Chrysoviruses Inhabited Symbiotic Fungi of Lichens


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
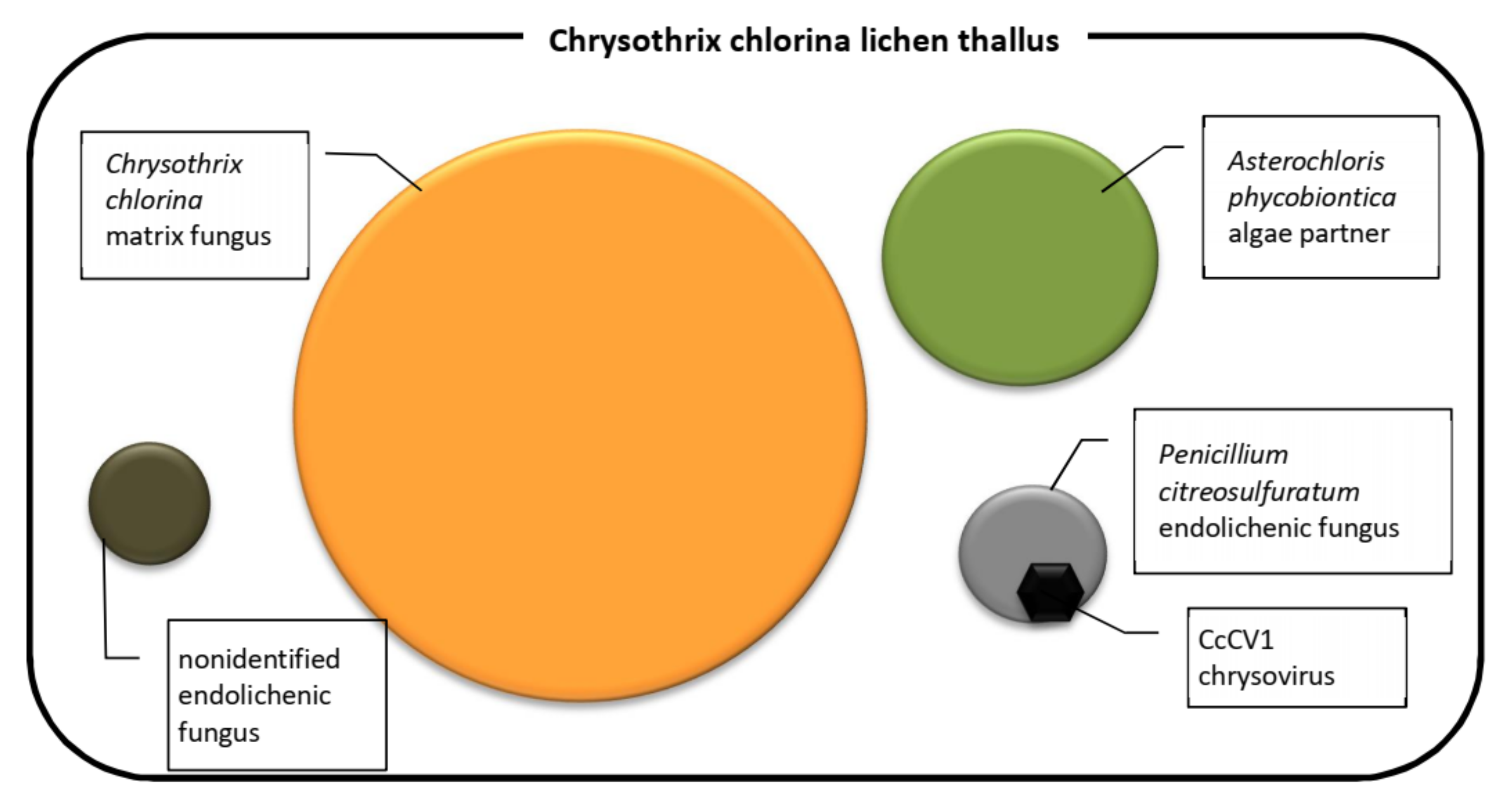
2.1. Lichen Samples
2.2. Double-Stranded RNA Extraction and Virus Sequencing
2.3. Lichen Symbionts Separation
2.4. In situ Hybridization
3. Results
3.1. Lichen Identification
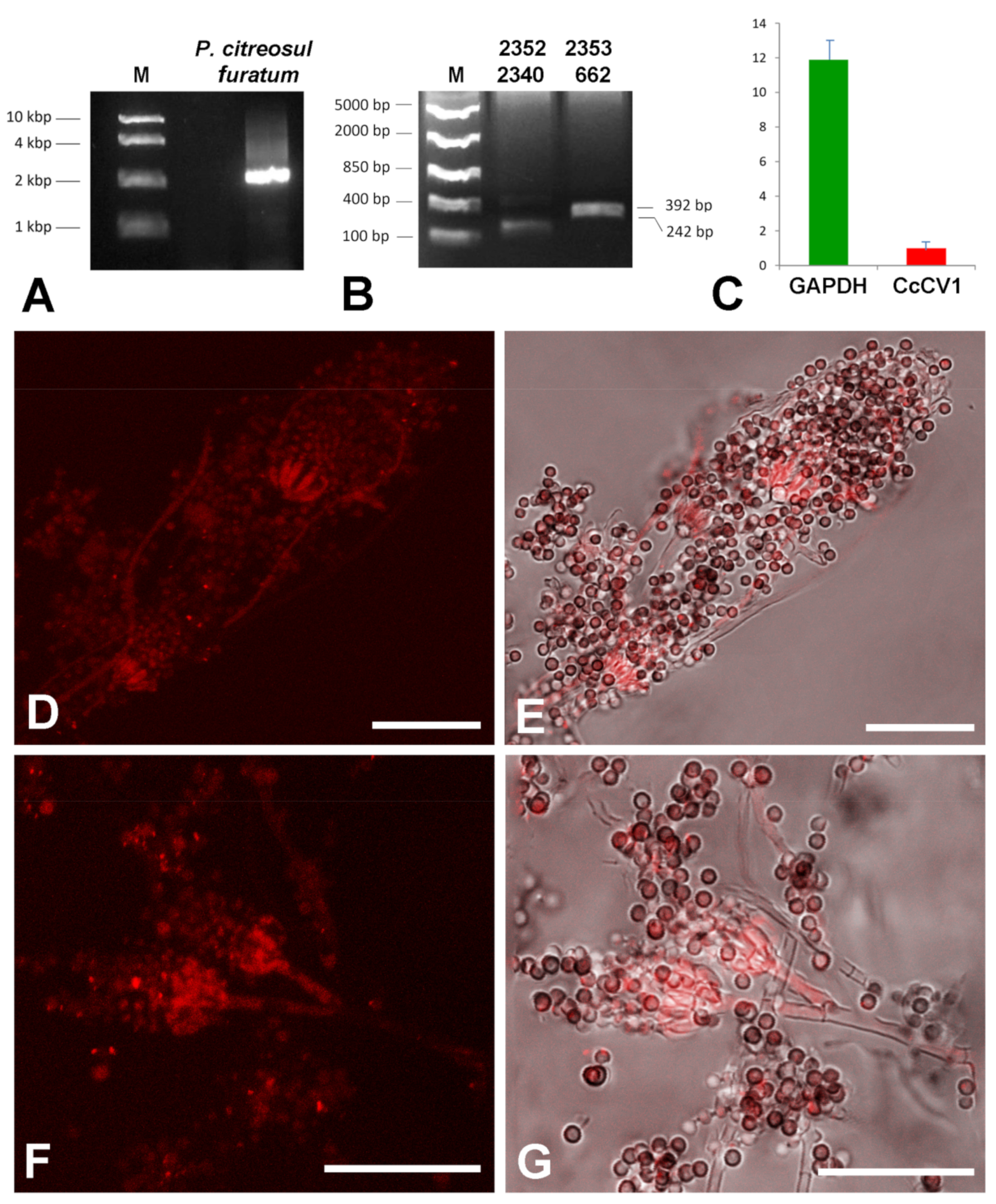
3.2. dsRNA Presence
3.3. Concentration of the Virus in Lichen
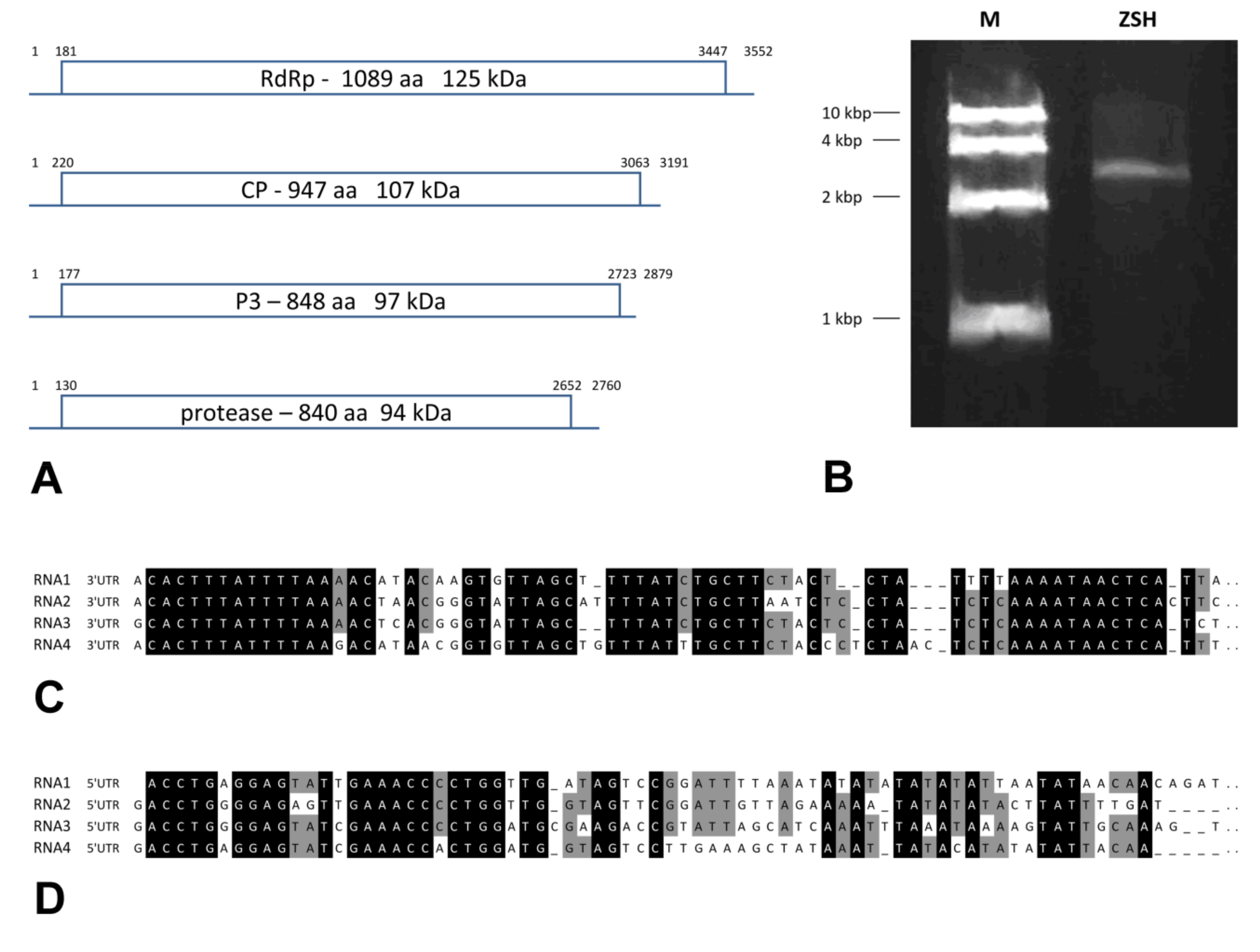
3.4. Genome Description of the Viruses
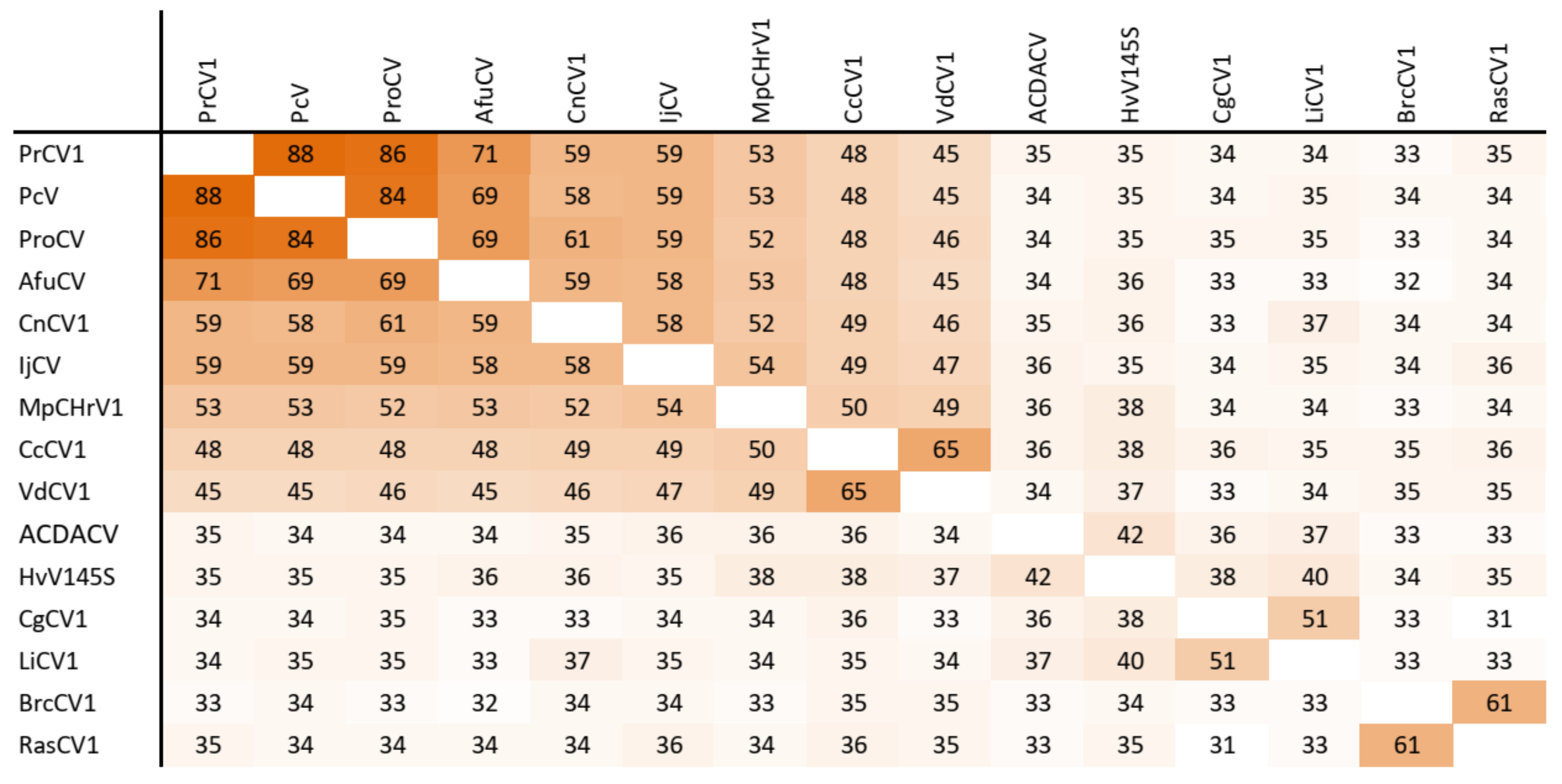
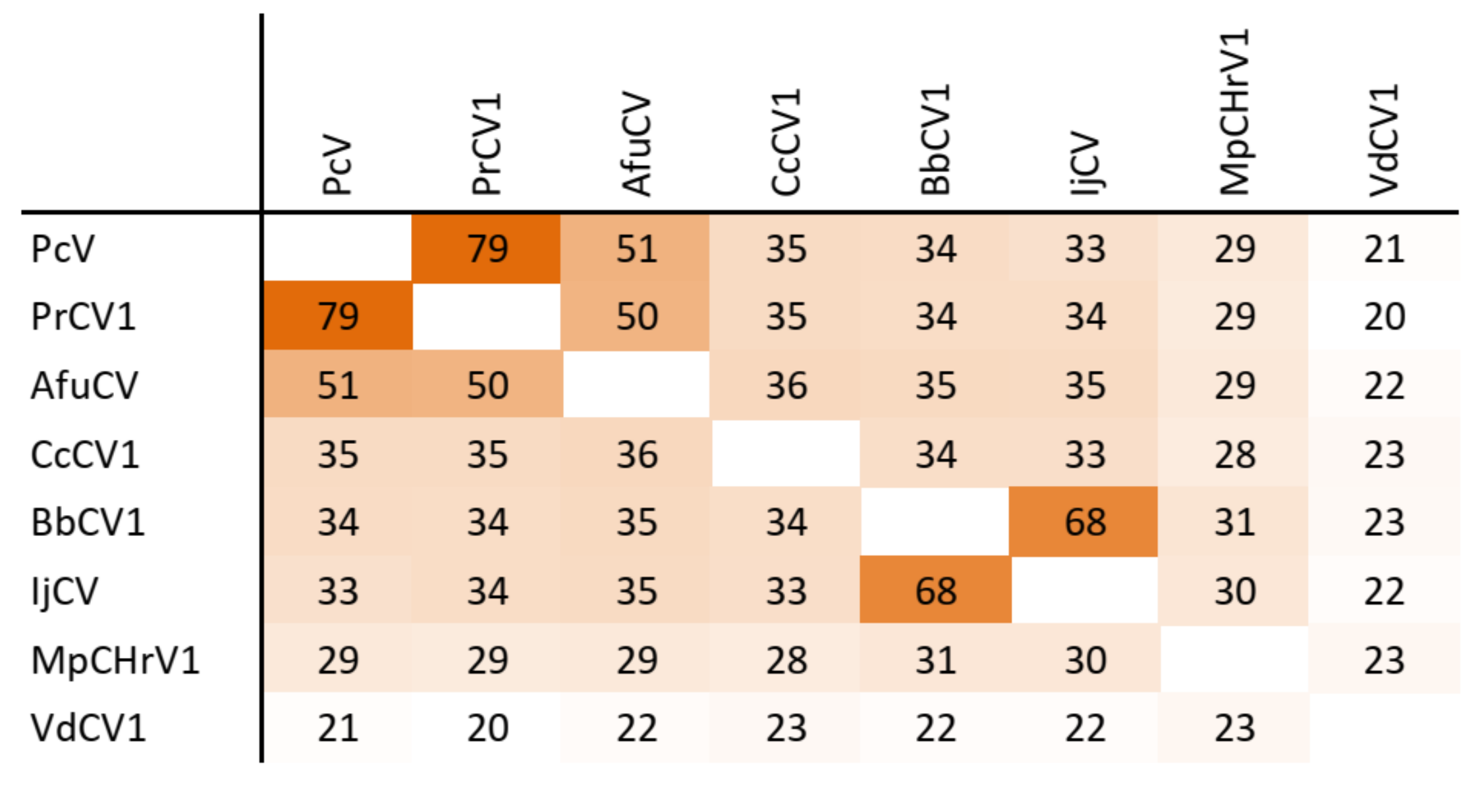
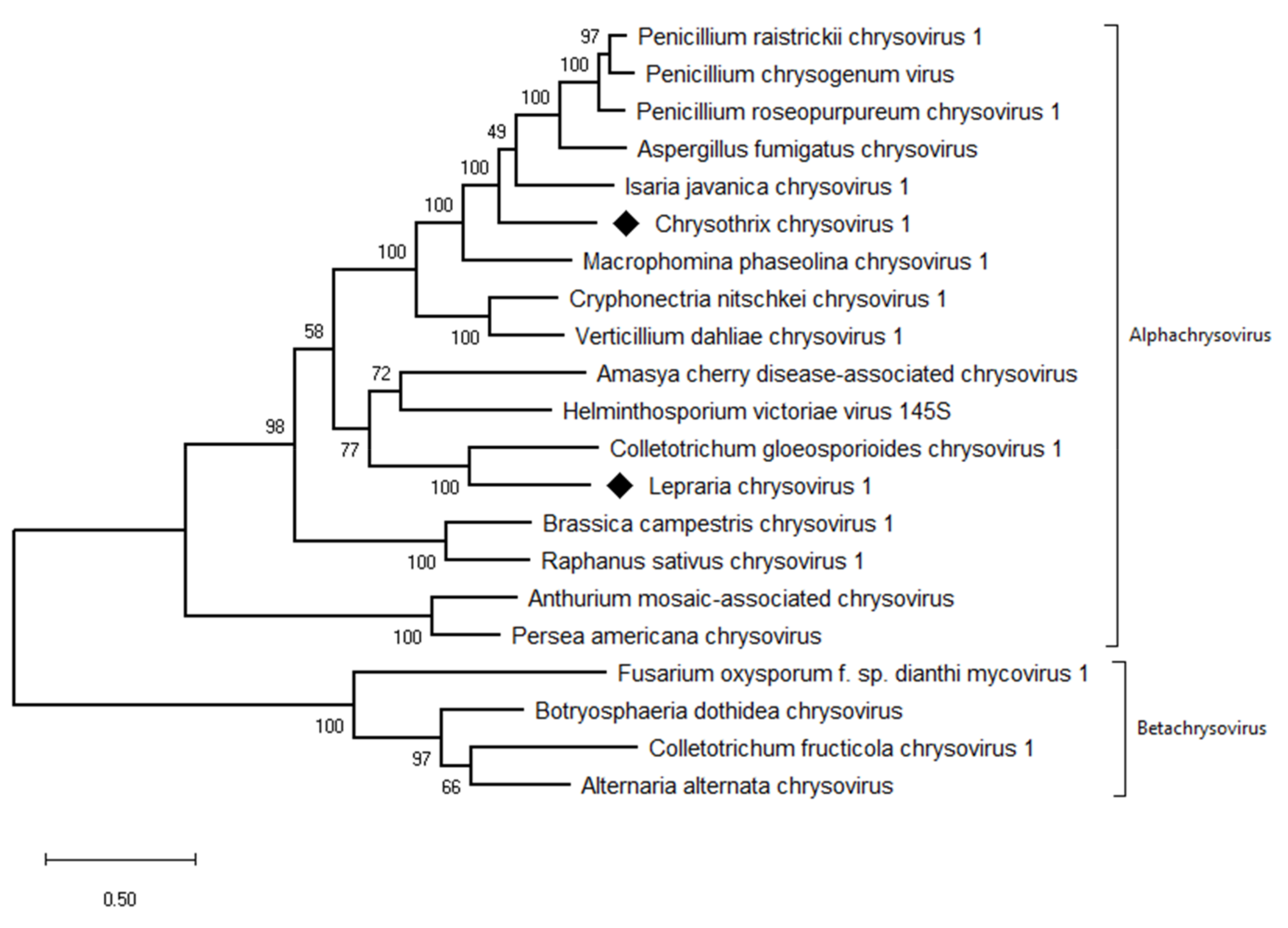
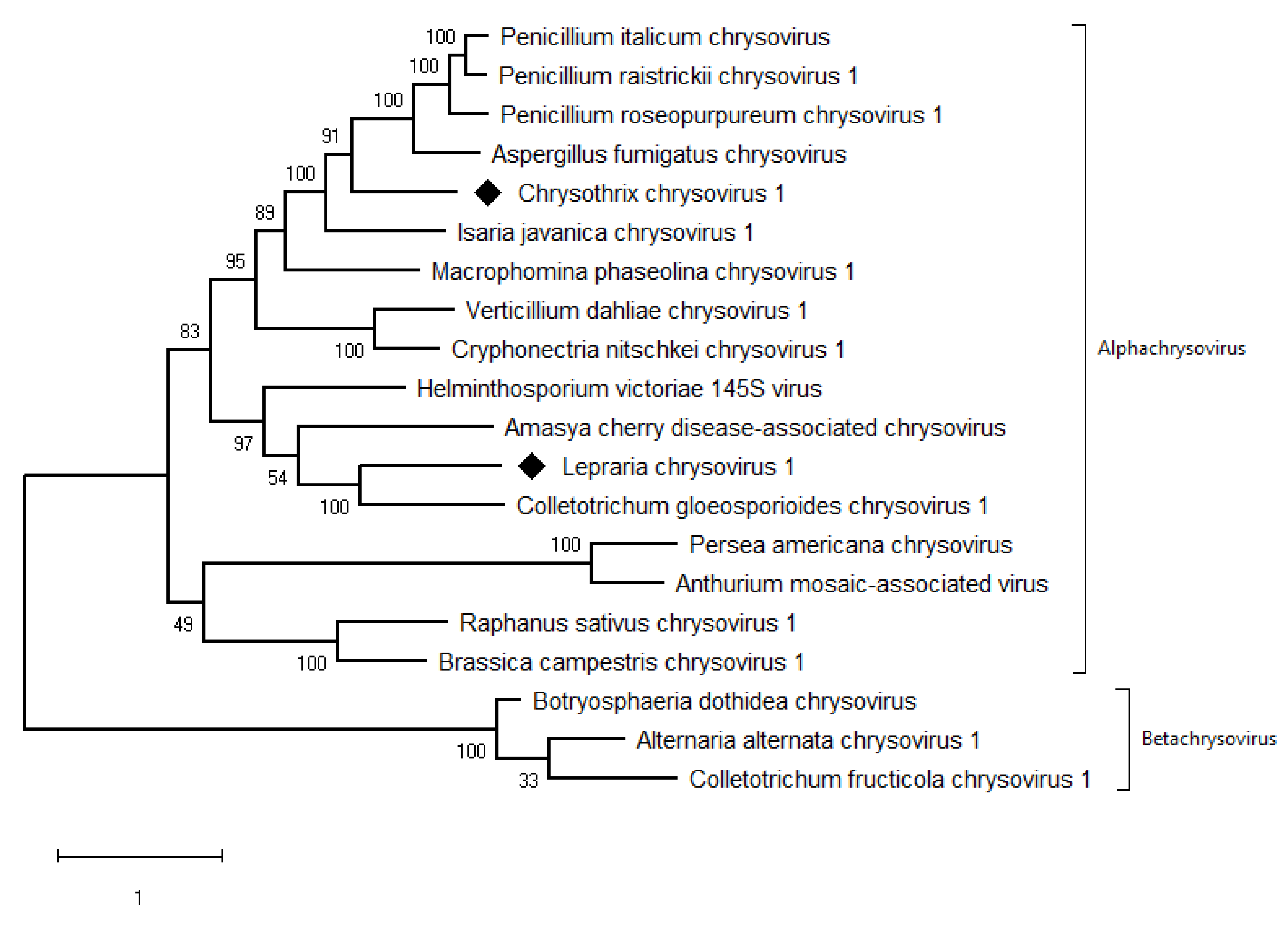
3.5. Taxonomic Relationship
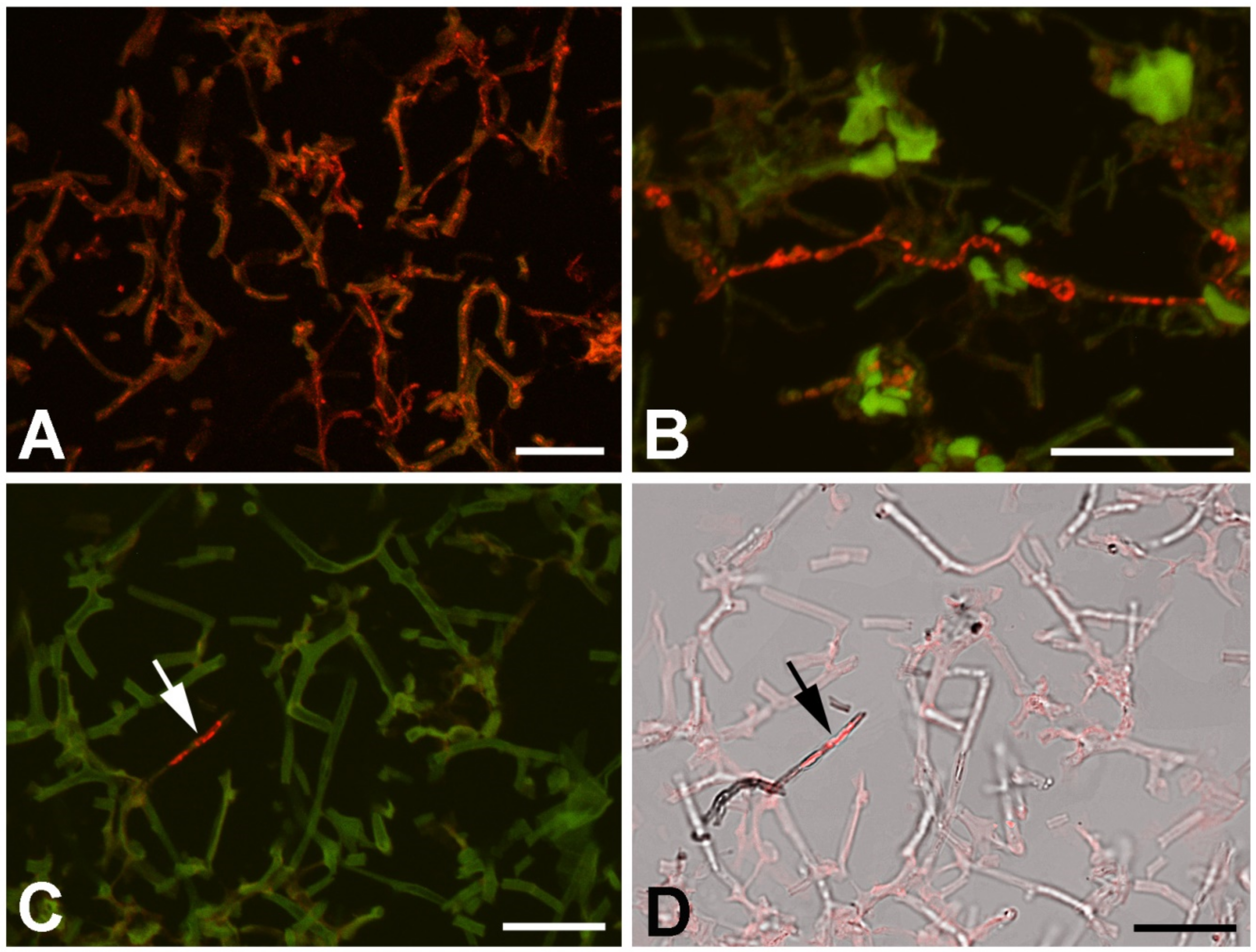
3.6. Virus Localization in Lichen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blair, J.E. Fungi. In The Timetree of life; Hedges, S.B., Kumar, S., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 215–219. [Google Scholar]
