Kzhyshkowska, J.; Kothe, K.; Muller-Molinet, I.; Kannookadan, S.; Utikal, J.; Goerdt, S. Mphi1 and Mphi2 can be re-polarized by Th2 or Th1 cytokines, respectively, and respond to exogenous danger signals. Immunobiology 2006, 211, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Stout, R.D.; Suttles, J. Functional plasticity of macrophages: Reversible adaptation to changing microenvironments. J. Leukoc. Biol. 2004, 76, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Kwon, H.J.; Lee, E.S.; Yang, B.C.; Bang, D.; Lee, S.; Sohn, S. Using pcin-mil-4 DNA vector to express mrna and protein and to improve herpes simplex virus-induced Behcet’s disease symptoms in mice. Vaccine 2007, 25, 7047–7055. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Katayama, N.; Fujieda, A.; Shibasaki, T.; Yamamura, K.; Sugimoto, Y.; Miyata, E.; Ohishi, K.; Nishii, K.; Masuya, M.; et al. IL-4 and IL-10 synergistically inhibit survival of human blood monocytes supported by GM-CSF. Int. J. Oncol. 2005, 26, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Anower, A.K.; Shim, J.A.; Choi, B.; Kwon, H.J.; Sohn, S. The role of classical and alternative macrophages in the immunopathogenesis of herpes simplex virus-induced inflammation in a mouse model. J. Dermatol. Sci. 2014, 73, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Forster, R.; Davalos-Misslitz, A.C.; Rot, A. Ccr7 and its ligands: Balancing immunity and tolerance. Nat. Rev. Immunol. 2008, 8, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Pay, S.; Musabak, U.; Simsek, I.; Erdem, H.; Pekel, A.; Sengul, A.; Dinc, A. Synovial lymphoid neogenetic factors in Behcet’s synovitis: Do they play a role in self-limiting and subacute course of arthritis? Clin. Exp. Rheumatol. 2007, 25, S21–S26. [Google Scholar] [PubMed]
- Choi, B.; Lim, H.C.; Lee, E.S.; Anower, A.K.; Sohn, S. CCL21 attenuates HSV-induced inflammation through up-regulation of CD8+ memory cells. Immunobiology 2013, 218, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Direskeneli, H.; Ergun, T.; Yavuz, S.; Hamuryudan, V.; Eksioglu-Demiralp, E. Thalidomide has both anti-inflammatory and regulatory effects in Behcet’s disease. Clin. Rheumatol. 2008, 27, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Jalili, A.; Pashenkov, M.; Kriehuber, E.; Wagner, C.; Nakano, H.; Stingl, G.; Wagner, S.N. Induction of targeted cell migration by cutaneous administration of a DNA vector encoding a biologically active chemokine CCL21. J. Invest. Dermatol. 2010, 130, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Lodolce, J.P.; Burkett, P.R.; Boone, D.L.; Chien, M.; Ma, A. T cell-independent interleukin 15ralpha signals are required for bystander proliferation. J. Exp. Med. 2001, 194, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, I.; Dingley, A.J.; Jacques, Y.; Grotzinger, J. The structure of the interleukin-15 alpha receptor and its implications for ligand binding. J. Biol. Chem. 2006, 281, 6642–6647. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cella, M.; Gilfillan, S.; Colonna, M. Cutting edge: Polyinosinic:Polycytidylic acid boosts the generation of memory CD8 T cells through melanoma differentiation-associated protein 5 expressed in stromal cells. J. Immunol. 2010, 184, 2751–2755. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, E.S.; Choi, B.; Sohn, S. Therapeutic potency of Poly I:C in HSV-induced inflammation through up-regulation of IL-15 receptor alpha. Immunobiology 2013, 218, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Patel, H.J.; Waldmann, T.A.; Tagaya, Y. The IL-15/IL-15ralpha on cell surfaces enables sustained IL-15 activity and contributes to the long survival of CD8 memory T cells. Proc. Natl. Acad. Sci. USA 2007, 104, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Van Deventer, S.J. Tumour necrosis factor and Crohn’s disease. Gut 1997, 40, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Watanabe, N.; Neda, H.; Yamauchi, N.; Okamoto, T.; Sasaki, H.; Tsuji, Y.; Akiyama, S.; Tsuji, N.; Niitsu, Y. Serum tumor necrosis factor activity in inflammatory bowel disease. Immunopharmacol. Immunotoxicol. 