Equine Mx1 Restricts Influenza A Virus Replication by Targeting at Distinct Site of its Nucleoprotein


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Plasmids and Antibodies
2.3. Polymerase Reconstitution Assay/Minireplicon Assay
2.4. Co-Immunoprecipitation Assay
2.5. Virus Growth Kinetics
2.6. Measurement of Gene Expression Using RT-qPCR
2.7. Immunofluorescence Assay (IFA)
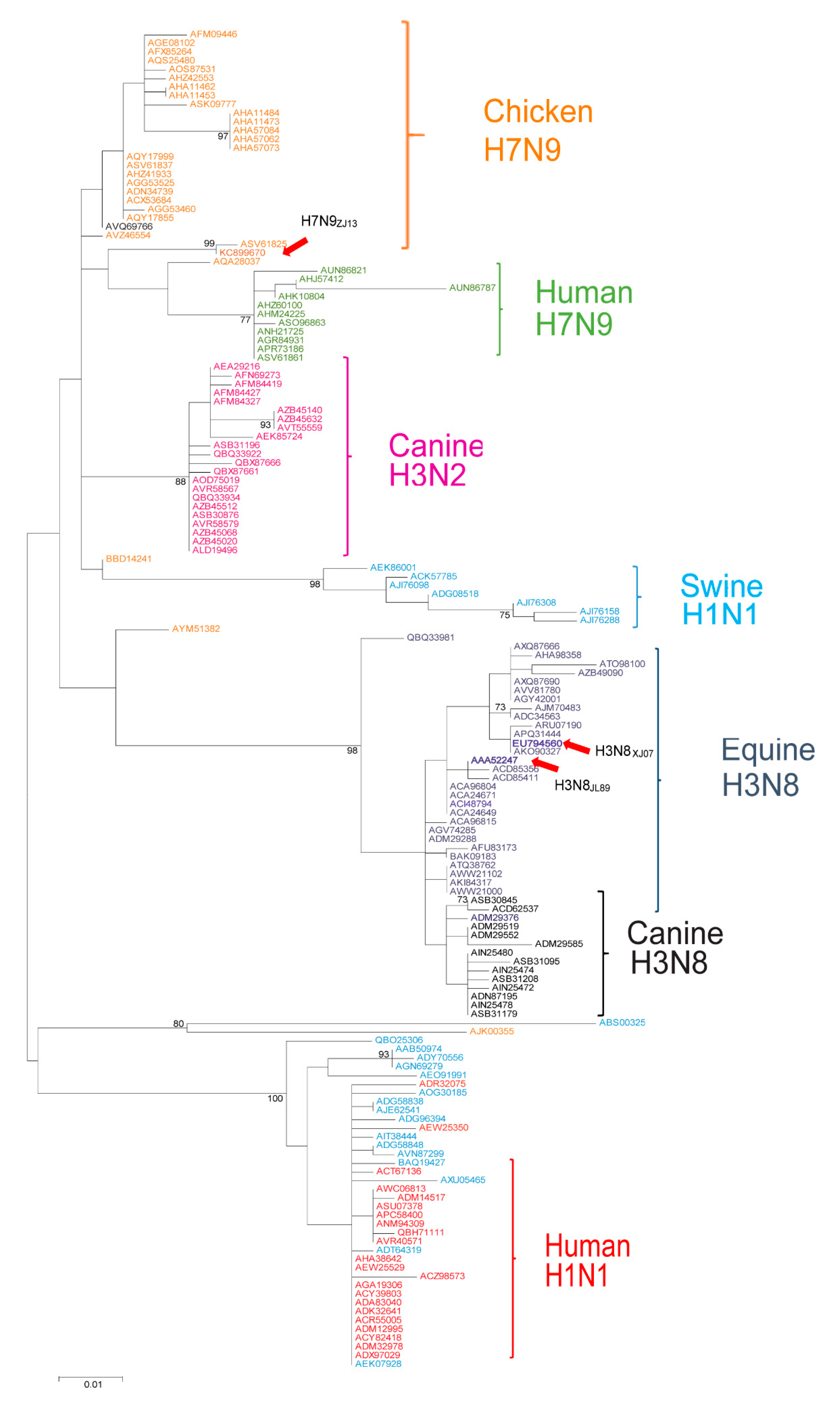
2.8. Evolutionary Analysis
2.9. Statistical Analysis
3. Results
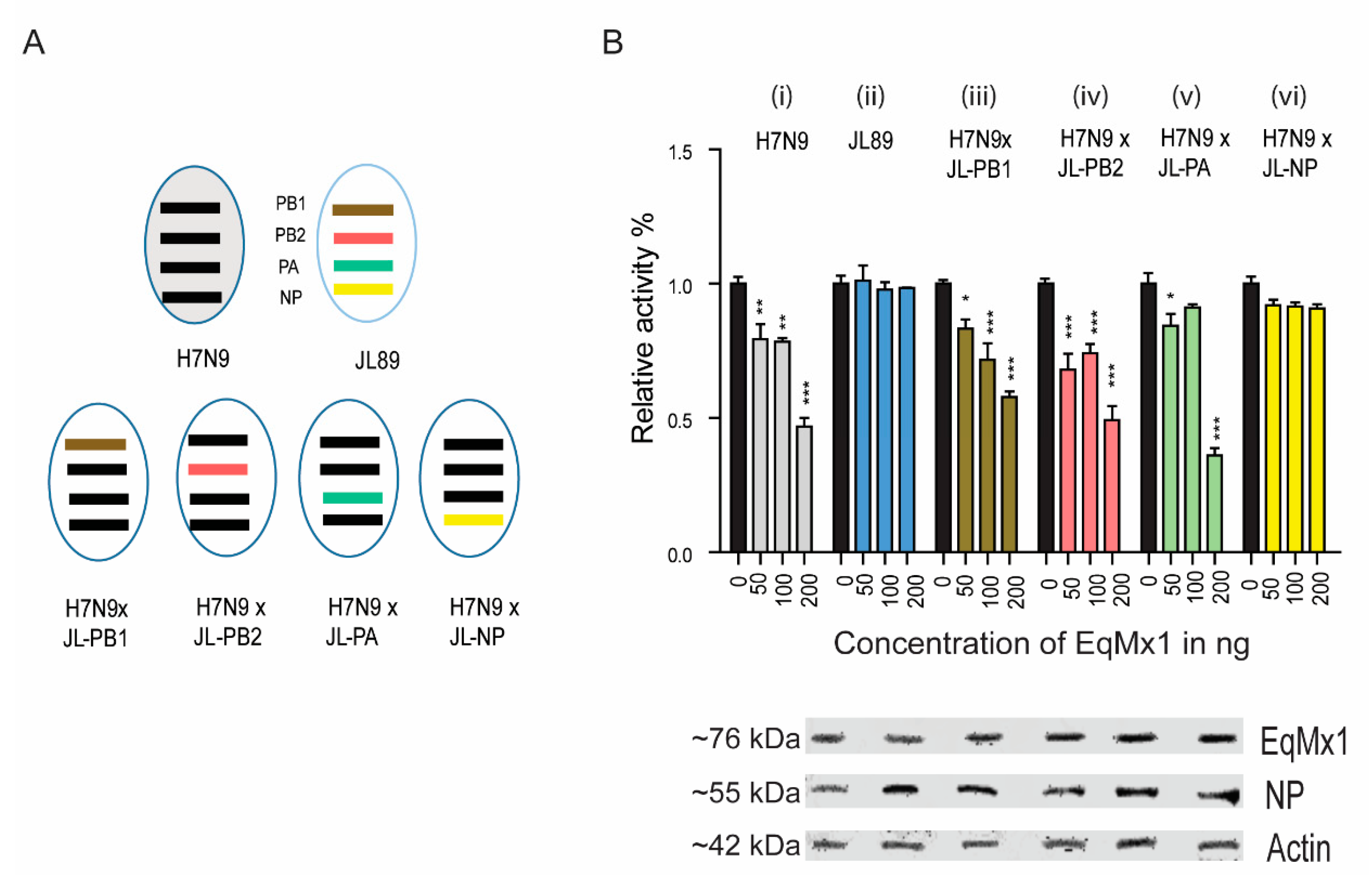
3.1. Identification of Cross-Species Antiviral Potential of Human and Equine Mx1
3.2. The Viral Nucleoprotein as a Possible Target of Mx1 Action
3.3. Identification of Residues in NP of the H3N8JL89 Influenza A Virus Responsible for Resistance to eqMx1
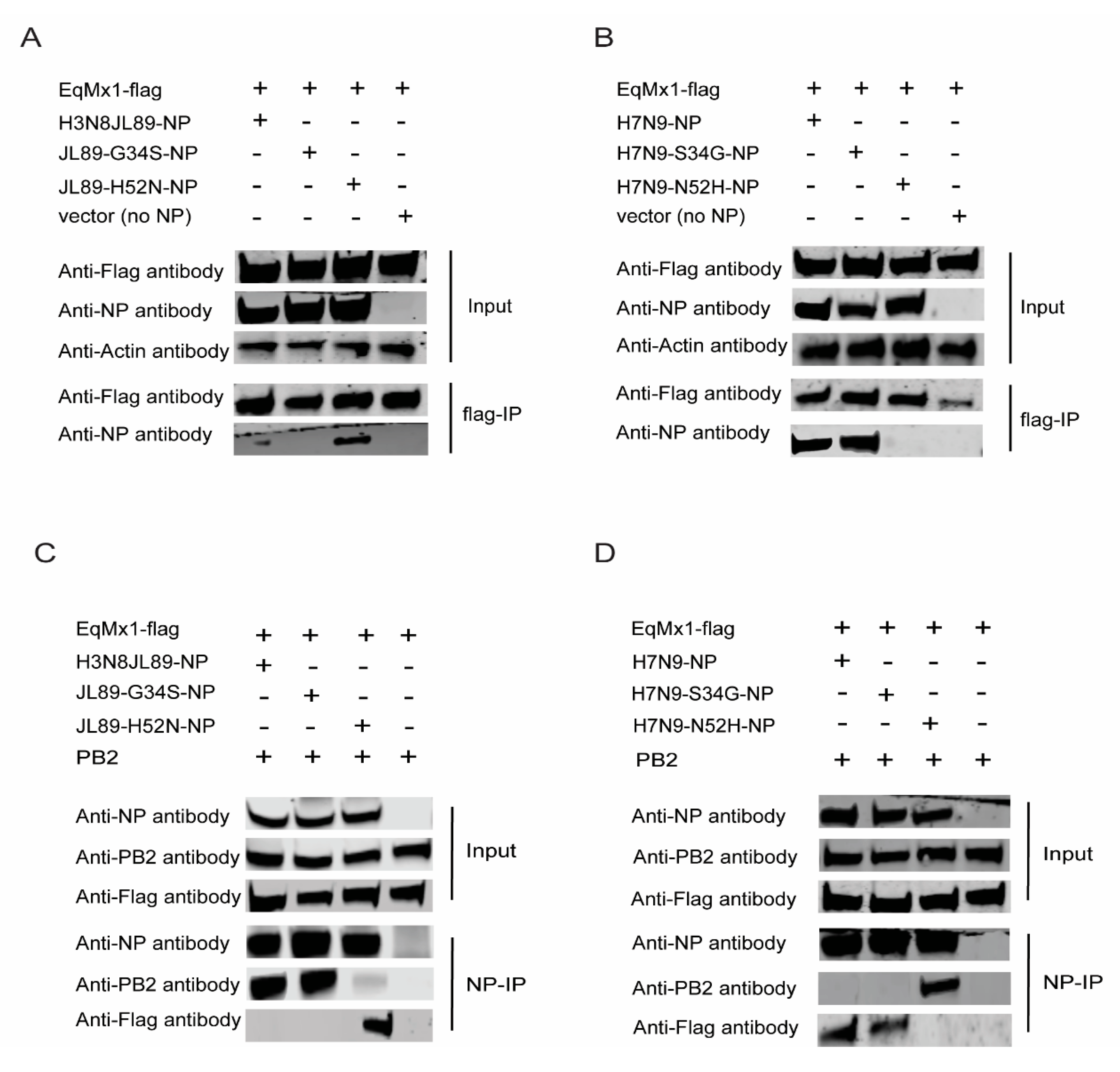
3.4. The Amino Acid Position 52 of NP Affects its Interaction with eqMx1
3.5. A Single Mutation at Position 52 in H3N8JL89 NP Allows the Virus to escape from eqMx1 Restriction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taubenberger, J.K.; Kash, J.C. Influenza virus evolution, host adaptation, and pandemic formation. Cell Host Microbe 2010, 7, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, T.; Kawaoka, Y. Pandemic threat posed by avian influenza A viruses. Clin. Microbiol. Rev. 2001, 14, 129–149. [Google Scholar] [CrossRef]
- Short, K.R.; Richard, M.; Verhagen, J.H.; van Riel, D.; Schrauwen, E.J.; van den Brand, J.M.; Mänz, B.; Bodewes, R.; Herfst, S. One health, multiple challenges: The inter-species transmission of influenza A virus. One Health 2015, 1, 1–13. [Google Scholar] [CrossRef]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Mol. Biol. Rev. 1992, 56, 152–179. [Google Scholar]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic potential of influenza A viruses: A comprehensive overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef]