- Lawrey, J.D.; Diederich, P. Lichenicolous fungi: Interactions, evolution, and biodiversity. Bryol. 2003, 106, 80–120. [Google Scholar] [CrossRef]
- Hodkinson, B.P.; Lutzoni, F. A microbiotic survey of lichen-associated bacteria reveals a new lineage from the Rhizobiales. Symbiosis 2009, 49, 163–180. [Google Scholar] [CrossRef]
- Grube, M.; Berg, G. Microbial consortia of bacteria and fungi with focus on the lichen symbiosis. Fungal Biol. Rev. 2009, 23, 72–85. [Google Scholar] [CrossRef]
- Kellog, J.J.; Raja, H.A. Endolichenic fungi: A new source of rich bioactive secondary metabolites on the horizon. Phytochem. Rev. 2017, 16, 271–293. [Google Scholar] [CrossRef] [Green Version]
- Lawrey, J.D. Chemical interactions between two lichen-degrading fungi. J. Chem. Ecol. 2000, 26, 1821–1831. [Google Scholar] [CrossRef]
- Lawrey, J.D.; Torzilli, A.P.; Chandhoeke, V. Destruction of lichen chemical defenses by fungal pathogen. Am. J. Bot. 1999, 86, 187–189. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The variety of fungal-algal symbioses, their evolutionary significance, and the nature of lichens. Bot. J. Linn. Soc. 1988, 96, 3–20. [Google Scholar] [CrossRef]
- Armstrong, R.A.; Welch, A.R. Competition in lichen communities. Symbiosis 2007, 43, 1–12. [Google Scholar]
- Richardson, D.H.S. War in the world of lichens: Parasitism and symbiosis as exemplified by lichens and lichenicolous fungi. Mycol. Res. 1999, 103, 641–650. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Coy, S.R.; Gann, E.R.; Pound, H.L.; Short, S.M.; Wilhelm, S.W. Viruses of eukaryotic algae: Diversity, methods for detection, and future directions. Viruses 2018, 10, 487. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Li, T.; Deng, F.; Hu, Z. Freshwater cyanophages. Virol. Sin. 2013, 28, 253–259. [Google Scholar] [CrossRef]
- Petrzik, K.; Vondrák, J.; Barták, M.; Peksa, O.; Kubešová, O. Lichens—New source or yet unknown host of herbaceous plant viruses? Eur. J. Plant. Pathol. 2014, 138, 549–559. [Google Scholar] [CrossRef]
- Martin, R.R.; Zhou, J.; Tzanetakis, I.E. Blueberry latent virus: An amalgam of the Partitiviridae and Totiviridae. Virus Res. 2011, 155, 175–180. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Ghanem-Sabanadzovic, N.A. Molecular characterization and detection of a tripartite cryptic virus from rose. J. Plant. Pathol. 2008, 90, 287–293. [Google Scholar]
- Yolken, R.H.; Jones-Brando, L.; Dunigan, D.D.; Kannan, G.; Dickerson, F.; Severance, E.; Sabunciyan, S.; Talbot, C.C.; Prandovszky, E.; Gurnon, J.R.; et al. Chlorovirus ATCV-1 is part of the human oropharyngeal virome and is associated with changes in cognitive functions in humans and mice. Proc. Nat. Acad. Sci. USA 2014, 111, 16106–16111. [Google Scholar] [CrossRef] [Green Version]
- Ekman, S.; Tønsberg, T. Most species of Lepraria and Leproloma form a monophyletic group closely related to Stereocaulon. Mycol. Res. 2002, 106, 1262–1276. [Google Scholar] [CrossRef]
- Antane, S.; Caufield, C.E.; Hu, W.; Keeney, D.; Labthavikul, P.; Morris, K.; Naughton, S.M.; Petersen, P.J.; Rasmussen, B.A.; Singh, G.; et al. Pulvinones as bacterial cell wall biosynthesis inhibitors. Bioorg. Med. Chem. Lett. 2006, 16, 176–180. [Google Scholar] [CrossRef]
- Sahin, E.; Psav, S.D.; Avan, I.; Candan, M.; Sahinturk, V.; Koparal, A.T. Vulpinic acid, a lichen metabolite emerges as a potential drug candidate in the therapy of oxidative stress-related diseases, such as atherosclerosis. Human Exp. Toxicol. 