1992, 14, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, H. TNF as a target of inflammation in rheumatoid arthritis. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Sumimoto, S.; Kawai, M.; Kasajima, Y.; Hamamoto, T. Increased plasma tumour necrosis factor-alpha concentration in atopic dermatitis. Arch. Dis. Child. 1992, 67, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Karube, H.; Kamoi, K.; Ohno-Matsui, K. Anti-TNF therapy in the management of ocular attacks in an elderly patient with long-standing Behçet’s disease. Int. Med. Case Rep. J. 2016, 9, 301–304. [Google Scholar] [PubMed]
- Desbois, A.C.; Addimanda, O.; Bertrand, A.; Deroux, A.; Pérard, L.; Depaz, R.; Hachulla, E.; Lambert, M.; Launay, D.; Subran, B.; et al. Efficacy of Anti-TNFα in Severe and Refractory Neuro-Behcet Disease: An Observational Study. Medicine 2016, 95, e3550. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.D.; Vorhies, J.S.; Senzer, N.; Nemunaitis, J. siRNA vs. shRNA: Similarities and differences. Adv. Drug Deliv. Rev. 2009, 61, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Akdeniz, N.; Esrefoglu, M.; Keles, M.S.; Karakuzu, A.; Atasoy, M. Serum interleukin-2, interleukin-6, tumour necrosis factor-alpha and nitric oxide levels in patients with Behcet’s disease. Ann. Acad. Med. Singap. 2004, 33, 596–599. [Google Scholar] [PubMed]
- Heinrich, P.C.; Castell, J.V.; Andus, T. Interleukin-6 and the acute phase response. Biochem. J. 1990, 265, 621–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanangat, S.; Babu, J.S.; Knipe, D.M.; Rouse, B.T. HSV-1-mediated modulation of cytokine gene expression in a permissive cell line: Selective upregulation of IL-6 gene expression. Virology 1996, 219, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Paludan, S.R. Requirements for the induction of interleukin-6 by herpes simplex virus-infected leukocytes. J. Virol. 2001, 75, 8008–8015. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Byun, H.O.; Lee, Y.D.; Lee, E.S.; Sohn, S. Interleukin-6 small interfering rna improved the herpes simplex virus-induced systemic inflammation in vivo Behcet’s disease-like mouse model. Gene Ther. 2009, 16, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, Y.; Sugita, Y.; Nagatani, T.; Takahashi, S.; Yamakawa, T.; Tanaka, S.; Nakamura, S.; Ohno, S.; Sekihara, H.; Okuda, K.; et al. Interleukin-6 (IL-6) in patients with Behcet’s disease. J. Dermatol. Sci. 1996, 11, 189–195. [Google Scholar] [CrossRef]
- Davidson-Moncada, J.; Papavasiliou, F.N.; Tam, W. miRNAs of the immune system: Roles in inflammation and cancer. Ann. N. Y. Acad. Sci. 2010, 1183, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Xiao, X.; Wang, C.; Zhang, X.; Li, F.; Zhou, Y.; Kijlstra, A.; Yang, P. Decreased microRNA-155 expression in ocular Behcet’s disease but not in Vogt Koyanagi Harada syndrome. Invest. Ophthalmol. Vis. Sci. 2012, 53, 5665–5674. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Hou, S.; Liang, L.; Li, X.; Tan, X.; Wei, L.; Lei, B.; Kijlstra, A.; Yang, P. microRNA-146a and Ets-1 gene polymorphisms in ocular Behcet’s disease and Vogt-Koyanagi-Harada syndrome. Ann. Rheum. Dis. 2014, 73, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Keerthana, R.; Pazhanimuthu, A.; Perumal, P. Overexpression of circulating miRNA-21 and miRNA-146a in plasma samples of breast cancer patients. Indian J. Biochem. Biophys. 2013, 50, 210–214. [Google Scholar] [PubMed]