- Verhelst, J.; Parthoens, E.; Schepens, B.; Fiers, W.; Saelens, X. Interferon-inducible protein Mx1 inhibits influenza virus by interfering with functional viral ribonucleoprotein complex assembly. J. Virol. 2012, 86, 13445–13455. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef]
- Verhelst, J.; Hulpiau, P.; Saelens, X. Mx proteins: Antiviral gatekeepers that restrain the uninvited. Microbiol. Mol. Biol. Rev. 2013, 77, 551–566. [Google Scholar] [CrossRef]
- Xiao, H.; Killip, M.J.; Staeheli, P.; Randall, R.E.; Jackson, D. The human interferon-induced MxA protein inhibits early stages of influenza A virus infection by retaining the incoming viral genome in the cytoplasm. J. Virol. 2013, 87, 13053–13058. [Google Scholar] [CrossRef]
- Pavlovic, J.; Haller, O.; Staeheli, P. Human and mouse Mx proteins inhibit different steps of the influenza virus multiplication cycle. J. Virol. 1992, 66, 2564–2569. [Google Scholar]
- Schneider-Schaulies, S.; Schneider-Schaulies, J.; Schuster, A.; Bayer, M.; Pavlovic, J.; Ter Meulen, V. Cell type-specific MxA-mediated inhibition of measles virus transcription in human brain cells. J. Virol. 1994, 68, 6910–6917. [Google Scholar]
- Kochs, G.; Janzen, C.; Hohenberg, H.; Haller, O. Antivirally active MxA protein sequesters La Crosse virus nucleocapsid protein into perinuclear complexes. Proc. Natl. Acad. Sci. USA 2002, 99, 3153–3158. [Google Scholar] [CrossRef]
- Frese, M.; Kochs, G.; Feldmann, H.; Hertkorn, C.; Haller, O. Inhibition of bunyaviruses, phleboviruses, and hantaviruses by human MxA protein. J. Virol. 1996, 70, 915–923. [Google Scholar]
- Dam Van, P.; Desmecht, D.; Garigliany, M.-M.; Bui Tran Anh, D.; Van Laere, A.-S. Anti-Influenza A Virus Activities of Type I/III Interferons-Induced Mx1 GTPases from Different Mammalian Species. J. Interferon Cytokine Res. 2019, 39, 274–282. [Google Scholar] [CrossRef]
