A Comprehensive RNA-seq Analysis of Human Bocavirus 1 Transcripts in Infected Human Airway Epithelium



Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Airway Epithelium Cultured at an Air-Liquid Interface (HAE-ALI)
2.2. Virus Infection and Quantification of Apically Released Virions
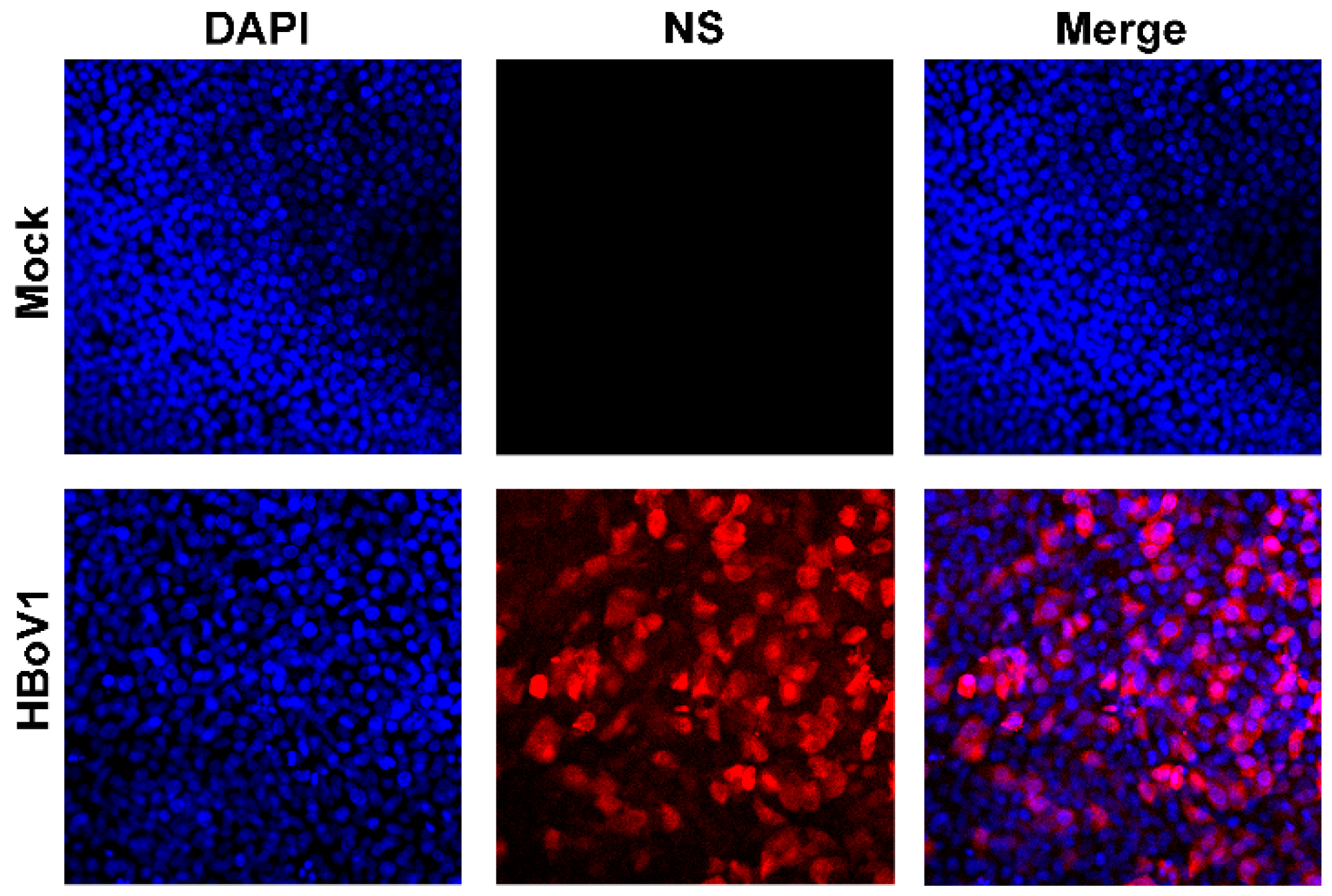
2.3. Immunofluorescence Analysis
2.4. RNA Extraction
2.5. mRNA-seq and Small RNA-seq
2.6. mRNA-seq Read Mapping and Junction Analysis
2.7. Detection of Polyadenylation Sites
2.8. HBoV1 RNA Transcripts Assembling
2.9. Mapping of Small RNA-seq Reads and Transcript Abundance Estimation
3. Results
3.1. Virus Infection
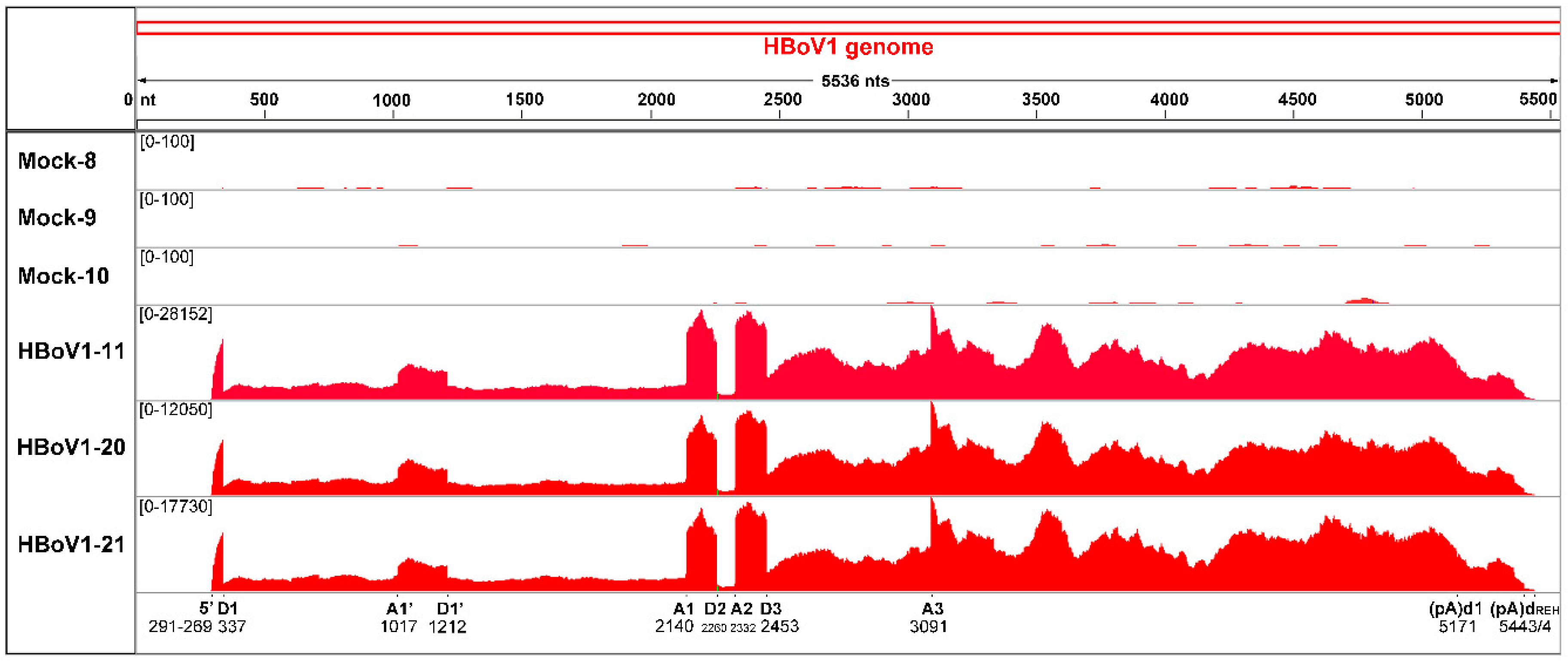
3.2. Illumina mRNA-seq Next-Generation Sequencing
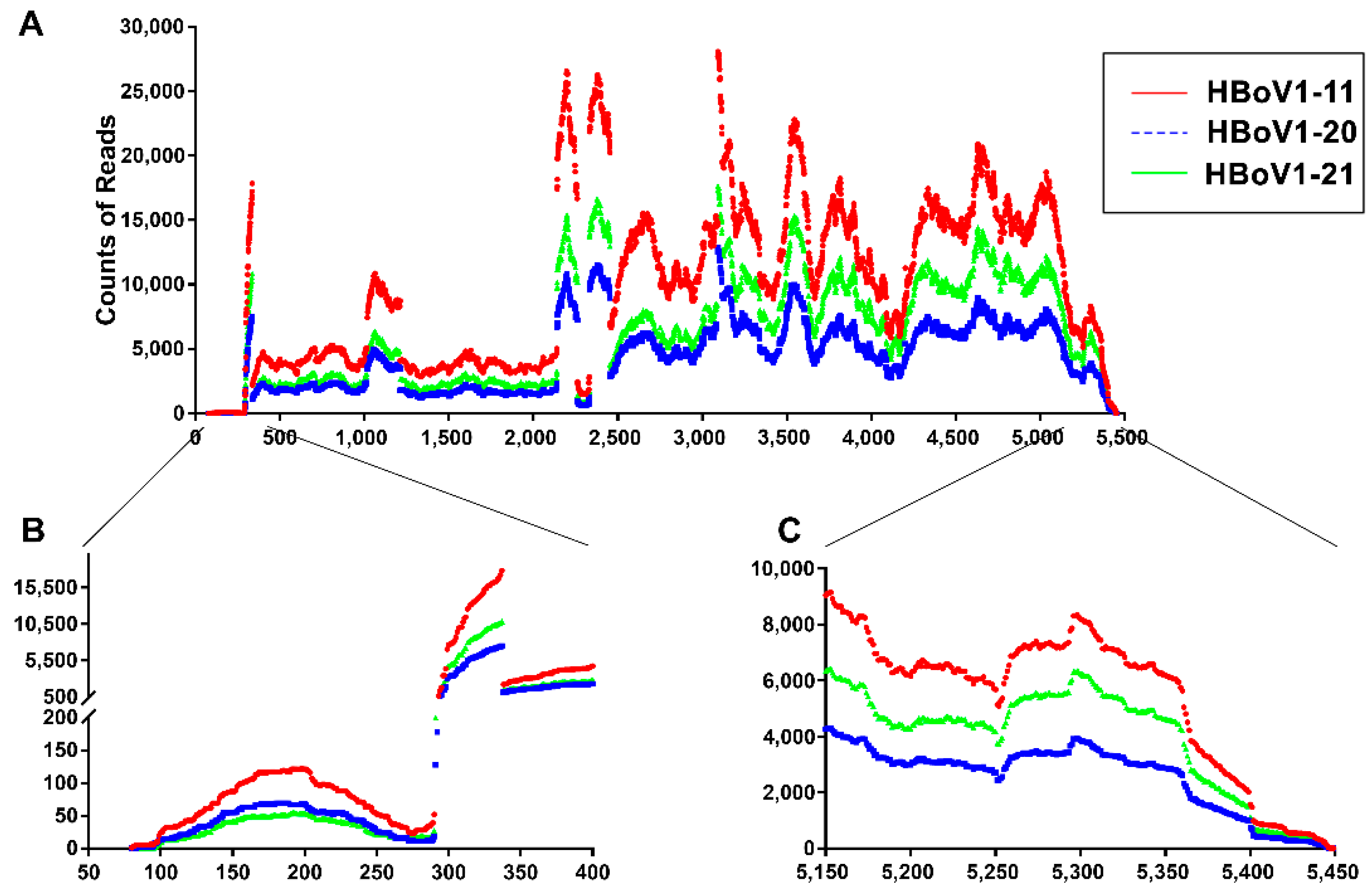
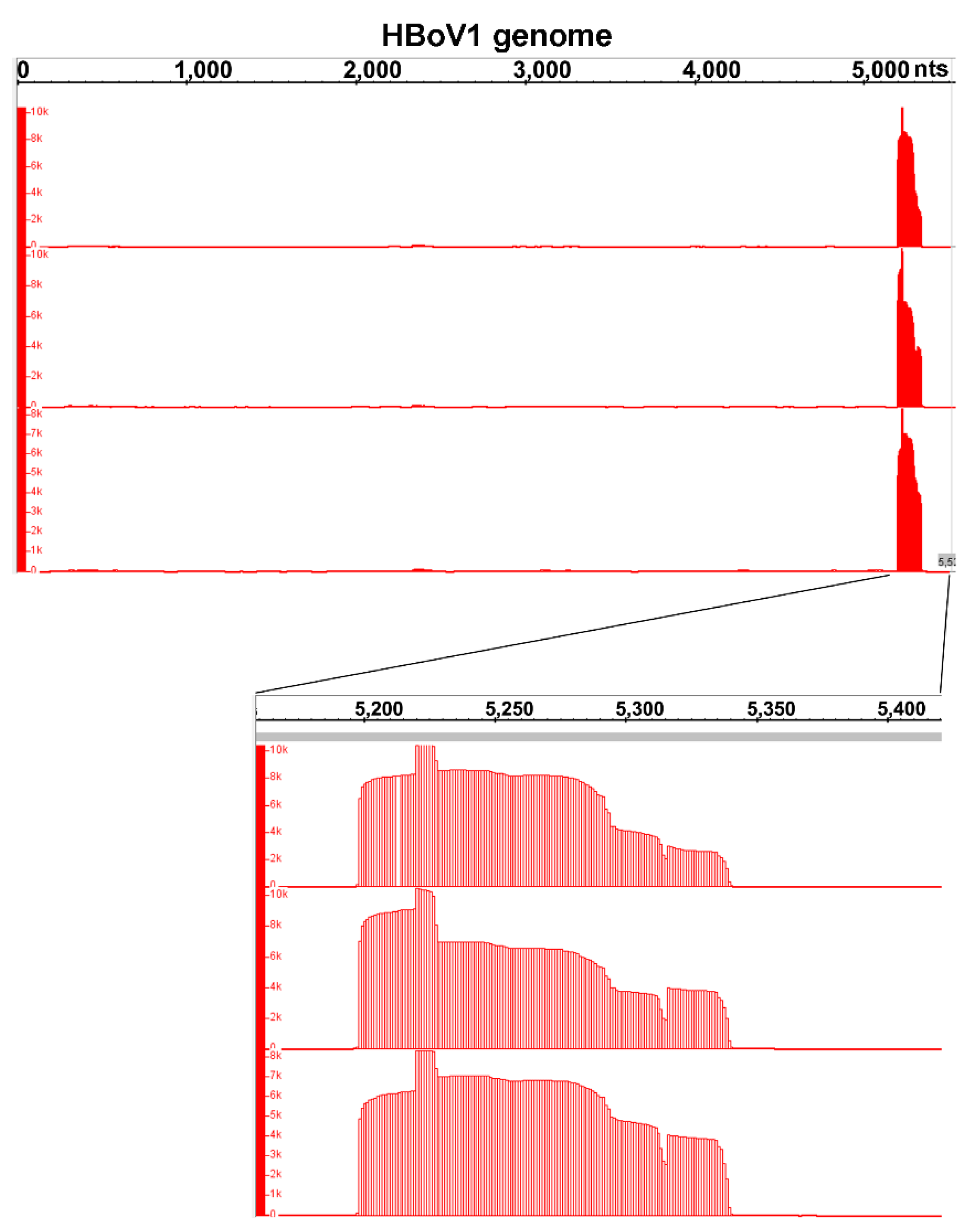
3.3. mRNA-seq Reads Mapping on the HBoV1 Plus Strand
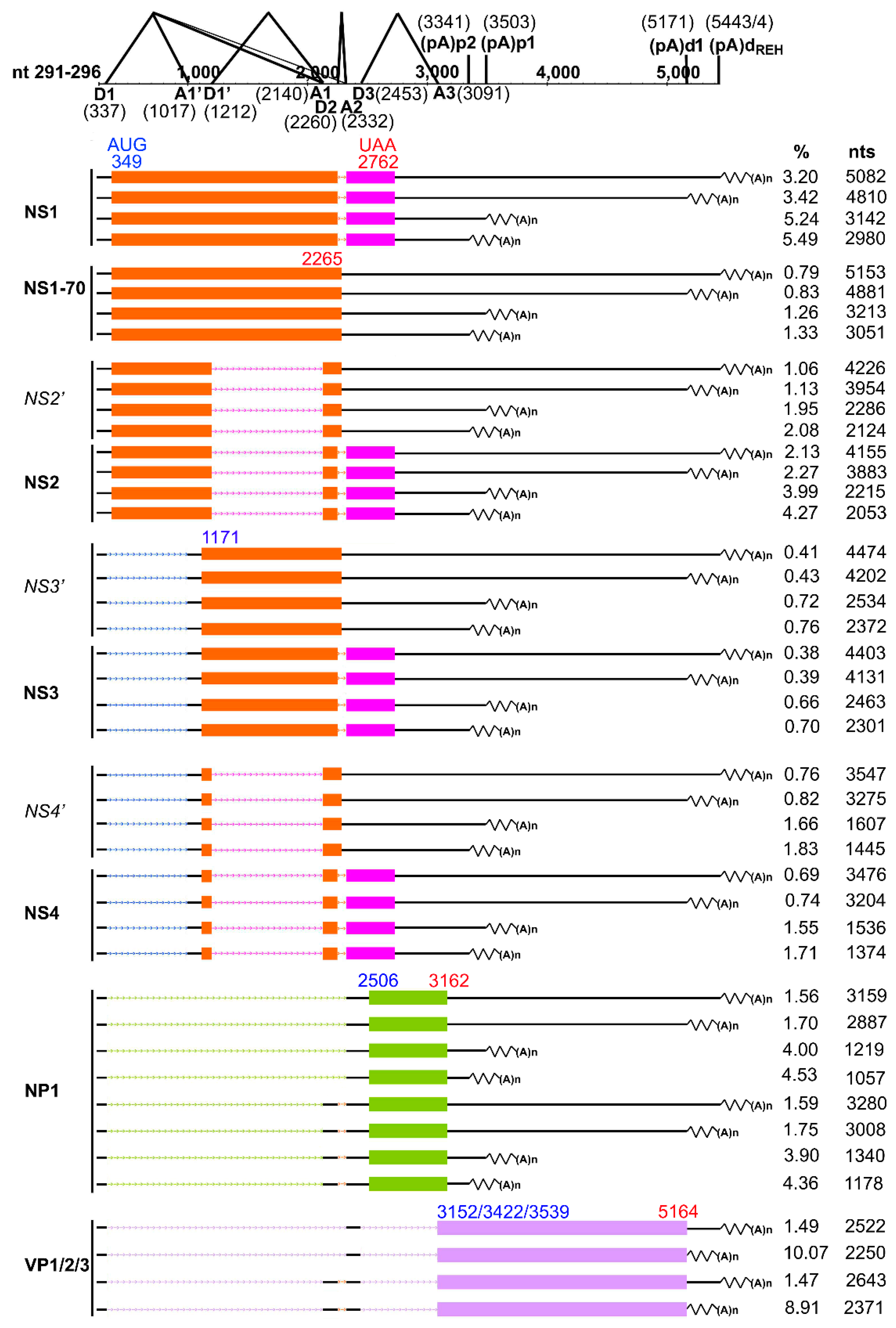
3.4. Analyses of Alternative Splicing of the HBoV1 pre-mRNA
3.5. Analyses of Alternative Polyadenylation in the HBoV1 pre-mRNA
3.6. Summarized HBoV1 mRNA Transcripts by RNA-seq
3.7. Small RNA-seq Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allander, T.; Jartti, T.; Gupta, S.; Niesters, H.G.; Lehtinen, P.; Osterback, R.; Vuorinen, T.; Waris, M.; Bjerkner, A.; Tiveljung-Lindell, A.; et al. Human bocavirus and acute wheezing in children. Clin. Infect. Dis. 2007, 44, 904–910. [Google Scholar] [CrossRef]
- Qiu, J.; Söderlund-Venermo, M.; Young, N.S. Human parvoviruses. Clin. Microbiol. Rev. 2017, 30, 43–113. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zeng, A.; Yang, N.; Lin, H.; Yang, E.; Wang, S.; Pintel, D.; Qiu, J. Quantification of human bocavirus in lower respiratory tract infections in China. Infect. Agent Cancer 2007, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Jartti, T.; Hedman, K.; Jartti, L.; Ruuskanen, O.; Allander, T.; Söderlund-Venermo, M. Human bocavirus-the first 5 years. Rev. Med. Virol. 2011, 22, 46–64. [Google Scholar] [CrossRef] [PubMed]
