Genetic Characteristics and Pathogenicity Analysis in Chickens and Mice of Three H9N2 Avian Influenza Viruses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Isolation
2.2. Phylogenetic and Molecular Analysis
2.3. Animal Studies
2.3.1. Chickens
2.3.2. Mice
2.4. Ethics Statements
3. Results
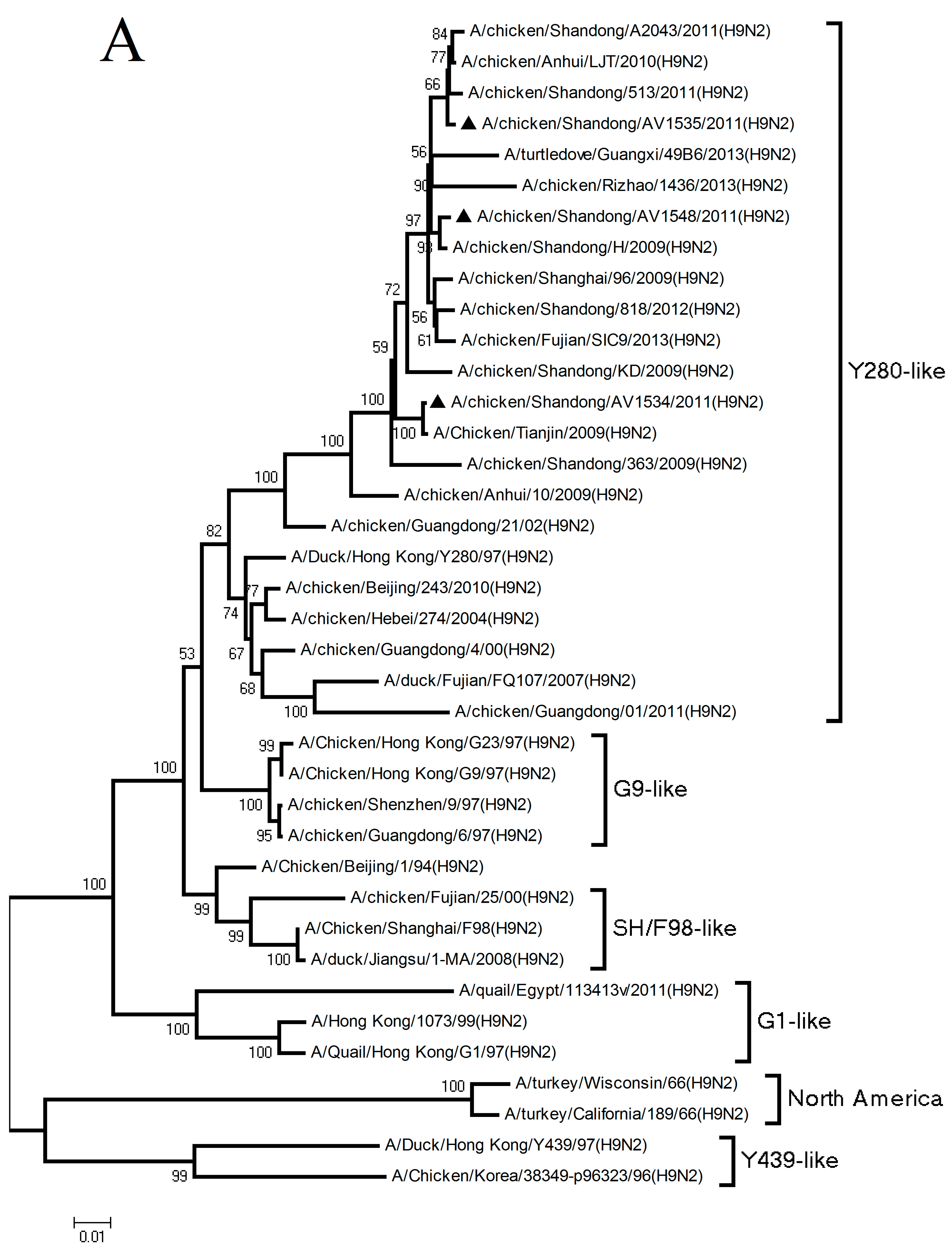
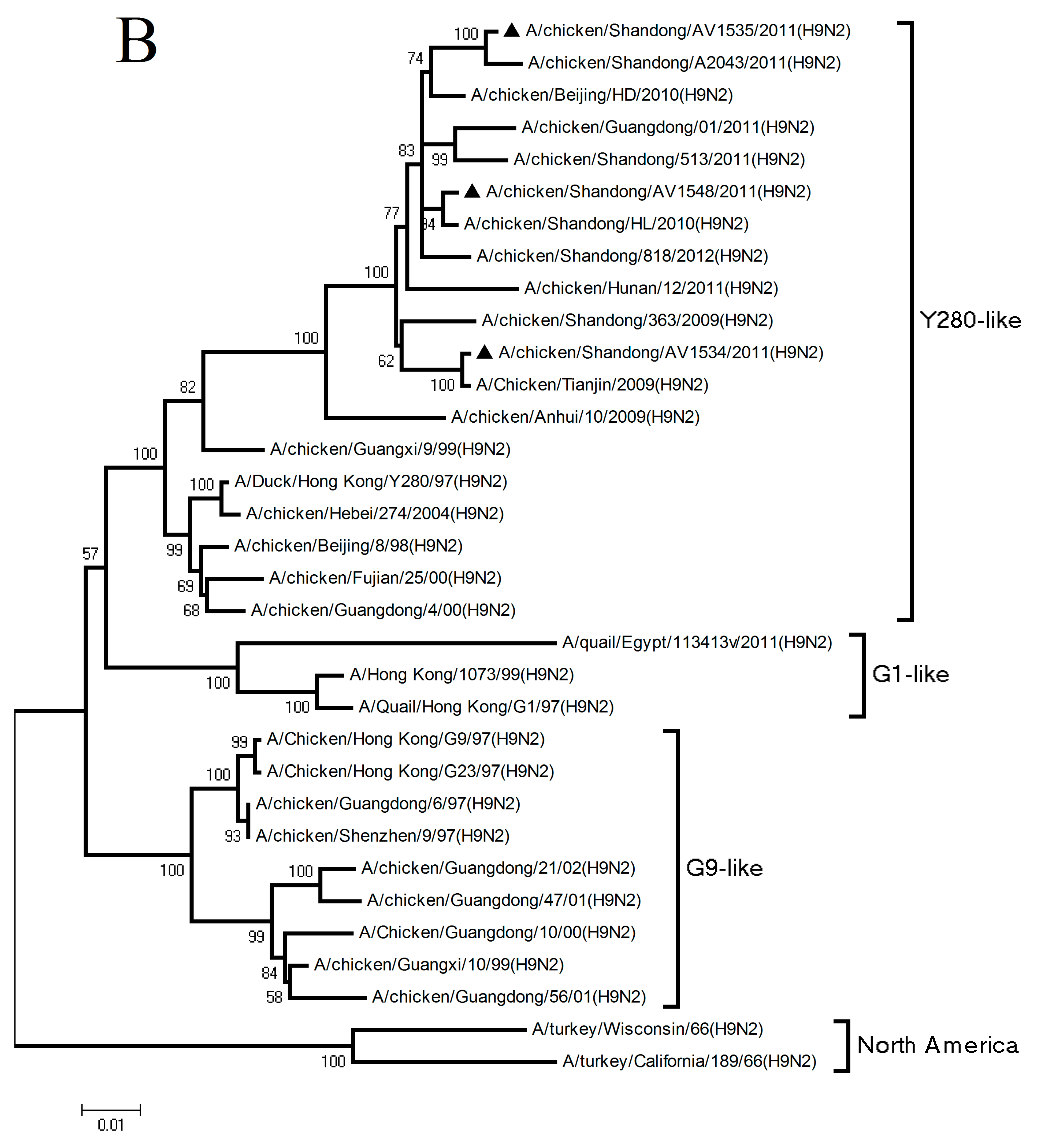