- Gao, S.; von der Malsburg, A.; Dick, A.; Faelber, K.; Schröder, G.F.; Haller, O.; Kochs, G.; Daumke, O. Structure of myxovirus resistance protein a reveals intra-and intermolecular domain interactions required for the antiviral function. Immunity 2011, 35, 514–525. [Google Scholar] [CrossRef]
- Gao, S.; von der Malsburg, A.; Paeschke, S.; Behlke, J.; Haller, O.; Kochs, G.; Daumke, O. Structural basis of oligomerization in the stalk region of dynamin-like MxA. Nature 2010, 465, 502. [Google Scholar] [CrossRef]
- Daumke, O.; Gao, S.; von der Malsburg, A.; Haller, O.; Kochs, G. Structure of the MxA stalk elucidates the assembly of ring-like units of an antiviral module. Small GTPases 2010, 1, 502–506. [Google Scholar] [CrossRef]
- Stranden, A.M.; Staeheli, P.; Pavlovic, J. Function of the mouse Mx1 protein is inhibited by overexpression of the PB2 protein of influenza virus. Virology 1993, 197, 642–651. [Google Scholar] [CrossRef]
- Huang, T.; Pavlovic, J.; Staeheli, P.; Krystal, M. Overexpression of the influenza virus polymerase can titrate out inhibition by the murine Mx1 protein. J. Virol. 1992, 66, 4154–4160. [Google Scholar]
- Mänz, B.; Dornfeld, D.; Götz, V.; Zell, R.; Zimmermann, P.; Haller, O.; Kochs, G.; Schwemmle, M. Pandemic influenza A viruses escape from restriction by human MxA through adaptive mutations in the nucleoprotein. PLoS Pathog. 2013, 9, e1003279. [Google Scholar] [CrossRef]
- Dittmann, J.; Stertz, S.; Grimm, D.; Steel, J.; García-Sastre, A.; Haller, O.; Kochs, G. Influenza A virus strains differ in sensitivity to the antiviral action of Mx-GTPase. J. Virol. 2008, 82, 3624–3631. [Google Scholar] [CrossRef] [PubMed]
- Dornfeld, D.; Petric, P.P.; Hassan, E.; Zell, R.; Schwemmle, M. Eurasian Avian-Like Swine Influenza A Viruses Escape Human MxA Restriction through Distinct Mutations in Their Nucleoprotein. J. Virol. 2019, 93, e00997-18. [Google Scholar] [CrossRef] [PubMed]