- Don, M.; Söderlund-Venermo, M.; Valent, F.; Lahtinen, A.; Hedman, L.; Canciani, M.; Hedman, K.; Korppi, M. Serologically verified human bocavirus pneumonia in children. Pediatr. Pulmonol. 2010, 45, 120–126. [Google Scholar] [CrossRef]
- Garcia-Garcia, M.L.; Calvo, C.; Falcon, A.; Pozo, F.; Perez-Brena, P.; De Cea, J.M.; Casas, I. Role of emerging respiratory viruses in children with severe acute wheezing. Pediatr. Pulmonol. 2010, 45, 585–591. [Google Scholar] [CrossRef]
- Liu, W.K.; Liu, Q.; Chen, D.H.; Tan, W.P.; Cai, Y.; Qiu, S.Y.; Xu, D.; Li, C.; Li, X.; Lin, Z.S.; et al. Epidemiology of HBoV1 infection and relationship with meteorological conditions in hospitalized pediatric patients with acute respiratory illness: A 7-year study in a subtropical region. BMC Infect. Dis. 2018, 18, 329–3225. [Google Scholar] [CrossRef]
- Praznik, A.; Vinsek, N.; Prodan, A.; Erculj, V.; Pokorn, M.; Mrvic, T.; Paro, D.; Krivec, U.; Strle, F.; Petrovec, M.; et al. Risk factors for bronchiolitis severity: A retrospective review of patients admitted to the university hospital from central region of Slovenia. Influenza Other Respir Viruses 2018, 12, 765–771. [Google Scholar] [CrossRef]
- Ursic, T.; Steyer, A.; Kopriva, S.; Kalan, G.; Krivec, U.; Petrovec, M. Human bocavirus as the cause of a life-threatening infection. J. Clin. Microbiol. 2011, 49, 1179–1181. [Google Scholar] [CrossRef]
- Korner, R.W.; Soderlund-Venermo, M.; van Koningsbruggen-Rietschel, S.; Kaiser, R.; Malecki, M.; Schildgen, O. Severe human bocavirus infection, Germany. Emerg. Infect. Dis. 2011, 17, 2303–2305. [Google Scholar] [CrossRef]
