The Phylogeny and Pathogenesis of Sacbrood Virus (SBV) Infection in European Honey Bees, Apis mellifera


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bee Sample Collection and SBV Inoculum Preparation
2.3. RNA Extraction
2.4. RT-PCR and RACE
2.5. Phylogenetic Analysis
2.6. Laboratory Induced Sbv Pathogenic Infection
2.7. Cold Stress on Survivorship of SBV-Infected Larvae
3. Results
3.1. Complete Genome Sequences of U.S. Strains of SBV
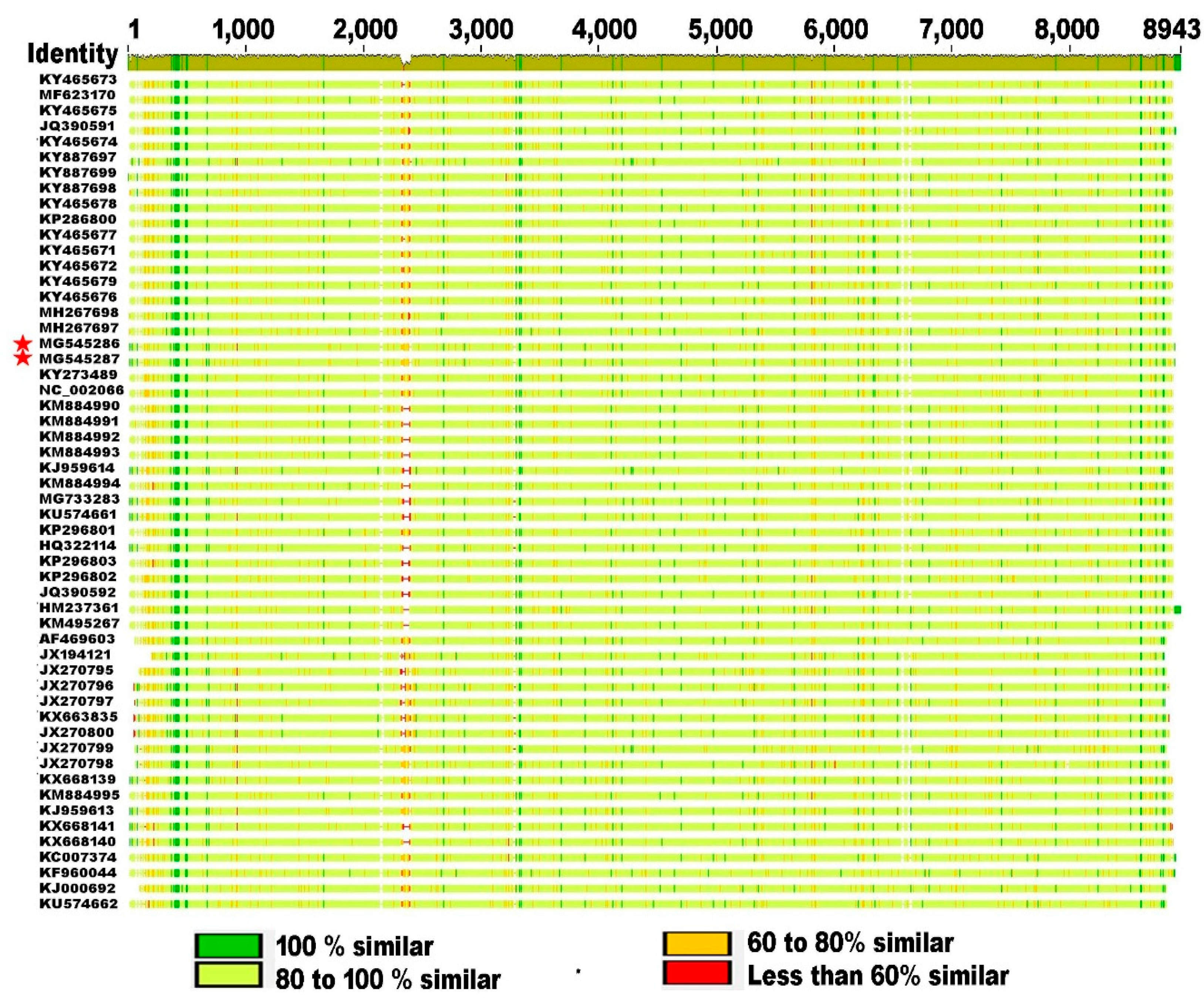
3.2. Genetic Relationship of SBV Strains Worldwide
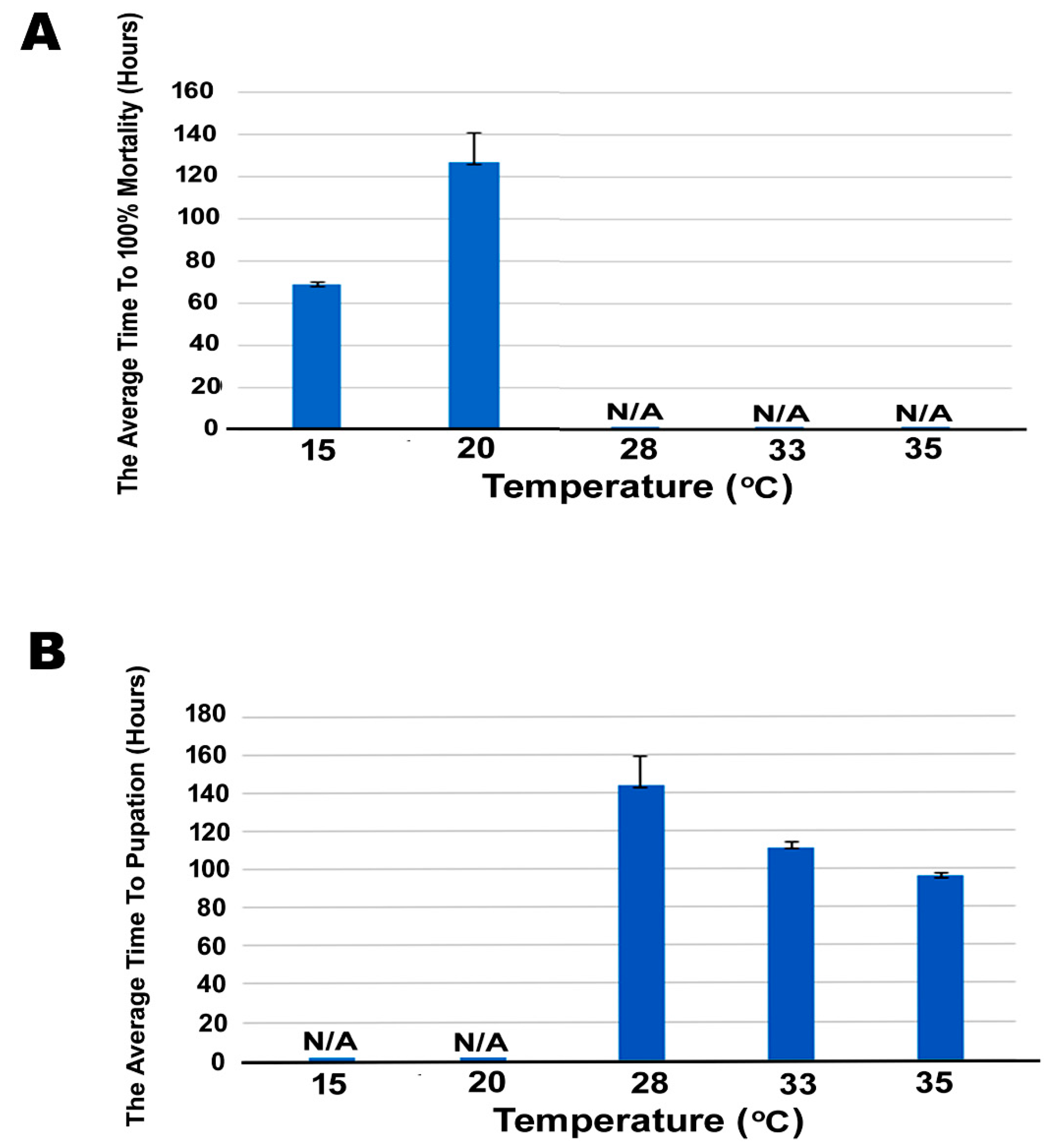
3.3. Effect of Cold Stress on Mortality Of SBV-Infected Larvae
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Potts, S.G.; Petanidou, T.; Roberts, S.; O’Toole, C.; Hulbert, A.; Willmer, P. Plant-pollinator biodiversity and pollination services in a complex Mediterranean landscape. Biol. Conserv. 2006, 129, 519–529. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef] [Green Version]
- Aizen, M.A.; Harder, L.D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Levy, S. The pollinator crisis: What’s best for bees. Nature 2011, 479, 164–165. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef]
- Kulhanek, K.; Steinhauer, N.; Rennich, N.K.; Caron, D.M.; Sagili, R.R.; Pettis, J.P.; Ellis, J.D.; Wilson, M.J.; Wilkes, J.T.; Tarpy, D.R.; et al. A national survey of managed honey bee 2015–2016 annual colony losses in the USA. J. Apic. Res. 2017, 56, 328–340. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Vanbergen, A.J.; Insect Pollinators Initiative. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; vanEngelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef] [PubMed]
- Spivak, M.; Mader, E.; Vaughan, M.; Euliss, N.H., Jr. The plight of the bees. Environ. Sci. Technol. 2011, 45, 34–38. [Google Scholar] [CrossRef] [PubMed]
- White, G.F. Sacbrood, a Disease of Bees; US Department of Agriculture, Bureau of Entomology: Washington, DC, USA, 1913.