- Asano, A.; Ko, J.H.; Morozumi, T.; Hamashima, N.; Watanabe, T. Polymorphisms and the antiviral property of porcine Mx1 protein. J. Vet. Med. Sci. 2002, 64, 1085–1089. [Google Scholar] [CrossRef]
- Babiker, H.; Nakatsu, Y.; Yamada, K.; Yoneda, A.; Takada, A.; Ueda, J.; Hata, H.; Watanabe, T. Bovine and water buffalo Mx2 genes: Polymorphism and antiviral activity. Immunogenetics 2007, 59, 59–67. [Google Scholar] [CrossRef]
- Baise, E.; Pire, G.; Leroy, M.; Gérardin, J.; Goris, N.; Clercq, K.D.; Kerkhofs, P.; Desmecht, D. Conditional expression of type I interferon-induced bovine Mx1 GTPase in a stable transgenic vero cell line interferes with replication of vesicular stomatitis virus. J. Interferon Cytokine Res. 2004, 24, 513–521. [Google Scholar] [CrossRef]
- Yamada, K.; Nakatsu, Y.; Onogi, A.; Ueda, J.; Watanabe, T. Specific intracellular localization and antiviral property of genetic and splicing variants in bovine Mx1. Viral Immunol. 2009, 22, 389–395. [Google Scholar] [CrossRef]
- Riegger, D.; Hai, R.; Dornfeld, D.; Mänz, B.; Leyva-Grado, V.; Sánchez-Aparicio, M.T.; Albrecht, R.A.; Palese, P.; Haller, O.; Schwemmle, M. The nucleoprotein of newly emerged H7N9 influenza A virus harbors a unique motif conferring resistance to antiviral human MxA. J. Virol. 2015, 89, 2241–2252. [Google Scholar] [CrossRef]
- Götz, V.; Magar, L.; Dornfeld, D.; Giese, S.; Pohlmann, A.; Höper, D.; Kong, B.-W.; Jans, D.A.; Beer, M.; Haller, O. Influenza A viruses escape from MxA restriction at the expense of efficient nuclear vRNP import. Sci. Rep. 2016, 6, 23138. [Google Scholar] [CrossRef]
