Antibody Responses to Bovine Alphaherpesvirus 1 (BoHV-1) in Passively Immunized Calves


Abstract
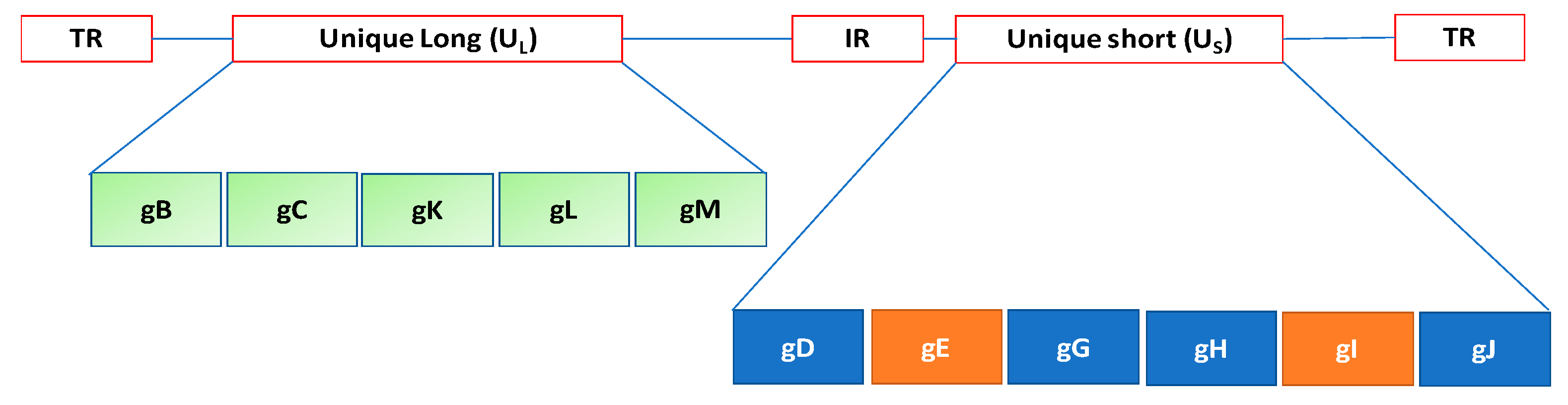
:1. Introduction
2. Passive Immunity from Non-Marker Vaccines
2.1. Colostrum
2.2. Protection Provided by Colostrum
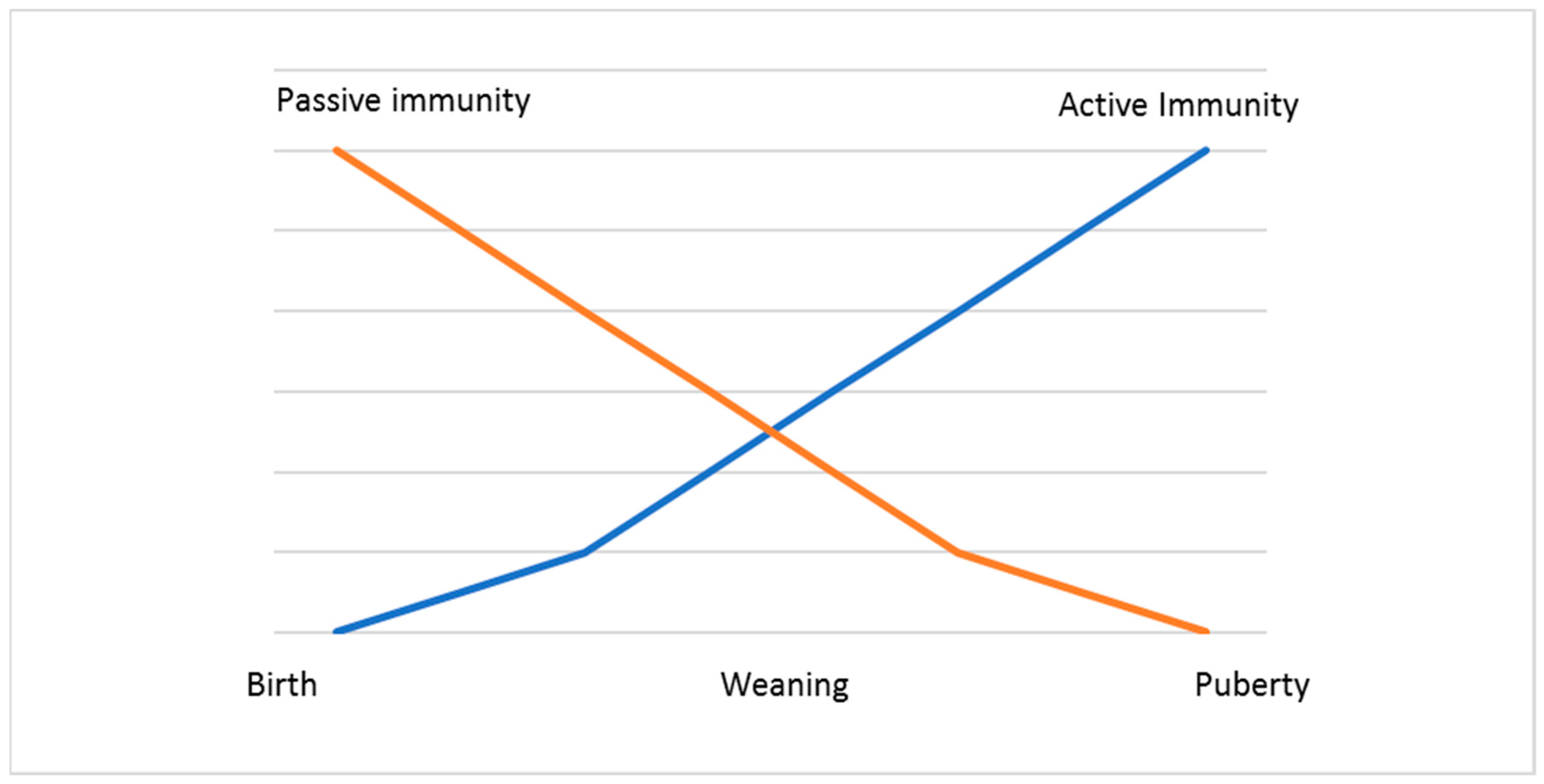
2.3. Passive Immunity
2.4. Vaccines
2.5. Seronegative Latent Carriers (SNLCs)
3. Passive Immunity from Marker Vaccines
3.1. Antibody Response
3.2. Clinical Form
3.3. Latency
3.4. Excretion
3.5. Disadvantages
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Fauquet, C.M.; Mayo, M.A.; Maniloff, J.; Desselberger, U.; Ball, L.A. Family Herpesviridae. In Virus Taxonomy; Eight Report of the International Committee on Taxonomy of Viruses; Elsevier Academic Press: San Diego, CA, USA, 2006; pp. 193–212. ISBN 13 978-0-12-249951-7. [Google Scholar]
- Jones, C.; Chowdhury, S. A review of the biology of bovine herpesvirus type 1 (BoHV-1), its role as a cofactor in the bovine respiratory disease complex and development of improved vaccines. Anim. Health Res. Rev. 2008, 8, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Levings, R.L.; Roth, J.A. Immunity to bovine herpesvirus 1: I. Viral lifecycle and innate immunity. Anim. Health Res. Rev. 2013, 14, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Levings, R.L.; Roth, J.A. Immunity to bovine herpesvirus 1: II. Adaptative immunity and vaccinology. Anim. Health Res. Rev. 2013, 14, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Rixon, F.J. Structure and assembly of herpesviruses. Semin. Virol. 1993, 4, 135–144. [Google Scholar] [CrossRef]
- Murphy, F.A.; Gibbs, E.P.J.; Horzinek, M.C.; Studdert, M.J. Herpesviridae. In Veterinary Virology, 9th ed.; Academic Press: London, UK, 2006; pp. 301–325. ISBN 0-12-511340-4. [Google Scholar]
- Fulton, R.W.; d’Offay, J.M.; Eberle, R.; Moeller, R.B.; Van Campen, H.; O’Toole, D.; Chase, C.; Miller, M.M.; Sprowls, R.; Nydam, D.V. Bovine herpesvirus-1: Evaluation of genetic diversity of subtypes derived from fields strains of varied clinical syndromes and their relationship to vaccine strains. Vaccine 2015, 33, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Raaperi, K.; Orro, T.; Viltrop, A. Epidemiology and control of bovine herpesvirus 1 infection in Europe. Vet. J. 2014, 201, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Maresca, C.; Scoccia, E.; Dettori, A.; Felici, A.; Guarcini, R.; Petrini, S.; Quaglia, A.; Filippini, G. National surveillance plan for infectious bovine rhinotracheitis (IBR) in autochthonous Italian cattle breeds: Results of first year of activity. Vet. Microbiol. 2018, 219, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Whetstone, C.A.; van der Maaten, M.J. Abortifacient property of bovine herpesvirus type 1 isolates that represent three subtypes determined by restriction endonuclease analysis of viral DNA. Am. J. Vet. Res. 1991, 52, 458–461. [Google Scholar] [PubMed]
- Edwards, S.; White, H.; Nixon, P. A study of the predominant genotypes of bovid herpesvirus 1 found in the U.K. Vet. Microbiol. 1990, 22, 213–223. [Google Scholar] [CrossRef]
- Wentink, G.H.; van Oirschot, J.T.; Verhoeff, J. Risk of infection with bovine herpesvirus 1 (BHV1): A review. Vet. Q. 1993, 15, 30–33. [Google Scholar] [CrossRef]
- Ludwing, H. Bovine herpesviruses. In The Herpesviruses; Plenum Press: New York, NY, USA, 1983; Volume 2, pp. 133–214. [Google Scholar]
- Beer, M.; Dastjerdy, A. Chapter 2.4.12 Infectious bovine rhinotracheitis/infectious pustular vulvovaginitis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 7th ed.; Vallant, B., Edwards, S., Eds.; Office International Des Epizooties: Paris, France, 2017; Volume 1, pp. 1–19. ISBN 978-92-9044-878-5. [Google Scholar]