- Wang, X.; Tang, S.; Le, S.Y.; Lu, R.; Rader, J.S.; Meyers, C.; Zheng, Z.M. Aberrant expression of oncogenic and tumor-suppressive micrornas in cervical cancer is required for cancer cell growth. PLoS ONE 2008, 3, e2557. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Neilson, J.R.; Kumar, P.; Manocha, M.; Shankar, P.; Sharp, P.A.; Manjunath, N. miRNA profiling of naive, effector and memory CD8 T cells. PLoS ONE 2007, 2, e1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.; Kim, H.A.; Suh, C.H.; Byun, H.O.; Jung, J.Y.; Sohn, S. The relevance of miRNA-21 in HSV-induced inflammation in a mouse model. Int. J. Mol. Sci. 2015, 16, 7413–7427. [Google Scholar] [CrossRef] [PubMed]
- Sugi-Ikai, N.; Nakazawa, M.; Nakamura, S.; Ohno, S.; Minami, M. Increased frequencies of interleukin-2- and interferon-gamma-producing T cells in patients with active Behcet’s disease. Invest. Ophthalmol. Vis. Sci. 1998, 39, 996–1004. [Google Scholar] [PubMed]
- Alpsoy, E.; Cayirli, C.; Er, H.; Yilmaz, E. The levels of plasma interleukin-2 and soluble interleukin-2R in Behcet’s disease: A marker of disease activity. J. Dermatol. 1998, 25, 513–516. [Google Scholar] [PubMed]
- Choi, J.Y.; Choi, B.; Shim, J.A.; Lee, E.S.; Kim, D.Y.; Bang, D.; Sohn, S. IL-2/IL-2 antibody immune complex regulates HSV-induced inflammation through induction of IL-2 receptor alpha, beta, and gamma in a mouse model. Immunobiology 2015, 220, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Castro, I.; Yu, A.; Dee, M.J.; Malek, T.R. The basis of distinctive IL-2- and IL-15-dependent signaling: Weak CD122-dependent signaling favors CD8+ T central-memory cell survival but not t effector-memory cell development. J. Immunol. 2011, 187, 5170–5182. [Google Scholar] [CrossRef] [PubMed]
- Choe, J.Y.; Lee, H.; Kim, S.G.; Kim, M.J.; Park, S.H.; Kim, S.K. The distinct expressions of interleukin-15 and interleukin-15 receptor alpha in Behcet’s disease. Rheumatol. Int. 2013, 33, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Luan, L.; Rabacal, W.; Bohannon, J.K.; Fensterheim, B.A.; Hernandez, A.; Sherwood, E.R. IL-15 superagonist-mediated immunotoxicity: Role of NK cells and IFN-γ. J. Immunol. 2015, 195, 2353–2364. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.M.S.; Choi, B.; Choi, J.; Lee, E.-S.; Sohn, S. Frequencies of IL-15rα+ cells in patients with Behçet’s disease and the effects of overexpressing IL-15rα+ on disease symptoms in mice. Cytokine 2018, 110, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Shah, P.K.; Faure, E.; Equils, O.; Thomas, L.; Fishbein, M.C.; Luthringer, D.; Xu, X.P.; Rajavashisth, T.B.; Yano, J.; et al. Toll-like receptor-4 is expressed by macrophages in murine and human lipid-rich atherosclerotic plaques and upregulated by oxidized LDL. Circulation 2001, 104, 3103–3108. [Google Scholar] [CrossRef] [PubMed]
- Nara, K.; Kurokawa, M.S.; Chiba, S.; Yoshikawa, H.; Tsukikawa, S.; Matsuda, T.; Suzuki, N. Involvement of innate immunity in the pathogenesis of intestinal Behcet’s disease. Clin. Exp. Immunol. 2008, 152, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Do, J.E.; Kwon, S.Y.; Park, S.; Lee, E.S. Effects of vitamin d on expression of Toll-like receptors of monocytes from patients with Behcet’s disease. Rheumatology 2008, 47, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, L.B.; Nashold, F.E.; Spach, K.M.; Hayes, C.E. 1,25-dihydroxyvitamin D3 reverses experimental autoimmune encephalomyelitis by inhibiting chemokine synthesis and monocyte trafficking. J. Neurosci. Res. 2007, 85, 2480–2490. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, K.; Wessner, B.; Laggner, U.; Ploder, M.; Tamandl, D.; Friedl, J.; Zugel, U.; Steinmeyer, A.; Pollak, A.; Roth, E.; et al. Vitamin D3 down-regulates monocyte TLR expression and triggers hyporesponsiveness to pathogen-associated molecular patterns. Eur. J. Immunol. 2006, 36, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.; Lee, E.S.; Sohn, S. Vitamin D3 ameliorates herpes simplex virus-induced Behcet’s disease-like inflammation in a mouse model through down-regulation of Toll-like receptors. Clin. Exp. Rheumatol. 2011, 29, S13–S19. [Google Scholar] [PubMed]