- Deeg, C.M.; Hassan, E.; Mutz, P.; Rheinemann, L.; Götz, V.; Magar, L.; Schilling, M.; Kallfass, C.; Nürnberger, C.; Soubies, S. In vivo evasion of MxA by avian influenza viruses requires human signature in the viral nucleoprotein. J. Exp. Med. 2017, 214, 1239–1248. [Google Scholar] [CrossRef]
- Mino, S.; Mojsiejczuk, L.; Guo, W.; Zhang, H.; Qi, T.; Du, C.; Zhang, X.; Wang, J.; Campos, R.; Wang, X. Equine Influenza Virus in Asia: Phylogeographic Pattern and Molecular Features Reveal Circulation of an Autochthonous Lineage. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Sovinova, O.; Tumova, B.; Pouska, F.; Nemec, J. Isolation of a virus causing respiratory disease in horses. Acta Virol. 1958, 2, 52. [Google Scholar] [PubMed]
- Gorman, O.; Bean, W.; Kawaoka, Y.; Donatelli, I.; Guo, Y.; Webster, R. Evolution of influenza A virus nucleoprotein genes: Implications for the origins of H1N1 human and classical swine viruses. J. Virol. 1991, 65, 3704–3714. [Google Scholar] [PubMed]
- Lin, Y.-Z.; Cao, X.-Z.; Li, L.; Li, L.; Jiang, C.-G.; Wang, X.-F.; Ma, J.; Zhou, J.-H. The pathogenic and vaccine strains of equine infectious anemia virus differentially induce cytokine and chemokine expression and apoptosis in macrophages. Virus Res. 2011, 160, 274–282. [Google Scholar] [CrossRef]
- Zimmermann, P.; Mänz, B.; Haller, O.; Schwemmle, M.; Kochs, G. The viral nucleoprotein determines Mx sensitivity of influenza A viruses. J. Virol. 2011, 85, 8133–8140. [Google Scholar] [CrossRef] [PubMed]
- Pleschka, S.; Jaskunas, R.; Engelhardt, O.G.; Zürcher, T.; Palese, P.; Garcia-Sastre, A. A plasmid-based reverse genetics system for influenza A virus. J. Virol. 1996, 70, 4188–4192. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Z.; Wang, Y.; Wang, M.; Wang, X.; Zhang, X.; Ji, S.; Du, C.; Chen, H.; Wang, X. Fundamental contribution and host range determination of ANP32 protein family in influenza A virus polymerase activity. bioRxiv 2019, 529412. [Google Scholar]