- Allen, M.; Ball, B. The incidence and world distribution of honey bee viruses. Bee World 1996, 77, 141–162. [Google Scholar] [CrossRef]
- Chen, Y.P.; Siede, R. Honey bee viruses. Adv. Virus Res. 2007, 70, 33–80. [Google Scholar] [PubMed]
- Bailey, L.; Gibbs, A.J.; Woods, R.D. Sacbrood virus of the larval honeybee (Apis mellifera Linnaeus). Virology 1964, 23, 425–429. [Google Scholar] [CrossRef]
- Shen, M.; Cui, L.; Ostiguy, N.; Cox-Foster, D. Intricate transmission routes and interactions between picorna-like viruses (Kashmir bee virus and sacbrood virus) with the honeybee host and the parasitic varroa mite. J. Gen. Virol. 2005, 86, 2281–2289. [Google Scholar] [CrossRef] [Green Version]
- Grabensteiner, E.; Ritter, W.; Carter, M.J.; Davison, S.; Pechhacker, H.; Kolodziejek, J.; Boecking, O.; Derakhshifar, I.; Moosbeckhofer, R.; Licek, E.; et al. Sacbrood virus of the honeybee (Apis mellifera): Rapid identification and phylogenetic analysis using reverse transcription-PCR. Clin. Diagn. Lab. Immunol. 2001, 8, 93–104. [Google Scholar] [CrossRef]
- Anderson, D.L.; Gibbs, A.J. Inapparent virus infections and their interactions in pupae of the honeybee (Apis mellifera L.) in Australia. J. Gen. Virol. 1988, 69, 1617–1625. [Google Scholar] [CrossRef]
- Berenyi, O.; Bakonyi, T.; Derakhshifar, I.; Koglberger, H.; Nowotny, N. Occurrence of six honeybee viruses in diseased Austrian apiaries. Appl. Environ. Microbiol. 2006, 72, 2414–2420. [Google Scholar] [CrossRef]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef]
- De Miranda, J.R.; Gauthier, L.; Ribière, M.; Chen, Y.P. Honey bee viruses and their effects on bee and colony health. In Honey Bee Colony Health: Challenges and Sustainable Solution; Sammataro, D., Yoder, J.A., Eds.; CRC Press Taylor&Francis Group: Boca Raton, FL, USA, 2011; pp. 71–102. [Google Scholar]