- Edner, N.; Castillo-Rodas, P.; Falk, L.; Hedman, K.; Soderlund-Venermo, M.; Allander, T. Life-threatening respiratory tract disease with human bocavirus-1 infection in a four-year-old child. J. Clin. Microbiol. 2011, 50, 531–532. [Google Scholar] [CrossRef] [PubMed]
- Jula, A.; Waris, M.; Kantola, K.; Peltola, V.; Söderlund-Venerm, M.; Hedman, K.; Ruuskanen, O. Primary and secondary human bocavirus 1 infections in a family, Finland. Emerg. Infect. Dis. 2013, 19, 1328–1331. [Google Scholar] [CrossRef] [PubMed]
- Ursic, T.; Krivec, U.; Kalan, G.; Petrovec, M. Fatal human bocavirus infection in an 18-month-old child with chronic lung disease of prematurity. Pediatr. Infect. Dis. J. 2015, 34, 111–112. [Google Scholar] [CrossRef]
- Eskola, V.; Xu, M.; Soderlund-Venermo, M. Severe Lower Respiratory Tract Infection Caused by Human Bocavirus 1 in an Infant. Pediatr. Infect. Dis. J. 2017, 36, 1107–1108. [Google Scholar] [CrossRef] [PubMed]
- Schlaberg, R.; Queen, K.; Simmon, K.; Tardif, K.; Stockmann, C.; Flygare, S.; Kennedy, B.; Voelkerding, K.; Bramley, A.; Zhang, J.; et al. Viral Pathogen Detection by Metagenomics and Pan Viral Group PCR in Children with Pneumonia Lacking Identifiable Etiology. J. Infect. Dis. 2017, 215, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Dijkman, R.; Koekkoek, S.M.; Molenkamp, R.; Schildgen, O.; van der Hoek, L. Human bocavirus can be cultured in differentiated human airway epithelial cells. J. Virol. 2009, 83, 7739–7748. [Google Scholar] [CrossRef]
- Huang, Q.; Deng, X.; Yan, Z.; Cheng, F.; Luo, Y.; Shen, W.; Lei-Butters, D.C.; Chen, A.Y.; Li, Y.; Tang, L.; et al. Establishment of a reverse genetics system for studying human bocavirus in human airway epithelia. PLoS Pathog. 2012, 8, e1002899. [Google Scholar] [CrossRef]
- Deng, X.; Yan, Z.; Luo, Y.; Xu, J.; Cheng, Y.; Li, Y.; Engelhardt, J.; Qiu, J. In vitro modeling of human bocavirus 1 infection of polarized primary human airway epithelia. J. Virol. 2013, 87, 4097–4102. [Google Scholar] [CrossRef]