- Mechor, G.D.; Rousseaux, G.C.; Radostits, O.M.; Babiuk, L.A.; Petrie, L. Protection of Newborn Calves against Fatal Multisystemic Infectious Bovine Rhinotracheitis by Feeding Colostrum from Vaccinated Cows. Can. J. Vet. Res. 1987, 51, 452–459. [Google Scholar]
- Ackerman, M.; Wyler, R. The DNA of an IPV strain of bovid herpesvirus 1 in sacral ganglia during latency after intravaginal infection. Vet. Microbiol. 1984, 9, 53–63. [Google Scholar] [CrossRef]
- Winkler, M.T.; Doster, A.; Jones, C. Persistence and reactivation of bovine herpesvirus 1 in the tonsils of latently infected calves. J. Virol. 2000, 74, 5337–5346. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Peterhans, E.; Wyler, R. DNA of bovine herpesvirus type 1 in the trigeminal ganglia of latently infected calves. Am. J. Vet. Res. 1982, 43, 36–40. [Google Scholar] [PubMed]
- Van Drunen Littel-van den Hurk, S.; Tikoo, S.K.; van den Hurk, J.V.; Babiuk, L.A.; Van Donkersgoed, J. Protective immunity in cattle following vaccination with conventional and marker bovine herpesvirus-1 (BHV1) vaccines. Vaccine 1997, 15, 36–44. [Google Scholar] [CrossRef]
- Belknap, E.B.; Walters, L.M.; Kelling, C.; Ayers, V.K.; Norris, J.; McMillen, J.; Hayhow, C.; Cochran, M.; Reddy, D.N.; Wright, J.; et al. Immunogenicity and protective efficacy of a gE, gG, and US2 gene-deleted bovine herpesvirus-1 (BHV-1) vaccine. Vaccine 1999, 17, 2297–2305. [Google Scholar] [CrossRef]
- Castrucci, G.; Frigeri, F.; Salvatori, D.; Ferrari, M.; Sardonini, Q.; Cassai, E.; Lo, D.M.; Rotola, A.; Angelini, R. Vaccination of calves against bovine herpesvirus-1: Assessment of the protective value of eight vaccines. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 29–41. [Google Scholar] [CrossRef]
- Ackermann, M.; Engels, M. Pro and contra IBR-eradication. Vet. Microbiol. 2006, 113, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, M.; Klapper, P.E. Herpes Simplex Virus Type 1 and type 2. In Principle & Practice of Clinical Virology, 6th ed.; Zuckerman, A.J., Banatvala, J.E., Scoub, B.D., Griffiths, P.D., Mortimer, P., Eds.; Wiley-Blackwell: West Sessex, UK, 2009; pp. 96–131. ISBN 978-0-470-51799-4. [Google Scholar]
- European Commission. Report on Bovine Herpesvirus 1 (BHV-1) Marker Vaccines and the Accompanying Diagnostic Tests; 25/10/2000 (SANCO/C3/AH/R20/2000); Scientific Committee on Animal Health and Animal Welfare: Brussels, Belgium, 2000. [Google Scholar]
- European Medicine Agency. Dossier of Scientific Discussion, EMA/CVMP/743800/2010 Veterinary Medicines and Product Data Management; European Medicine Agency: London, UK, 2010. [Google Scholar]
- Stube, W.; Abar, B.; Bergle, R.D.; Block, W.; Heinen, E.; Kretzdorn, D.; Rodenbach, C.; Schmeer, N. Safety Aspects in the Development of an Infectious Bovine Rhinotracheitis Marker Vaccine. Dev. Biol. Stand. 1995, 84, 75–81. [Google Scholar]
- Nuotio, L.; Neuvonen, E.; Hyytiainen, M. Epidemiology and eradication of infectious bovine rhinotracheitis/infectious pustular vulvovaginitis (IBR/IPV) virus in Finland. Acta Vet. Scand. 2007, 49, 1–6. [Google Scholar] [CrossRef]
- Turin, L.; Russo, S. BHV-1 infection in cattle: an update. Vet. Bull. 2003, 73, 15–21. [Google Scholar]
- Zhang, M.; Shulin, F.; Deng, M.; Xie, Q.; Xu, H.; Liu, Z.; Hu, C.; Chen, H.; Guo, A. Attenuation of bovine herpesvirus type 1 by deletion of its glycoprotein G and tk genes and protection against virulent viral challenge. Vaccine 2011, 29, 8943–8950. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Bandyopadhyay, S.; Dimri, U.; Patra, P.H. Bovine herpesvirus-1 a re-emerging concern in livestock: A revisit to its biology, epidemiology, diagnosis, and prophylaxis. Vet. Q. 2013, 33, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Castrucci, G.; Ferrari, M.; Marchini, C.; Salvatori, D.; Provinciali, M.; Tosini, A.; Petrini, S.; Sardonini, Q.; Lo, D.M.; Frigeri, F.; et al. Immunization against bovine herpesvirus-1 infection. Preliminary tests in calves with a DNA vaccine. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Petrini, S.; Ramadori, G.; Corradi, A.; Borghetti, P.; Lombardi, G.; Villa, R.; Bottareli, E.; Guercio, A.; Amici, A.; Ferrari, M. Evaluation of safety and efficacy of DNA vaccines against bovine herpesvirus-1 (BoHV-1) in calves. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; Boni, M.; Di Luca, D.; Salvatori, D.; Vita, A.; Cassai, E. A combined bovine herpesvirus 1 gB-gD DNA vaccines induces immune response in mice. Comp. Immunol. Microbiol. Infect. Dis. 2005, 28, 155–166. [Google Scholar] [CrossRef]
- Kendrik, J.W.; York, C.J.; Mckercher, D.G. A controlled field trial of a vaccine for infectious bovine rhinotracheitis. Proc. U. S. Livest. Sanit. Assoc. 1956, 60, 155–158. [Google Scholar]
- Schwarz, A.J.F.; York, C.J.; Zirbel, L.W.; Estela, L.A. Modification of infectious bovine rhinotracheitis (IBR) virus in tissue culture and development of a vaccine. Proc. Soc. Exp. Biol. Med. 1957, 96, 453–458. [Google Scholar] [CrossRef]
- Phillips, C.E. Potency evaluation of vaccines used against bovine respiratory diseases. J. Am. Vet. Med. Assoc. 1968, 152, 842–845. [Google Scholar]
- Phillip, J.I.H.; Darbyshire, J.H. Respiratory viruses of cattle. Adv. Vet. Sci. 1971, 15, 159–199. [Google Scholar]
- Gibbs, E.P.J.; Rweyemamu, M.M. Bovine herpesviruses. Part I. Bovine herpesvirus 1. Vet. Bull. 1977, 47, 317–343. [Google Scholar]
- Tizard, I.R. Immunity at body surface (Chapter 20). In Veterinary Immunology: An Introduction, 6th ed.; Saunders: London, UK, 2000; pp. 222–234. [Google Scholar]
- Matte, J.J.; Girard, C.L.; Seoane, J.R.; Brisson, G.J. Adsorption of colostral immunoglobulin G in the newborn dairy calf. J. Dairy Sci. 1982, 65, 1765–1770. [Google Scholar] [CrossRef]
- Chase, C.L.; Hurley, D.J.; Reber, A.J. Neonatal immune development in the Calf and Its Impact on Vaccine response. Vet. Clin. Food Anim. 2008, 24, 87–104. [Google Scholar] [CrossRef]
- Robinson, J.D.; Stott, G.H.; DeNise, S.K. Effects of passive immunity on growth and survival in the dairy heifer. J. Dairy Sci. 1988, 71, 1283–1287. [Google Scholar] [CrossRef]
- Wittum, T.E.; Perino, L.J. Passive immune status at postpartum hour 24 and long-term health and performance of calves. Am. J. Vet. Res. 1995, 56, 1149–1154. [Google Scholar] [PubMed]
- Jones, C.; Heinrichs, J. Colostrum Supplements and Replacer Penn State. Available online: https://extension.psu.edu/colostrum-supplements-and-replacer (accessed on 5 May 2016).
- Chammorro, M.F.; Walz, P.H.; Hains, D.M.; Passler, T.; Earleywine, T.; Palomares, R.A.; Riddel, K.P.; Galik, P.; Zhang, Y.; Givens, M.D. Comparision of levels and duration of detection of antibodies to bovine viral diarrhea virus 1, bovine viral diarrhea virus 2, bovine respiratory syncytial virus, bovine herpesvirus 1, and bovine parainfluenza virus 3 in calves fed maternal colostrum or colostrum-replacement product. Can. J. Vet. Res. 2014, 78, 81–88. [Google Scholar]
- Weaver, D.M.; Tyler, J.W.; VanMetre, D.C.; Hostetler, D.E.; Barrington, G.M. Passive transfer of colostral immunoglobulins in calves. J. Vet. Intern. Med. 2000, 14, 569–577. [Google Scholar] [CrossRef]
- Quigley, J.D.; Kost, C.J.; Wolfe, T.M. Adsorption of protein and IgG in calves fed a colostrum supplement or replacer. J. Dairy Sci. 2002, 85, 1243–1248. [Google Scholar] [CrossRef]
- Chelack, B.J.; Morley, P.S.; Haines, D.M. Evaluation of methods for dehydration of bovine colostrum for total replacement of normal colostrum in calves. Can. Vet. J. 1993, 34, 407–412. [Google Scholar]