- Choi, B.; Suh, C.H.; Kim, H.A.; Sayeed, H.M.; Sohn, S. The correlation of CD206, CD209, and disease severity in Behcet’s disease with arthritis. Med. Inflamm. 2017, 2017, 7539529. [Google Scholar] [CrossRef] [PubMed]
- Shannon, B.; Yi, T.J.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; Tharao, W.; Huibner, S.; Remis, R.; Rebbapragada, A.; et al. Impact of asymptomatic herpes simplex virus type 2 infection on mucosal homing and immune cell subsets in the blood and female genital tract. J. Immunol. 2014, 192, 5074–5082. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Sayeed, H.M.; Islam, S.M.S.; Sohn, S. Role of N-acetyl galactosamine-4-SO4, a ligand of CD206 in HSV-induced mouse model of Behcet’s disease. Eur. J. Pharmacol. 2017, 813, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Guasp, P.; Barnea, E.; Gonzalez-Escribano, M.F.; Jimenez-Reinoso, A.; Regueiro, J.R.; Admon, A.; Lopez de Castro, J.A. The Behcet’s disease-associated variant of the aminopeptidase ERAP1 shapes a low-affinity HLA-B*51 peptidome by differential subpeptidome processing. J. Biol. Chem. 2017, 292, 9680–9689. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models and Symptoms | Published Number of Papers |
|---|---|
| Environmental pollutants→Mucocutaneous lesions in miniature swine | 1 |
| Human 60 kD heat shock protein-derived peptide 336–351→Uveitis in rats | 1 |
| HLA-B51 Transgenic mice→Excessive function of peripheral blood neutrophils but no symptoms | 1 |
| Herpes simplex virus type 1→Oral, genital, and skin ulcers, eye involvement, arthritis, and intestinal involvement in ICR mice and C57BL/6 mice | 26 |
| Tropomyosin→Inflammation in the skin, joints, and eyes of Lewis rats | 2 |
| Retinal soluble antigen (S-Ag)→Uveitis in rats | 1 |
| Sera from seven NeuroBD patients→Reduced locomotor activity in rats | 1 |
| Categories | Applied Materials | Therapy Targets |
|---|---|---|
| Conventional therapies | Thalidomide Infliximab Etanercept | ↓TNFα ↓TNFα ↓TNFα |
| Natural products | Derivatives of Gentiana macrophylla Radix Taraxacum mongolicum Hand.-Mazz. Chitosan | ↓TNFα ↑IL-10 ↑IL-4 |
| Potential therapies | IL-4 C-C chemokine ligand 21 Poly I:C TNFα siRNA IL-6 siRNA miRNA-21 antagomir IL-2/IL-2 Ab complex, IL-15/IL-15R-Fc complex Vitamin D3 4-sulfated N-acetyl galactosamine | ↑IL-4 ↑CCR7 ↑IL-15Rα, IL-10 ↓TNFα ↓IL-6 ↓miRNA-21 ↑IL-2Rα, ↑IL-15Rα ↓TLR-2, ↓TLR-4, ↓CD206 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, S.M.S.; Sohn, S. HSV-Induced Systemic Inflammation as an Animal Model for Behçet’s Disease and Therapeutic Applications. Viruses 2018, 10, 511. https://doi.org/10.3390/v10090511
Islam SMS, Sohn S. HSV-Induced Systemic Inflammation as an Animal Model for Behçet’s Disease and Therapeutic Applications. Viruses. 2018; 10(9):511. https://doi.org/10.3390/v10090511
Chicago/Turabian StyleIslam, S. M. Shamsul, and Seonghyang Sohn. 2018. "HSV-Induced Systemic Inflammation as an Animal Model for Behçet’s Disease and Therapeutic Applications" Viruses 10, no. 9: 511. https://doi.org/10.3390/v10090511
APA StyleIslam, S. M. S., & Sohn, S. (2018). HSV-Induced Systemic Inflammation as an Animal Model for Behçet’s Disease and Therapeutic Applications. Viruses, 10(9), 511. https://doi.org/10.3390/v10090511