- Deng, X.; Li, Y.; Qiu, J. Human bocavirus 1 infects commercially available primary human airway epithelium cultures productively. J. Virol. Methods 2014, 195, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Yan, Z.; Cheng, F.; Engelhardt, J.F.; Qiu, J. Replication of an Autonomous Human Parvovirus in Non-dividing Human Airway Epithelium Is Facilitated through the DNA Damage and Repair Pathways. PLoS Pathog. 2016, 12, e1005399. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, A.Y.; Cheng, F.; Guan, W.; Johnson, F.B.; Qiu, J. Molecular characterization of infectious clones of the minute virus of canines reveals unique features of bocaviruses. J. Virol. 2009, 83, 3956–3967. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Cheng, F.; Johnson, F.B.; Pintel, D. The transcription profile of the bocavirus bovine parvovirus is unlike those of previously characterized parvoviruses. J. Virol. 2007, 81, 12080–12085. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.Y.; Cheng, F.; Lou, S.; Luo, Y.; Liu, Z.; Delwart, E.; Pintel, D.; Qiu, J. Characterization of the gene expression profile of human bocavirus. Virology 2010, 403, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; Deng, X.; Zou, W.; Cheng, F.; Engelhardt, J.F.; Yan, Z.; Qiu, J. Identification and Functional Analysis of Novel Non-structural Proteins of Human Bocavirus 1. J. Virol. 2015, 89, 10097–10109. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Cheng, F.; Shen, W.; Engelhardt, J.F.; Yan, Z.; Qiu, J. Nonstructural Protein NP1 of Human Bocavirus 1 Plays a Critical Role in the Expression of Viral Capsid Proteins. J. Virol. 2016, 90, 4658–4669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Shen, W.; Cheng, F.; Deng, X.; Engelhardt, J.F.; Yan, Z.; Qiu, J. Parvovirus Expresses a Small Noncoding RNA That Plays an Essential Role in Virus Replication. J. Virol. 2017, 91, e02375-16. [Google Scholar] [CrossRef]