- Oldroyd, B.P. What’s killing American honey bees? PLoS Biol. 2007, 5, e168. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S96–S119. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, J.R.; Genersch, E. Deformed wing virus. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S48–S61. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef]
- Xia, X.; Zhou, B.; Wei, T. Complete genome of Chinese sacbrood virus from Apis cerana and analysis of the 3C-like cysteine protease. Virus Genes 2015, 50, 277–285. [Google Scholar] [CrossRef]
- Ghosh, R.C.; Ball, B.V.; Willcocks, M.M.; Carter, M.J. The nucleotide sequence of sacbrood virus of the honey bee: An insect picorna-like virus. J. Gen. Virol. 1999, 80 Pt 6, 1541–1549. [Google Scholar] [CrossRef]
- Choe, S.E.; Nguyen, L.T.K.; Noh, J.H.; Kweon, C.H.; Reddy, K.E.; Koh, H.B.; Chang, K.Y.; Kang, S.W. Analysis of the complete genome sequence of two Korean sacbrood viruses in the Honey bee, Apis mellifera. Virology 2012, 432, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Anderson, D.L. A novel strain of sacbrood virus of interest to world apiculture. J. Invertebr. Pathol. 2014, 118, 71–74. [Google Scholar] [CrossRef]
- Roy, C.; Vidal-Naquet, N.; Provost, B. A severe sacbrood virus outbreak in a honeybee (Apis mellifera L.) colony: A case report. Vet. Med. 2015, 60, 330–335. [Google Scholar] [CrossRef]
- Bailey, L.; Carpenter, J.M.; Woods, R.D. A strain of sacbrood virus from Apis cerana. J. Invertebr. Pathol. 1982, 39, 264–265. [Google Scholar] [CrossRef]
- Rana, B.S.; Garg, I.D.; Khurana, S.P.; Verma, L.R.; Agrawal, H.O. Thai sacbrood virus of honeybees (Apis cerana indica F) in North-west Himalayas. Indian J. Virol. 1986, 2, 127–131. [Google Scholar]
- Verma, L.R.; Rana, B.S.; Verma, S. Observations on Apis cerana colonies surviving from Thai sacbrood virus infestation. Apidologie 1990, 21, 169–174. [Google Scholar] [CrossRef]