- Husband, A.J.; Lascelles, A.K. Antibody responses to neonatal immunization in calves. Res. Vet. Sci. 1975, 18, 201–207. [Google Scholar] [CrossRef]
- Pospisil, Z.; Krejci, J.; Rodax, C. Demonstration of colostral antibodies in the nasal secretion of calves and their protective effect against infection. Acta Vet. Brno 1983, 52, 59–65. [Google Scholar] [CrossRef]
- Rouse, B.T.; Babiuk, L.A. Mechanisms of recovery from herpesvirus infections—A review. Can. J. Comp. Med. 1978, 42, 414–427. [Google Scholar] [PubMed]
- Patel, J.R. Characteristics of live bovine herpesvirus-1 vaccines. Vet. J. 2005, 169, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Straub, O.C. Zur ubertragung vivaler antikorper durch das kolostrum. Tierarztl. Umsch. 1969, 24, 571–573. [Google Scholar]
- Menanteau-Horta, A.M.; Ames, T.R.; Johnson, D.W. Effect of maternally antibody upon vaccination with infectious bovine rhinotracheitis and bovine virus diarrhea vaccines. Can. J. Comp. Med. 1985, 49, 10–14. [Google Scholar]
- Kirkpatrick, J.; Fulton, R.W.; Burge, L.J.; Dubois, W.R.; Payton, M. Passively transferred immunity in newborn calves, rate of antibody decay, and effect on subsequent vaccination with modified live virus vaccine. Bov. Pract. 2001, 35, 47–55. [Google Scholar]
- Munoz-Zanzi, C.A.; Thurmond, M.C.; Johnson, W.O.; Heitela, S.K. Predicted ages of dairy calves when colostrum-derived bovine viral diarrhea virus antibodies would no longer offer protection against diseases or interfere with vaccination. J. Am. Vet. Med. Assoc. 2002, 221, 678–685. [Google Scholar] [CrossRef]
- Smith, B.I.; Rieger, R.H.; Dickens, C.M.; Schultz, R.D.; Aceto, H. Anti-bovine herpesvirus and anti-bovine viral diarrhea virus antibody responses in pregnant Holstein dairy cattle following administration of a multivalent killed virus vaccine. Am. J. Vet. Res. 2015, 76, 913–920. [Google Scholar] [CrossRef]
- Archambault, D.; Morin, G.; Elazhary, Y.; Roy, R.S.; Joncas, J.H. Immune response of pregnant heifers and cows to bovine rotavirus inoculation and passive protection to rotavirus infection in newborn calves fed colostral antibodies or colostral lymphocytes. Am. J. Vet. Res. 1988, 49, 1084–1091. [Google Scholar]
- Mallard, B.A.; Wagtner, L.C.; Ireland, M.J.; Dekkers, J.C.M. Effect of growth hormone, insulin-like growth factor-I, and cortisol and periparturient antibody response profiles of dairy cattle. Vet. Immunol. Immunopathol. 1997, 60, 61–76. [Google Scholar] [CrossRef]
- Kimman, T.G.; Westenbrink, F.; Straver, P.J. Priming for local and systemic antibody memory response to bovine respiratory syncytial virus: Effect of amount of virus, viral replication, route of administration and maternal antibodies. Vet. Immunol. Immunophatol. 1989, 22, 145–160. [Google Scholar] [CrossRef]
- Kirkpatrick, J.G.; Step, D.L.; Payton, M.E.; Richards, J.B.; McTague, L.F.; Saliki, J.T.; Confer, A.W.; Cook, B.J.; Ingram, S.H.; Wringht, J.C. Effect of age at the time of vaccination on antibody titers and feedlot performance in beef calves. J. Am. Vet. Med. Assoc. 2008, 233, 136–142. [Google Scholar] [CrossRef]
- Van der Poel, W.H.M.; Midel, W.G.J.; Schukken, Y.H. Antibody titers against bovine respiratory syncytial virus in colostrum-fed dairy calves born in various seasons. Am. J. Vet. Res. 1999, 60, 1098–1101. [Google Scholar] [PubMed]
- Fulton, R.W.; Briggs, R.E.; Payton, M.E.; Confer, W.A.; Saliki, J.T.; Ridpath, J.F.; Burge, L.J.; Duff, G.C. Maternally derived humoral immunity to bovine viral diarrhea virus (BVDV) 1a, BVDV 1b, BVDV2, Bovine herpesvirus-1, parainfluenza-3 virus, bovine respiratory syncytial virus, Mannheimia haemolitica and Pasteurella Multocida in beef calves, antibody decline by half-life studies and effect on response to vaccination. Vaccine 2004, 22, 643–649. [Google Scholar] [CrossRef]