- Flotte, T.R.; Afione, S.A.; Solow, R.; Drumm, M.L.; Markakis, D.; Guggino, W.B.; Zeitlin, P.L.; Carter, B.J. Expression of the cystic fibrosis transmembrane conductance regulator from a novel adeno-associated virus promoter. J. Biol. Chem. 1993, 268, 3781–3790. [Google Scholar]
- Qiu, J.; Nayak, R.; Tullis, G.E.; Pintel, D.J. Characterization of the transcription profile of adeno-associated virus type 5 reveals a number of unique features compared to previously characterized adeno-associated viruses. J. Virol. 2002, 76, 12435–12447. [Google Scholar] [CrossRef]
- Murthy, K.G.; Manley, J.L. The 160-kD subunit of human cleavage-polyadenylation specificity factor coordinates pre-mRNA 3′-end formation. Genes Dev. 1995, 9, 2672–2683. [Google Scholar] [CrossRef]
- Stutika, C.; Mietzsch, M.; Gogol-Doring, A.; Weger, S.; Sohn, M.; Chen, W.; Heilbronn, R. Comprehensive Small RNA-Seq of Adeno-Associated Virus (AAV)-Infected Human Cells Detects Patterns of Novel, Non-Coding AAV RNAs in the Absence of Cellular miRNA Regulation. PLoS ONE 2016, 11, e0161454. [Google Scholar] [CrossRef]
- Sukhu, L.; Fasina, O.; Burger, L.; Rai, A.; Qiu, J.; Pintel, D.J. Characterization of the nonstructural proteins of the bocavirus minute virus of canines. J. Virol. 2013, 87, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Fasina, O.O.; Dong, Y.; Pintel, D.J. NP1 Protein of the Bocaparvovirus Minute Virus of Canines Controls Access to the Viral Capsid Genes via Its Role in RNA Processing. J. Virol. 2015, 90, 1718–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasina, O.O.; Stupps, S.; Figueroa-Cuilan, W.; Pintel, D.J. The minute virus of canines (MVC) NP1 protein governs the expression of a subset of essential NS proteins via its role in RNA processing. J. Virol. 