- Zhang, J.; Feng, J.; Liang, Y.; Cheng, D.; Zhou, Z.H.; Zhang, Q.; Lu, X. Three-dimensional structure of the Chinese sacbrood bee virus. Sci. China 2001, 44, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Mingxiao, M.; Ming, L.; Jian, C.; Song, Y.; Shude, W.; Pengfei, L. Molecular and Biological Characterization of Chinese Sacbrood Virus LN Isolate. Comp. Funct. Genom. 2011, 2011, 409386. [Google Scholar] [CrossRef]
- Choe, S.E.; Nguyen, L.T.K.; Noh, J.H.; Koh, H.B.; Jean, Y.H.; Kweon, C.H.; Kang, S.W. Prevalence and distribution of six bee viruses in Korean Apis cerana populations. J. Invertebr. Pathol. 2012, 109, 330–333. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Le, T.H. Complete Genome Sequence of Sacbrood Virus Strain SBM2, Isolated from the Honeybee Apis cerana in Vietnam. Genome Announc. 2013, 1, e00076-12. [Google Scholar] [CrossRef]
- Rao, K.M.; Katna, S.; Rana, B.S.; Rana, R. Thai sacbrood and sacbrood viruses versus European foulbrood of hive bees in India—A review. J. Apic. Res. 2015, 54, 192–199. [Google Scholar]
- Li, Y.; Zeng, Z.J.; Wang, Z.L. Phylogenetic analysis of the honeybee Sacbrood virus. J. Apic. Sci. 2016, 60, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.R.; Chen, X.X.; Chen, Y.P.; Hu, F.L.; Zhang, J.L.; Lin, Z.G.; Yu, J.W.; Zheng, H.Q. Evidence of Apis cerana Sacbrood virus Infection in Apis mellifera. Appl. Environ. Microbiol. 2016, 82, 2256–2262. [Google Scholar] [CrossRef]
- Li, J.L.; Cornman, R.S.; Evans, J.D.; Pettis, J.S.; Zhao, Y.; Murphy, C.; Peng, W.J.; Wu, J.; Hamilton, M.; Boncristiani, H.F., Jr.; et al. Systemic spread and propagation of a plant-pathogenic virus in European honeybees, Apis mellifera. mBio 2014, 5, e00898-13. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Vandenberg, J.D.; Shimanuki, H. Technique for Rearing Worker Honeybees in the Laboratory. J. Apic. Res. 1987, 26, 90–97. [Google Scholar] [CrossRef]
- LME4: Linear Mixed-Effects Models Using S4 Classes. Available online: http://cran.R-project.org/package=lme4 (accessed on 5 January 2019).
- Fares, A. Factors influencing the seasonal patterns of infectious diseases. Int. J. Prev. Med. 2013, 4, 128–132. [Google Scholar] [PubMed]