- Brar, J.S.; Johnson, D.W.; Muscoplat, C.C.; Shope, R.E.; Meiske, J.C. Maternal immunity to infectious bovine rhinotracheitis and bovine viral diarrhea viruses. Am. J. Vet. Res. 1978, 60, 1098–1101. [Google Scholar]
- Mechor, G.D.; Virtala, A.M.K.; Dubovi, E.J.; Grohn, Y.T. The half-life for maternally derived immunoglobulin G anti-viral antibodies in data from an observational field study. Bov. Pract. 2001, 35, 131–136. [Google Scholar]
- Ellis, J.A.; Hassard, L.E.; Cortese, V.S.; Morley, P.S. Effects of perinatal vaccination on humoral and cellular immune responses in cows and young calves. J. Am. Vet. Med. Assoc. 1996, 208, 393–400. [Google Scholar]
- Brodshaw, B.J.; Edwards, S. Antibody isotype responses to experimental infection with bovine herpesvirus 1 in calves with colostrally derived antibody. Vet. Microbiol. 1996, 53, 143–151. [Google Scholar] [CrossRef]
- Edwards, S.; Newman, R.H.; White, H. The virulence of British isolates of Bovid herpesvirus 1 in relationship to viral genotype. Br. Vet. J. 1991, 147, 216–231. [Google Scholar] [CrossRef]
- Madic, J.; Magdalena, J.; Quack, J.; Van Oirschot, J.T. Isotype-specific antibody responses in sera and mucosal secretions of calves experimentally infected with bovine herpesvirus 1. Vet. Immunol. Immunophatol. 1995, 46, 267–283. [Google Scholar] [CrossRef]
- Lemaire, M.; Meyer, G.; Ernst, E.; Vanherreweghe, V.; Limbourg, B.; Pastoret, P.P.; Thiry, E. Latent bovine herpesvirus 1 infection in calves protected by colostral immunity. Vet. Rec. 1995, 137, 70–71. [Google Scholar] [CrossRef] [PubMed]
- Endsley, J.J.; Roth, J.A.; Ridpath, J.; Neill, J. Maternal antibody blocks humoral but not T cell responses to BVDV. Biologicals 2003, 31, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Todd, J.D. Intranasal vaccination of cattle against IBR and PI3: Field and laboratory observations in dairy, beef and neonatal calf populations. Dev. Biol. Stand. 1976, 33, 391–395. [Google Scholar] [PubMed]
- Aboul Soud, S.M.; El Sayed, M.; Zedan, S.M.; Wasel, M.S.; El-Sawalhy, A.A. Evaluation of a live trivalent vaccine for infectious bovine rhinotracheitis virus, para-influenza type-3 and Pasteurella multocida in calves. Egypt. J. Immunol. 2004, 11, 101–108. [Google Scholar]
- Fitzpatrick, D.R.; Snider, m.; McDougall, L.; Beskorwayne, T.; Babiuk, L.A.; Zamb, T.J.; Bielefeldt Ohmann, H. Molecular mimicry: A herpes virus glycoprotein antigenically related to a cell-surface glycoprotein expressed by macrophages, polymorphonuclear leucocytes, and platelets. Immunology 1990, 70, 504–512. [Google Scholar] [PubMed]
- Lemaire, M.; Weynants, V.; Godfroid, J.; Schynts, F.; Meyer, G.; Letesson, J.J.; Thiry, E. Effects of bovine herpesvirus type 1 infection in calves with maternal antibodies on immune response and virus latency. J. Clin. Microbiol. 2000, 38, 1885–1894. [Google Scholar] [PubMed]
- Lemaire, M.; Schynts, F.; Meyer, G.; Georgin, J.P.; Baranowski, E.; Gabriel, A.; Ros, C.; Belak, S.; Thiry, E. Latency and reactivation of glycoprotein E negative bovine herpesvirus type 1 vaccine: Influence of virus load and effect of specific maternal antibodies. Vaccine 2001, 19, 4795–4804. [Google Scholar] [CrossRef]
- Turin, L.; Russo, S.; Poli, G. BHV-1: New molecular approaches to control a common and widespread infection. Mol. Med. 1999, 5, 261–284. [Google Scholar] [CrossRef]
- Castrucci, G.; Frigeri, F.; Salvatori, D.; Ferrari, M.; Lo Dico, M.; Rotola, A.; Sardonini, Q.; Petrini, S.; Cassai, E. A study on latency in calves by five vaccines against bovine herpesvirus-1 infection. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 205–215. [Google Scholar] [CrossRef]
- Lemaire, M.; Schynts, F.; Meyer, G.; Thiry, E. Antibody response to glycoprotein E after bovine herpesvirus type 1 infection in passively immunized, glycoprotein E-negative calves. Vet. Rec. 1999, 144, 172–176. [Google Scholar] [CrossRef]