2017, 91, e00260-17. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Fasina, O.O.; Pintel, D.J. The Human Bocavirus 1 NP1 Protein Is a Multifunctional Regulator of Viral RNA Processing. J. Virol. 2018, 92, e01187-18. [Google Scholar] [CrossRef]
- Zhao, J.; Hyman, L.; Moore, C. Formation of mRNA 3′ ends in eukaryotes: Mechanism, regulation, and interrelationships with other steps in mRNA synthesis. Microbiol. Mol. Biol. Rev. 1999, 63, 405–445. [Google Scholar] [PubMed]
- Hao, S.; Zhang, J.; Chen, Z.; Xu, H.; Wang, H.; Guan, W. Alternative Polyadenylation of Human Bocavirus at Its 3′ End Is Regulated by Multiple Elements and Affects Capsid Expression. J. Virol. 2017, 91, e02026-16. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Total Reads | No. (%) of Mapped Reads | No. (%) of Human Reads | No. (%) of HBoV1 reads | ||
|---|---|---|---|---|---|---|
| Plus Strand | Minus Strand | Ambiguous | ||||
| Mock-8 | 92,255,534 | 69,845,826 (75.71) | 69,845,805 (75.71) | 19 (0.00) | 2 (0.00) | 0 (0.00) |
| Mock-9 | 83,459,160 | 63,548,966 (76.14) | 63,548,950 (76.14) | 15 (0.00) | 1 (0.00) | 0 (0.00) |
| Mock-10 | 95,389,110 | 72,602,804 (76.11) | 72,602,787 (76.11) | 17 (0.00) | 0 (0.00) | 0 (0.00) |
| HBoV1-11 | 86,814,188 | 64,362,968 (74.14) | 63,865,552 (73.57) | 474,990 (0.55) | 21,580 (0.02) | 846 (0.00) |
| HBoV1-20 | 90,567,600 | 68,650,250 (75.80) | 68,429,393 (75.56) | 208,998 (0.23) | 9458 (0.01) | 56 (0.00) |
| HBoV1-21 | 7,175,830 | 54,487,120 (75.93) | 54,162,045 (75.48) | 309,318 (0.43) | 12,952 (0.02) | 142 (0.00) |
| Donor | Acceptor | Splicing Events | Strand | HBoV1-11 | HBoV1-20 | HBoV1-21 | Ratio # (%) |
|---|---|---|---|---|---|---|---|
| 337 § | 1016 | D1/A1’ | + | 2349 | 1081 | 1334 | 85.10 |