- Gammon, D.B.; Mello, C.C. RNA interference-mediated antiviral defense in insects. Curr. Opin. Insect Sci. 2015, 8, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brutscher, L.M.; Flenniken, M.L. RNAi and Antiviral Defense in the Honey Bee. J. Immunol. Res. 2015, 2015, 941897. [Google Scholar] [CrossRef]
- Nylin, S.; Gotthard, K. Plasticity in Life-History Traits. Annu. Rev. Entomol. 1998, 43, 63–83. [Google Scholar] [CrossRef]
- Dodson, B.L.; Kramer, L.D.; Rasgon, J.L. Effects of larval rearing temperature on immature development and West Nile virus vector competence of Culex tarsalis. Parasites Vectors 2012, 5, 199. [Google Scholar] [CrossRef] [Green Version]
- Kronenberg, F.; Heller, H.C. Colonial thermoregulation in honey bees (Apis mellifera). J. Comp. Physiol. B 1982, 148, 65–76. [Google Scholar] [CrossRef]
- Wang, Q.; Xu, X.; Zhu, X.; Chen, L.; Zhou, S.; Huang, Z.Y.; Zhou, B. Low-Temperature Stress during Capped Brood Stage Increases Pupal Mortality, Misorientation and Adult Mortality in Honey Bees. PLoS ONE 2016, 11, e0154547. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′ to 3′) |
|---|---|
| F01 | TACGAATCGTGATTCGATTCATT |
| F908 | ACGCTAAGTGTGCGCCTAAT |
| F1149 | GATATCATCGCGCCTTTGTT |
| F1211 | GGAAGTTTGCTAGTATTTACGTG |
| F2329 | TGGAGGTAAGGGACAACCTG |
| F2408 | TGGACACTGGTGCTAAAGAAGATG |
| F3083 | GAAGCTGGGGATGATTTTGA |
| F3366 | CGCTATAGGTGGCATGCAGA |
| F3619 | GGCATGGATTGATCGAAGTT |
| F4526 | CCGGAAATGGCTCATTTAGA |
| F4648 | TACCGTAGGAGAGATGTGTTATTGT |
| F5394 | TCGAGAAGTGGTTCAGTGCC |
| F5436 | AAGTAGTCCAGTGCCCGATG |
| F5762 | TCGGATGGTGAAGATGATGA |
| F6040 | CCAGCGGTACAAAGAGGAAA |
| F6155 | CACTGGATGAGAGCGAATGA |
| F6890 | CGGTATTTTATGCGAATGATGT |
| F7627 | CCCAGCGTTCTGGAGGAAAT |
| F8256 | AACGAGGGCAAACTTGGGAA |
| F8471 | CATGGGTTTCATCCCCACGA |
| F8598 | GTCGAGCCGCTCTGTATCAA |
| F8613 | ATCAAGCGCATGGTCATGGA |
| R501 | ACTGCGCGTCTAACATTCCA |
| R683 | AACTCTGCTGTGTAGCGTCC |
| R793 | TTGTTGCGTTGGTTCGGAAG |