- Lemaire, M.; Mayer, G.; Baranowski, E.; Schynts, F.; Wellemans, G.; Kerkhofs, P.; Thiry, E. Production of Bovin Herpesvirus Type 1-Seronegative latent Carriers by Administration of a Live-Attenuated Vaccine in Passively Immunized Calves. J. Clin. Microbiol. 2000, 38, 4233–4238. [Google Scholar] [PubMed]
- Hage, J.J.; Glas, R.D.; Westra, H.H.; Maris-Veldhuis, M.A.; van Oirschot, J.T.; Rijsewijk, F.A. Reactivation of latent bovine herpesvirus 1 in cattle seronegative to glycoproteins gB and gE. Vet. Microbiol. 1998, 60, 87–98. [Google Scholar] [CrossRef]
- Kucera, C.J.; White, R.G.; Beckenhauer, W.H. Evaluation of the safety and efficacy of an intranasal vaccine containing a temperature-sensitive strain of infectious bovine rhinotracheitis virus. Am. J. Vet. Res. 1978, 39, 607–610. [Google Scholar]
- Zygraich, N.; Lobmann, M.; Vascoboinic, E.; Berge, E.; Huygelen, C. In vivo and in vitro properties of a temperature sensitive mutant of infectious bovine rhinotracheitis virus. Res. Vet. Sci. 1974, 16, 328–335. [Google Scholar] [CrossRef]
- Nettleton, P.F.; Sharp, J.M. Infectious bovine rhinotracheitis virus excretion after vaccination. Vet. Rec. 1980, 107, 379. [Google Scholar] [CrossRef]
- Frerichs, G.N.; Woods, S.B.; Lucas, M.H.; Sands, J.J. Safety and efficacy of live and inactivated infectious bovine rhinotracheitis vaccines. Vet. Rec. 1982, 111, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, M.; Hanon, E.; Schynts, F.; Meyer, G.; Thiry, E. Specific passive immunity reduces the excretion of glycoprotein E-negative bovine herpesvirus type 1 vaccine strain in calves. Vaccine 2001, 19, 1013–1017. [Google Scholar] [CrossRef]
- Pastoret, P.P.; Thiry, E. Diagnosis and prophylaxis of infectious bovine rhinotracheitis: Role of virus latency. Comp. Immunol. Microbiol. Infect. Dis. 1985, 8, 35–42. [Google Scholar] [CrossRef]
- North American Compendiums. Compendium of Veterinary Products, 12th ed.; North American Compendiums: Port Huron, MI, USA, 2010; pp. 1–1848. [Google Scholar]
- Van Drunen Littel-van den Hurk, S. Cell-mediated immune responses induced by BHV-1: Rational vaccine design. Expert Rev. Vaccines 2007, 6, 369–380. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; Kaashoek, M.J.; Moerman, A.; Madic, J.; Rijsewijk, F.A.; Quak, J.; Gielkens, A.L.; van Oirschot, J.T. A Conventionally attenuated glycoprotein E-negative strain of bovine herpesvirus type 1 is an efficacious and safe vaccine. Vaccine 1994, 12, 239–444. [Google Scholar] [CrossRef]
- Van Oirshot, J.T.; Kaashoek, M.J.; Maris-Veldhuis, M.A.; Weerd-meester, K.; Rijsewijk, F.A. An enzyme-linked immunosorbent assay to detect antibodies against glycoprotein gE of bovine herpesvirus 1 allows differentiation between infected and vaccinated cattle. J. Virol. Methods 1997, 67, 23–34. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; van Oirschot, J.T. Early immunity induced by a live gE-negative bovine herpesvirus 1 marker vaccine. Vet. Microbiol. 1996, 53, 191–197. [Google Scholar] [CrossRef]
- Tignon, M.; De Baere, M.; Hanon, J.B.; Goolaerts, A.; Houtain, J.Y.; Delooz, L.; Cay, A.B. Characterization of three commercial ELISA kits for detection of BOHV-1 gE specific antibodies in serum and milk samples and applicability of bulk milk for determination of herd status. J. Virol. Methods 2017, 245, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Muratore, E.; Bertolotti, L.; Nogarol, C.; Caruso, C.; Lucchese, L.; Lotti, B.; Ariello, D.; Moresco, A.; Masoero, L.; Nardelli, S.; et al. Surveillance of Infectious Bovine Rhinotracheitis in marker-vaccinated dairy herds: Application of a recombinant gE ELISA on bulk milk samples. Vet. Immunol. Immunopathol. 