| 337 | 1060 | + | 35 | 0 | 0 | ||
| 337 | 1108 | + | 327 | 144 | 160 | 14.90 | |
| 337 | 1118 | + | 101 | 0 | 67 | ||
| 337 | 2139 | D1/A1 | + | 7962 | 2985 | 4698 | 97.82 |
| 337 | 2198 | + | 158 | 83 | 107 | 2.18 | |
| 337 | 2331 | D1/A2 | + | 4734 | 2084 | 3124 | 99.4 |
| 337 | 861 | + | 30 | 0 | 0 | 0.60 | |
| 337 | 998 | + | 0 | 0 | 30 | ||
| 1212 | 2139 | D1’/A1 | + | 3818 | 1462 | 1814 | 95.12 |
| 1212 | 2198 | + | 0 | 89 | 111 | 4.88 | |
| 1201 | 2187 | + | 0 | 52 | 112 | ||
| 1212 | 2331 | D1’/A2 | + | 95 | 71 | 46 | 14.73 |
| 1212 | 2372 | + | 414 | 0 | 0 | ||
| 1630 | 2139 | + | 644 | 0 | 0 | 85.27 | |
| 2250 | 2362 | + | 169 | 0 | 0 | ||
| 2260 | 2331 | D2/A2 | + | 14,175 | 6222 | 8782 | 98.86 |
| 2423 | 2576 | + | 228 | 107 | 0 | 1.14 | |
| 2453 | 3090 | D3/A3 | + | 12,973 | 5794 | 9006 | 99.44 |
| 3180 | 3865 | + | 0 | 0 | 156 | 0.56 |
| Transcripts (Splicing form) | mRNA Polyadenylation Sites | |||
|---|---|---|---|---|
| (pA)p1 | (pA)p2 | (pA)d1 | (pA)dREH | |
| NS1 (D2A2) | 15,342 | 16,071 | 10,029 | 9364 |
| NS70 | 3686 | 3883 | 2440 | 2303 |
| NS2 (D1’A1/D2A2) | 11,700 | 12,500 | 6646 | 6243 |
| NS2 (D1’A1) | 5721 | 6105 | 3313 | 3098 |
| NS3 (D1A1’/D2A2) | 1938 | 2061 | 1155 | 1101 |
| NS3 (D1A1’) | 2102 | 2227 | 1268 | 1190 |
| NS4 (D1A1’/D1’A1/D2A2) | 4549 | 4998 | 2181 | 2007 |
| NS4 (D1A1’/D1’A1) | 4870 | 5350 | 2397 | 2213 |
| NP1 (D1A1/D2A2) | 11,434 | 12,782 | 5114 | 4667 |
| NP1 (D1A2) | 11,724 | 13,269 | 4971 | 4580 |
| VP (D1A1/D2A2/D3A3) | NA | NA | 26,107 | 4306 |
| VP (D1A2/D3A3) | NA | NA | 29,504 | 4378 |
| Start | End | Length | Strand | Sample 1 (Reads) | Sample 2 (Reads) | Sample 3 (Reads) | Total (Reads) |
|---|---|---|---|---|---|---|---|
| 5199 | 5228 | 29 | + | 373 | 664 | 277 | 1314 |
| 5317 | 5340 | 23 | + | 316 | 532 | 421 | 1269 |