- Verhelst, J.; De Vlieger, D.; Saelens, X. Co-immunoprecipitation of the mouse Mx1 protein with the influenza a virus nucleoprotein. JoVE (J. Vis. Exp.) 2015, e52871. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, Z.; Wang, X. Strain-specific antagonism of the human H1N1 influenza A virus against equine tetherin. Viruses 2018, 10, 264. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Ashenberg, O.; Padmakumar, J.; Doud, M.B.; Bloom, J.D. Deep mutational scanning identifies sites in influenza nucleoprotein that affect viral inhibition by MxA. PLoS Pathog. 2017, 13, e1006288. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.K.; Boutz, P.L.; Nayak, D.P. Influenza virus nucleoprotein interacts with influenza virus polymerase proteins. J. Virol. 1998, 72, 5493–5501. [Google Scholar]
- Lear, T.; Breen, M.; Ponce de Leon, F.; Coogle, L.; Ferguson, E.; Chambers, T.; Bailey, E. Cloning and chromosomal localization of MX1 and ETS2 to chromosome 26 of the horse (Equus caballus). Chromosome Res. 1998, 6, 333–334. [Google Scholar] [CrossRef]
- Heinz, H.; Marquardt, J.; Schuberth, H.-J.; Adolf, G.; Leibold, W. Proteins induced by recombinant equine interferon-β1 within equine peripheral blood mononuclear cells and polymorphonuclear neutrophilic granulocytes. Vet. Immunol. Immunopathol. 1994, 42, 221–235. [Google Scholar] [CrossRef]
- Taubenberger, J.; Morens, D. Pandemic influenza–including a risk assessment of H5N1. Rev. Sci. Tech. (Int. Off. Epizoot.) 2009, 28, 187. [Google Scholar] [CrossRef] [Green Version]
- Crawford, P.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J. Transmission of equine influenza virus to dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.; Zhou, H.; Jiang, T.; Li, C.; Zhang, A.; Guo, X.; Zou, W.; Chen, H.; Jin, M. Isolation and molecular characterization of equine H3N8 influenza viruses from pigs in China. Arch. Virol. 2009, 154, 887–890. [Google Scholar] [CrossRef]