2017, 185, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, L.; Muratore, E.; Nogarol, C.; Caruso, C.; Lucchese, L.; Profiti, M.; Anfossi, L.; Masoero, L.; Nardelli, S.; Rosati, S. Development and validation of an indirect ELISA as a confirmatory test for surveillance of infectious bovine rhinotracheitis in vaccinated herds. BMC Vet. Res. 2015, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Van Oirschot, J.T.; Kaashoek, M.J.; Rijsewijk, F.A.; Stegeman, J.A. The use of marker vaccines in eradication of herpesviruses. J. Biotechnol. 1996, 44, 75–81. [Google Scholar] [CrossRef]
- Strube, W.; Auer, S.; Block, W.; Heinen, E.; Kretzdorn, D.; Rodenbach, C.; Schmeer, N. A gE deleted infectious bovine rhinotracheitis marker vaccine for use in improved bovine herpesvirus 1 control programs. Vet. Microbiol. 1996, 53, 181–189. [Google Scholar] [CrossRef]
- Petrini, S.; Ferrari, M.; Vincenzetti, S.; Vita, A.; Amici, A.; Ramadori, G. Immunoenzyme linked assay (ELISA) for the detection of antibodies to truncated glycoprotein D (tgD) of Bovine herpesvirus-1. Vet. Res. Commun. 2006, 30, 257–259. [Google Scholar] [CrossRef]
- Jacobs, L. Glycoprotein E of pseudorabies virus and homologous proteins in other alphaherpesvirinae. Arch. Virol. 1994, 137, 209–228. [Google Scholar] [CrossRef]
- Kit, S.; Qavi, H.; Gaines, J.D.; Billingsley, P.; McConnell, S. Thymidine kinase-negative bovine herpesvirus type 1 mutant is stable and highly attenuated in calves. Arch. Virol. 1985, 86, 63–83. [Google Scholar] [CrossRef]
- Kaaschoek, M.J.; Straver, P.J.; Van Rooij, E.M.A.; Quak, J.; van Oirschot, J.T. Virulence, immunogenicy and reactivation of seven bovine herpesvirus 1.1 strains: Clinical and virological aspects. Vet. Rec. 1996, 139, 416–421. [Google Scholar] [CrossRef]
- European Medicine Agency. Scientific Discussion, EMA/CVMP/743800/2010; European Medicine Agency: London, UK, 2010; pp. 1–48. [Google Scholar]
- Schynts, F.; Lemaire, M.; Ros, C.; Belak, S.; Thiry, E. Establishment of latency associated with glycoprotein E (gE) seroconversion after bovine herpesvirus 1 infection in calves with high levels of passive antibodies lacking gE antibodies. Vet. Microbiol. 2001, 82, 211–222. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; Moerman, A.; Madic, J.; Weerdmeester, K.; Maris-Veldhuis, M.; Rijsewijk, F.A.; van Oirschot, J.T. An inactivated vaccine based on a glycoprotein E-negative strain of bovine herpesvirus 1 induces protective immunity and allows serological differentiation. Vaccine 1995, 13, 342–346. [Google Scholar] [CrossRef]
- Bosch, J.C.; Kaashoek, M.J.; Kroese, A.H.; van Oirschot, J.T. An attenuated bovine herpesvirus 1 marker vaccine induced a better protection than two inactivated marker vaccines. Vet. Microbiol. 1996, 52, 223–234. [Google Scholar] [CrossRef]
- Kaashoek, M.J.; Rijsewijk, F.A.; Ruuls, R.C.; Keil, G.M.; Thiry, E.; Pastoret, P.P.; van Oirschot, J.T. Virulence, immunogenicity and reactivation of bovine herpesvirus 1 mutants with a deletion in the gC, gG, gI, gE, or in both the gI and gE gene. Vaccine 1998, 16, 802–809. [Google Scholar] [CrossRef]
- Straub, O.C. Experiences with two BHV-1 marker vaccines. Bov. Pract. 1997, 31, 46–48. [Google Scholar]
- Mars, M.H.; De Jong, M.C.; van Oirschot, J.T. A gE-negative bovine herpesvirus 1 vaccine strain is not re-excreted nor transmitted in an experimental cattle population after corticosteroid treatments. Vaccine 2000, 18, 1975–1981. [Google Scholar] [CrossRef]
- Davies, D.H.; Carmicheal, L.E. Role of cell-mediated immunity in the recovery of cattle from primary and recurrent infections with infectious bovine rhinotracheitis virus. Infect. Immune 1973, 8, 510–518. [Google Scholar]