2019, 38, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Saag, L.; Saag, A.; Randlane, T. World survey of the genus Lepraria (Stereocaulaceae, lichenized Ascomycota). Lichenol. 2009, 41, 25–60. [Google Scholar] [CrossRef]
- Lubicz, J.V.; Rush, C.M.; Payton, M.; Colberg, T. Beet necrotic yellow vein virus accumulates inside resting spores and zoosporangia of its vector Polymyxa betae BNYVV infects P. betae. Virol. J. 2007, 4, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, T.J.; Dodds, J.A. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef] [Green Version]
- Darissa, O.; Willingmann, P.; Adam, G. Optimized approaches for the sequence determination of double-stranded RNA templates. J. Virol. Methods 2010, 169, 397–403. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Gasulla, F.; Guéra, A.; Barreno, E. A simple and rapid method for isolating lichen photobionts. Symbiosis 2010, 51, 175–179. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.X.; Chen, J. Transcriptome analysis identifies candidate genes associated with melanin and toxin biosynthesis and pathogenicity of the maize pathogen, Curvularia lunata. J. Phytopathol. 2018, 166, 233–241. [Google Scholar] [CrossRef]
- Baschien, C.; Manz, W.; Neu, T.R.; Marvanová, L.; Szewzyk, U. In situ detection of freshwater fungi in an alpine stream by new taxon-specific fluorescence in situ hybridization probes. Appl. Environ. Microb. 2008, 74, 6427–6436. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI Bioinformatics Toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- ICTV Taxonomy history: Alphachrysovirus, 2018b. Available online: https://talk.ictvonline.org//taxonomy/p/taxonomy-history?taxnode_id=201852890 (accessed on 4 November 2019).