- Engelhardt, O.G.; Ullrich, E.; Kochs, G.; Haller, O. Interferon-induced antiviral Mx1 GTPase is associated with components of the SUMO-1 system and promyelocytic leukemia protein nuclear bodies. Exp. Cell Res. 2001, 271, 286–295. [Google Scholar] [CrossRef]
- Patzina, C.; Haller, O.; Kochs, G. Structural requirements for the antiviral activity of the human MxA protein against Thogoto and influenza A virus. J. Biol. Chem. 2014, 289, 6020–6027. [Google Scholar] [CrossRef] [Green Version]
- Giese, S.; Ciminski, K.; Bolte, H.; Moreira, É.A.; Lakdawala, S.; Hu, Z.; David, Q.; Kolesnikova, L.; Götz, V.; Zhao, Y. Role of influenza A virus NP acetylation on viral growth and replication. Nat. Commun. 2017, 8, 1259. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatima, U.; Zhang, Z.; Zhang, H.; Wang, X.-F.; Xu, L.; Chu, X.; Ji, S.; Wang, X. Equine Mx1 Restricts Influenza A Virus Replication by Targeting at Distinct Site of its Nucleoprotein. Viruses 2019, 11, 1114. https://doi.org/10.3390/v11121114
Fatima U, Zhang Z, Zhang H, Wang X-F, Xu L, Chu X, Ji S, Wang X. Equine Mx1 Restricts Influenza A Virus Replication by Targeting at Distinct Site of its Nucleoprotein. Viruses. 2019; 11(12):1114. https://doi.org/10.3390/v11121114
Chicago/Turabian StyleFatima, Urooj, Zhenyu Zhang, Haili Zhang, Xue-Feng Wang, Ling Xu, Xiaoyu Chu, Shuang Ji, and Xiaojun Wang. 2019. "Equine Mx1 Restricts Influenza A Virus Replication by Targeting at Distinct Site of its Nucleoprotein" Viruses 11, no. 12: 1114. https://doi.org/10.3390/v11121114
APA StyleFatima, U., Zhang, Z., Zhang, H., Wang, X. -F., Xu, L., Chu, X., Ji, S., & Wang, X. (2019). Equine Mx1 Restricts Influenza A Virus Replication by Targeting at Distinct Site of its Nucleoprotein. Viruses, 11(12), 1114. https://doi.org/10.3390/v11121114