| R930 | GTGTTGGGTGCACACTTAGC |
| R1250 | AACGGTGACAGCATTTGCAC |
| R2450 | CCACAAGCTCCTTGTTGTGA |
| R2492 | GAATCCAAAGACTGAAAACCC |
| R3435 | CCCGTAACGGTATTCTGCAT |
| R3647 | CTGCAGTCCAACTTCGATCA |
| R3700 | GCTGCCCAAAAGTTGTAGCC |
| R3936 | ACCCCCACCAAATAAGAAGG |
| R4515 | TGCATTAAAGCTTGGGGTTC |
| R4807 | CAGGTTTAGATGTACACGAGGATG |
| R4884 | AACTTCGCAACCAACCAATC |
| R5453 | TCGGGCACTGGACTACTTCT |
| R5871 | ATTTCCCTCTCTCGCATCAA |
| R6256 | CCACTGCCGTCACTAAACCT |
| R6300 | TCATTCGCTCTCATCCAGTG |
| R6609 | ATACTCCCCACTGCCATCAC |
| R6981 | TCCTTAATGGCACGCACATA |
| R6995 | TGCATTCCCACAATAGGTCTTT |
| R8500 | TAGAATTCCAAGCCAGCGCA |
| R8863 | TAAAATGCCATATATTGATATTAATCCA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wang, T.; Evans, J.D.; Rose, R.; Zhao, Y.; Li, Z.; Li, J.; Huang, S.; Heerman, M.; Rodríguez-García, C.; et al. The Phylogeny and Pathogenesis of Sacbrood Virus (SBV) Infection in European Honey Bees, Apis mellifera. Viruses 2019, 11, 61. https://doi.org/10.3390/v11010061
Li J, Wang T, Evans JD, Rose R, Zhao Y, Li Z, Li J, Huang S, Heerman M, Rodríguez-García C, et al. The Phylogeny and Pathogenesis of Sacbrood Virus (SBV) Infection in European Honey Bees, Apis mellifera. Viruses. 2019; 11(1):61. https://doi.org/10.3390/v11010061
Chicago/Turabian StyleLi, Jianghong, Tingyun Wang, Jay D. Evans, Robyn Rose, Yazhou Zhao, Zhiguo Li, Jilian Li, Shaokang Huang, Matthew Heerman, Cristina Rodríguez-García, and et al. 2019. "The Phylogeny and Pathogenesis of Sacbrood Virus (SBV) Infection in European Honey Bees, Apis mellifera" Viruses 11, no. 1: 61. https://doi.org/10.3390/v11010061
APA StyleLi, J., Wang, T., Evans, J. D., Rose, R., Zhao, Y., Li, Z., Li, J., Huang, S., Heerman, M., Rodríguez-García, C., Banmeke, O., Brister, J. R., Hatcher, E. L., Cao, L., Hamilton, M., & Chen, Y. (2019). The Phylogeny and Pathogenesis of Sacbrood Virus (SBV) Infection in European Honey Bees, Apis mellifera. Viruses, 11(1), 61. https://doi.org/10.3390/v11010061