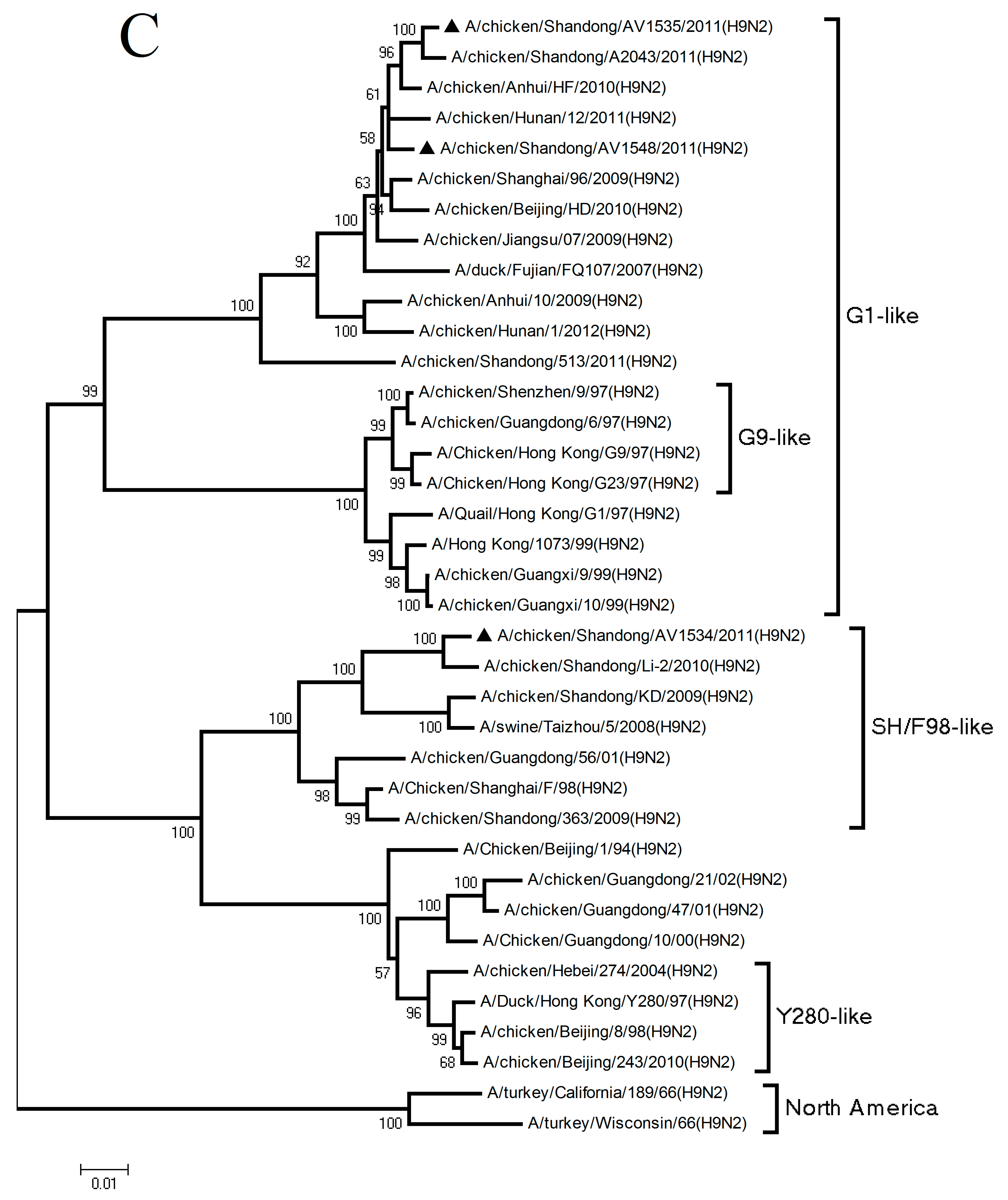
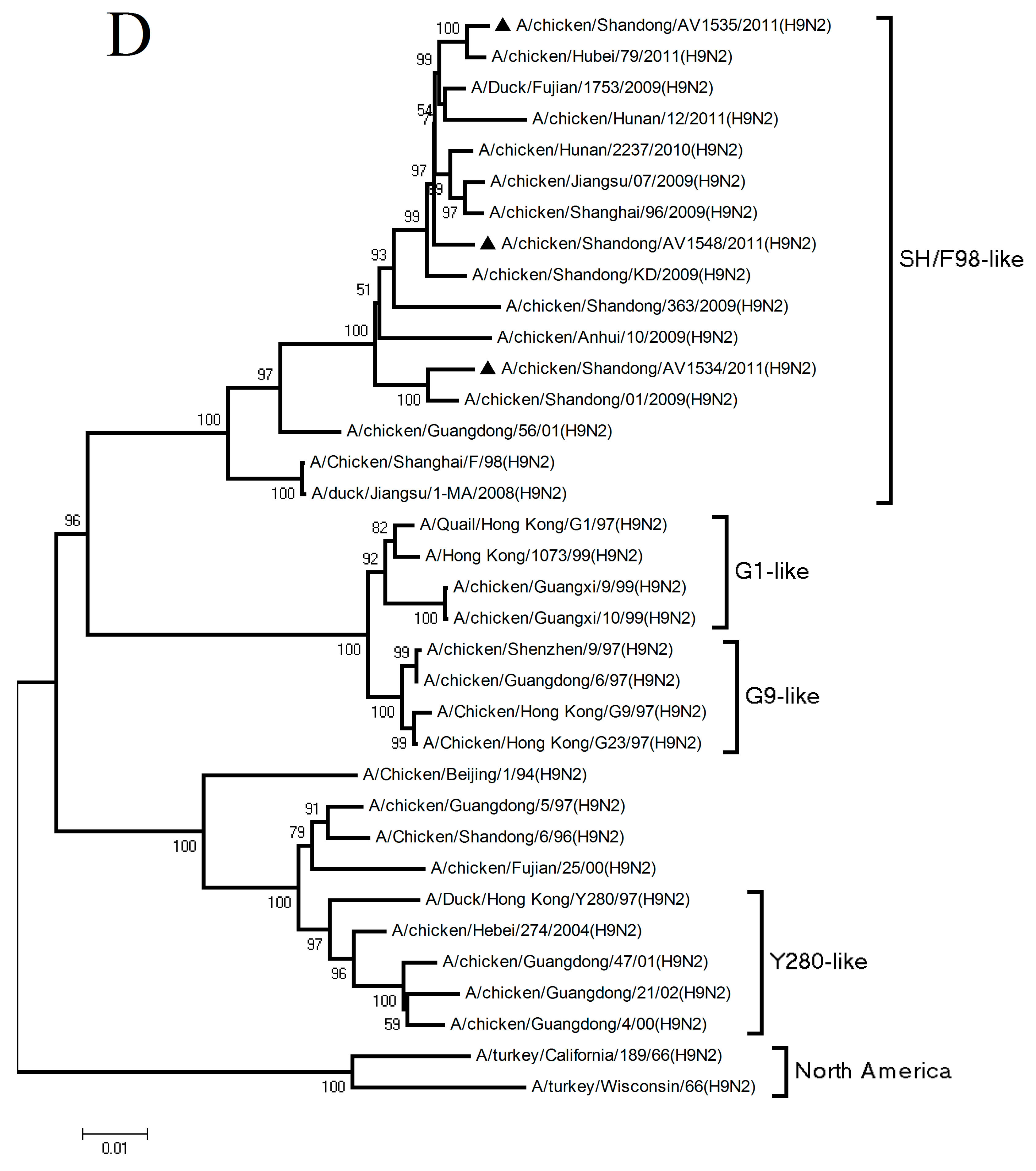