| 5221 | 5315 | 94 | + | 356 | 284 | 306 | 946 |
| 5221 | 5314 | 93 | + | 198 | 326 | 362 | 886 |
| 5199 | 5295 | 96 | + | 415 | 235 | 179 | 829 |
| 5199 | 5229 | 30 | + | 253 | 395 | 165 | 813 |
| 5199 | 5293 | 94 | + | 386 | 202 | 158 | 746 |
| 5199 | 5221 | 22 | + | 128 | 297 | 216 | 641 |
| 5200 | 5228 | 28 | + | 131 | 184 | 85 | 400 |
| 5317 | 5336 | 19 | + | 97 | 151 | 129 | 377 |
| 5317 | 5339 | 22 | + | 90 | 168 | 108 | 366 |
| 5199 # | 5339 | 140 | + | 37 | 94 | 148 | 279 |
| 5221 | 5317 | 96 | + | 135 | 61 | 80 | 276 |
| 5199 | 5294 | 95 | + | 109 | 88 | 71 | 268 |
| 5317 | 5341 | 24 | + | 63 | 122 | 82 | 267 |
| 5199 | 5290 | 91 | + | 111 | 72 | 61 | 244 |
| 5199 | 5227 | 28 | + | 76 | 120 | 40 | 236 |
| 5221 | 5313 | 92 | + | 45 | 69 | 118 | 232 |
| 5199 | 5338 | 139 | + | 21 | 67 | 134 | 222 |
| 5199 | 5340 | 141 | + | 34 | 72 | 113 | 219 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, W.; Xiong, M.; Deng, X.; Engelhardt, J.F.; Yan, Z.; Qiu, J. A Comprehensive RNA-seq Analysis of Human Bocavirus 1 Transcripts in Infected Human Airway Epithelium. Viruses 2019, 11, 33. https://doi.org/10.3390/v11010033
Zou W, Xiong M, Deng X, Engelhardt JF, Yan Z, Qiu J. A Comprehensive RNA-seq Analysis of Human Bocavirus 1 Transcripts in Infected Human Airway Epithelium. Viruses. 2019; 11(1):33. https://doi.org/10.3390/v11010033
Chicago/Turabian StyleZou, Wei, Min Xiong, Xuefeng Deng, John F. Engelhardt, Ziying Yan, and Jianming Qiu. 2019. "A Comprehensive RNA-seq Analysis of Human Bocavirus 1 Transcripts in Infected Human Airway Epithelium" Viruses 11, no. 1: 33. https://doi.org/10.3390/v11010033
APA StyleZou, W., Xiong, M., Deng, X., Engelhardt, J. F., Yan, Z., & Qiu, J. (2019). A Comprehensive RNA-seq Analysis of Human Bocavirus 1 Transcripts in Infected Human Airway Epithelium. Viruses, 11(1), 33. https://doi.org/10.3390/v11010033