- Visagie, C.M.; Seifert, K.A.; Houbraken, J.; Samson, R.A.; Jacobs, K. A phylogenetic revision of Penicillium sect. Exilicaulis, including nine new species from fynbos in South Africa. IMA Fungus 2016, 7, 75–117. [Google Scholar] [CrossRef] [Green Version]
- Ghabrial, S.A.; Castón, J.R. Chrysoviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 509–513. [Google Scholar]
- Webster, C.L.; Waldron, F.M.; Robertson, S.; Crowson, D.; Ferrari, G.; Quintana, J.F.; Brouqui, J.M.; Bayne, E.H.; Longdon, B.; Buck, A.H.; et al. The discovery, distribution, and evolution of viruses associated with Drosophila melanogaster. PLoS Biol. 2015, 13, e1002210. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.S.; Imrie, A.; Holmes, E.C. High-Resolution metatranscriptomics reveals the ecological dynamics of mosquito-associated RNA viruses in Western Australia. J. Virol. 2017, 92, e00680-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Discovery of novel dsRNA viral sequences by in silico cloning and implications for viral diversity, host range and evolution. PLoS ONE 2012, 7, e42147. [Google Scholar] [CrossRef] [PubMed]
- Petrzik, K. Evolutionary forces at work in partitiviruses. Virus Genes 2019, 55, 563–573. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Xu, A.; Wang, T.; Chen, J.; Zhu, X. Molecular characterization of a trisegmented chrysovirus isolated from the radish Raphanus sativus. Virus Res. 2016, 176, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Higashiura, T.; Katoh, Y.; Urayama, S.; Hayashi, O.; Aihara, M.; Fukuhara, T.; Fuji, S.; Kobayashi, T.; Hase, S.; Arie, T.; et al. Magnaporthe oryzae chrysovirus 1 strain D confers growth inhibition to the host fungus and exhibit multiform viral structural proteins. Virology 2019, 535, 241–254. [Google Scholar] [CrossRef]
- Ejmal, M.A.; Holland, D.J.; MacDiarmid, R.M.; Pearson, M.N. A novel chrysovirus from a clinical isolate of Aspergillus thermomutatus affects sporulation. PLoS ONE 2018, 13, e0209443. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef]
- Torres-Trenas, A.; Prieto, P.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Mycovirus Fusarium oxysporum f. sp. dianthi virus 1 decreases the colonizing efficiency of its fungal host. Front. Cell. Infect. Mi. 2019, 9, 51. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, J.; Wang, Y.; Hong, N.; Zhang, F.; Xu, W.; Wang, G. Hypovirulence of the phytopathogenic fungus Botryosphaeria dothidea: Association with a coinfecting chrysovirus and a partitivirus. J. Virol. 2014, 88, 7517–7527. [Google Scholar] [CrossRef] [Green Version]
- Urayama, S.; Kato, S.; Suzuki, Y.; Aoki, N.; Le, M.T.; Arie, T.; Teraoka, T.; Fukuhara, T.; Moriyama, H. Mycoviruses related to chrysovirus affect vegetative growth in the rice blast fungus Magnaporthe oryzae. J. Gen. Virol. 2010, 91, 3085–3094. [Google Scholar] [CrossRef] [PubMed]
- Urayama, S.; Sakoda, H.; Takai, R.; Katoh, Y.; Minh, L.T.; Fukuhara, T.; Arie, T.; Teraoka, T.; Moriyama, H. A dsRNA mycovirus, Magnaporthe oryzae chrysovirus 1-B, suppresses vegetative growth and development of the rice blast fungus. Virology 2014, 448, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmsen, M.C.; Tolner, B.; Kram, A.; Go, S.J.; de Haan, A.; Wessels, J.G.H. Sequences of three dsRNAs associated with La France disease of the cultivated mushroom (Agaricus bisporus). Curr. Genet. 1991, 20, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, H.; Urayama, S.; Higashiura, T.; Le, T.M.; Komatsu, K. Chrysoviruses in Magnaporthe oryzae. Viruses 2018, 10, 697. [Google Scholar] [CrossRef] [Green Version]
- Aoki, N.; Moriyama, H.; Kodama, M.; Arie, T.; Teraoka, T.; Fukuhara, T. A novel mycovirus associated with four double-stranded RNAs affects host fungal growth in Alternaria alternata. Virus Res. 2009, 140, 179–187. [Google Scholar] [CrossRef]
- Kim, J.M.; Jung, J.E.; Park, J.A.; Park, S.M.; Cha, B.J.; Kim, D.H. Biological function of a novel chrysovirus CnV1-BS122, in the Korean Cryphonectria nitschkei BS122 strain. J. Biosci. Bioeng. 2012, 115, 1–3. [Google Scholar] [CrossRef]
- Herrero, N. Identification and sequence determination of a new chrysovirus infecting the entomopathogenic fungus Isaria javanica. Arch. Virol. 2017, 162, 1113–1117. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrzik, K.; Koloniuk, I.; Sehadová, H.; Sarkisova, T. Chrysoviruses Inhabited Symbiotic Fungi of Lichens. Viruses 2019, 11, 1120. https://doi.org/10.3390/v11121120
Petrzik K, Koloniuk I, Sehadová H, Sarkisova T. Chrysoviruses Inhabited Symbiotic Fungi of Lichens. Viruses. 2019; 11(12):1120. https://doi.org/10.3390/v11121120
Chicago/Turabian StylePetrzik, Karel, Igor Koloniuk, Hana Sehadová, and Tatiana Sarkisova. 2019. "Chrysoviruses Inhabited Symbiotic Fungi of Lichens" Viruses 11, no. 12: 1120. https://doi.org/10.3390/v11121120
APA StylePetrzik, K., Koloniuk, I., Sehadová, H., & Sarkisova, T. (2019). Chrysoviruses Inhabited Symbiotic Fungi of Lichens. Viruses, 11(12), 1120. https://doi.org/10.3390/v11121120