- Bosch, J.C.; Kaashoek, M.J.; Van Oirscot, J.T. Inactivated bovine herpesvirus 1 (BHV1) marker vaccines are more efficacious in reducing virus excretion after reactivation than a live marker vaccine. Vaccine 1997, 15, 1512–1517. [Google Scholar] [CrossRef]
- Makoschey, B.; Beer, M. A live bovine herpesvirus-1 marker vaccine is not shed after intramuscular vaccination. Berl. Munch. Tierarztl. Wochenschr. 2007, 120, 480–482. [Google Scholar]
- Nagashunmugam, T.; Lubinski, J.; Wang, L.; Goldstein, L.T.; Weeks, B.S.; Sundaresan, P.; Kang, E.H.; Dubin, G.; Friedman, H.M. In vivo immune evasion mediated by the herpes simplex type 1 immunoglobulin G fc receptor. J. Virol. 1998, 72, 5351–5359. [Google Scholar] [PubMed]
- Favoreel, H.W.; Nauwynck, H.J.; Van Oostveldt, P.; Mettenleiter, T.C.; Pensaert, M.B. Antibody-induced and cytoskeleton-mediated redistribution and shedding of viral glycoproteins, expressed on pseudorabies virus-infected cells. J. Virol. 1999, 71, 8254–8261. [Google Scholar]
- Van Engelenburg, F.A.; Kaashoek, M.J.; van Oirschot, J.T.; Rijsewijk, F.A. A glycoprotein E deletion mutant of bovine herpesvirus 1 infects the same limited number of tissues in calves as wild-type virus, but for a shorter period. J. Gen. Virol. 1995, 76, 2387–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litwin, V.; Jackson, W.; Grose, C. Receptor properties of two varicella-zoster virus glycoproteins, gpI and gpIV, homologous to herpes simplex virus gE and gI. J. Virol. 1992, 66, 3643–3651. [Google Scholar] [PubMed]
- Johnson, D.C.; Frame, M.C.; Ligas, M.W.; Cross, A.M.; Stow, N.D. Herpes simplex virus immunoglobulin G Fc receptor activity depends on a complex of two viral glycoproteins, gE and gI. J. Virol. 1988, 62, 1347–1354. [Google Scholar]
- Ndjamen, B.; Farley, A.H.; Lee, T.; Fraser, S.E.; Biorkman, P.J. The Herpes Virus Fc Receptor gE-gI Mediates Antibody Bipolar Bridging to Clear Viral Antigens from the Cell Surface. PLoS Pathog. 2014, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Whitbeck, J.C.; Knappa, A.C.; Enquist, L.W.; Lawrence, W.C.; Bello, L.J. Synthesis, processing, and oligomerization of bovine herpesvirus 1 gE and gI membrane proteins. J. Virol. 1996, 70, 7878–7884. [Google Scholar]



{kind=link}
{kind=link}
| Name of Vaccine (Company) | Active Substance | Vaccine Strain | Dose | Route | Marketing Authorization Numbers |
|---|---|---|---|---|---|
| Hiprabovis IBR Marker Live (Hipra) | Live gE-, tk-, double-gene deleted BoHV-1 virus | Ceddel 106.3–107.30 CCID50 a | Single 2 mL | i.m. | EMEA/V/C/000158 |
| Cattlemarker IBR Inactivated (Zoetis) | gE-, inactivated virus | Difivac gE-, ≥5.5 log2 b | Single 2 mL | s.c. | EMEA/V/A/115 |
| Bayovac IBR Marker Vivum (Bayer) | gE -, modified live (attenuated) virus | Divifac 105 TCID50 (min)–107 TCID50 (max) c | Single 2 mL | i.n., i.m. | EMEA/V/A/023b/001 |
| Bovalto Ibraxion Inactivated IBR virus (Merial) | gE-, inactivated IBR virus | 0.75 VN.U d | Single 2 mL | s.c. | EMEA/V/C/000051 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrini, S.; Iscaro, C.; Righi, C. Antibody Responses to Bovine Alphaherpesvirus 1 (BoHV-1) in Passively Immunized Calves. Viruses 2019, 11, 23. https://doi.org/10.3390/v11010023
Petrini S, Iscaro C, Righi C. Antibody Responses to Bovine Alphaherpesvirus 1 (BoHV-1) in Passively Immunized Calves. Viruses. 2019; 11(1):23. https://doi.org/10.3390/v11010023
Chicago/Turabian StylePetrini, Stefano, Carmen Iscaro, and Cecilia Righi. 2019. "Antibody Responses to Bovine Alphaherpesvirus 1 (BoHV-1) in Passively Immunized Calves" Viruses 11, no. 1: 23. https://doi.org/10.3390/v11010023
APA StylePetrini, S., Iscaro, C., & Righi, C. (2019). Antibody Responses to Bovine Alphaherpesvirus 1 (BoHV-1) in Passively Immunized Calves. Viruses, 11(1), 23. https://doi.org/10.3390/v11010023