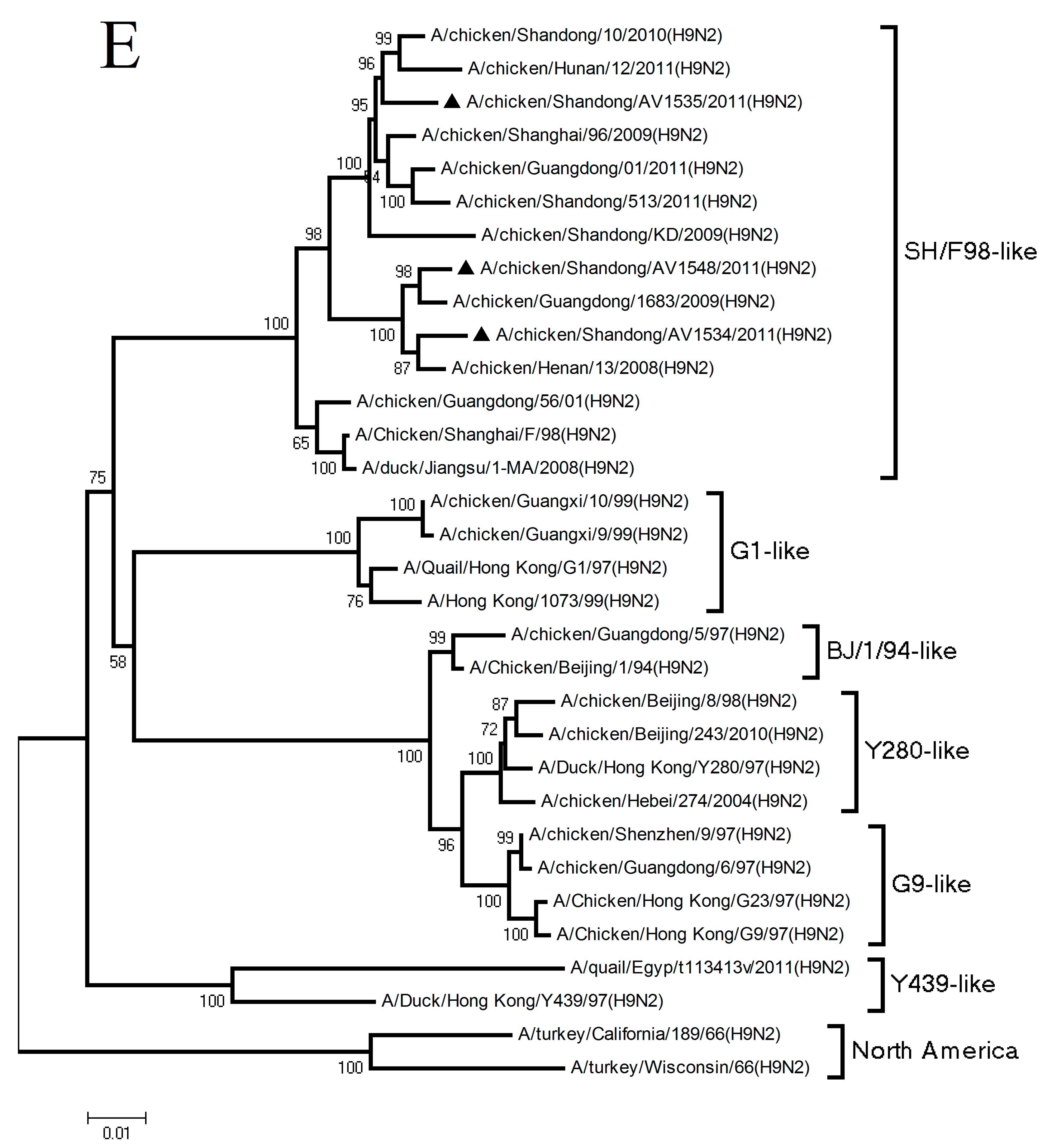
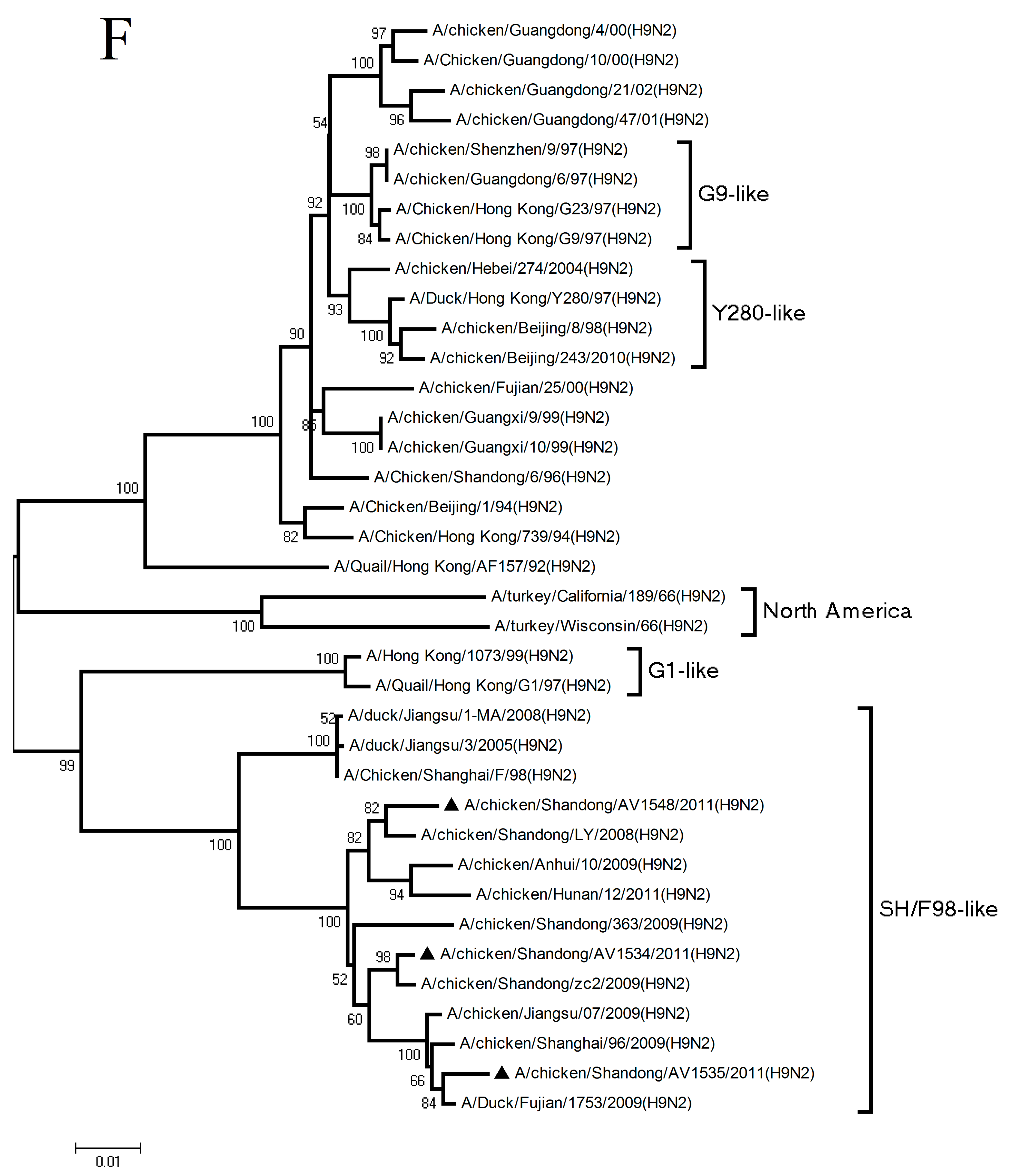
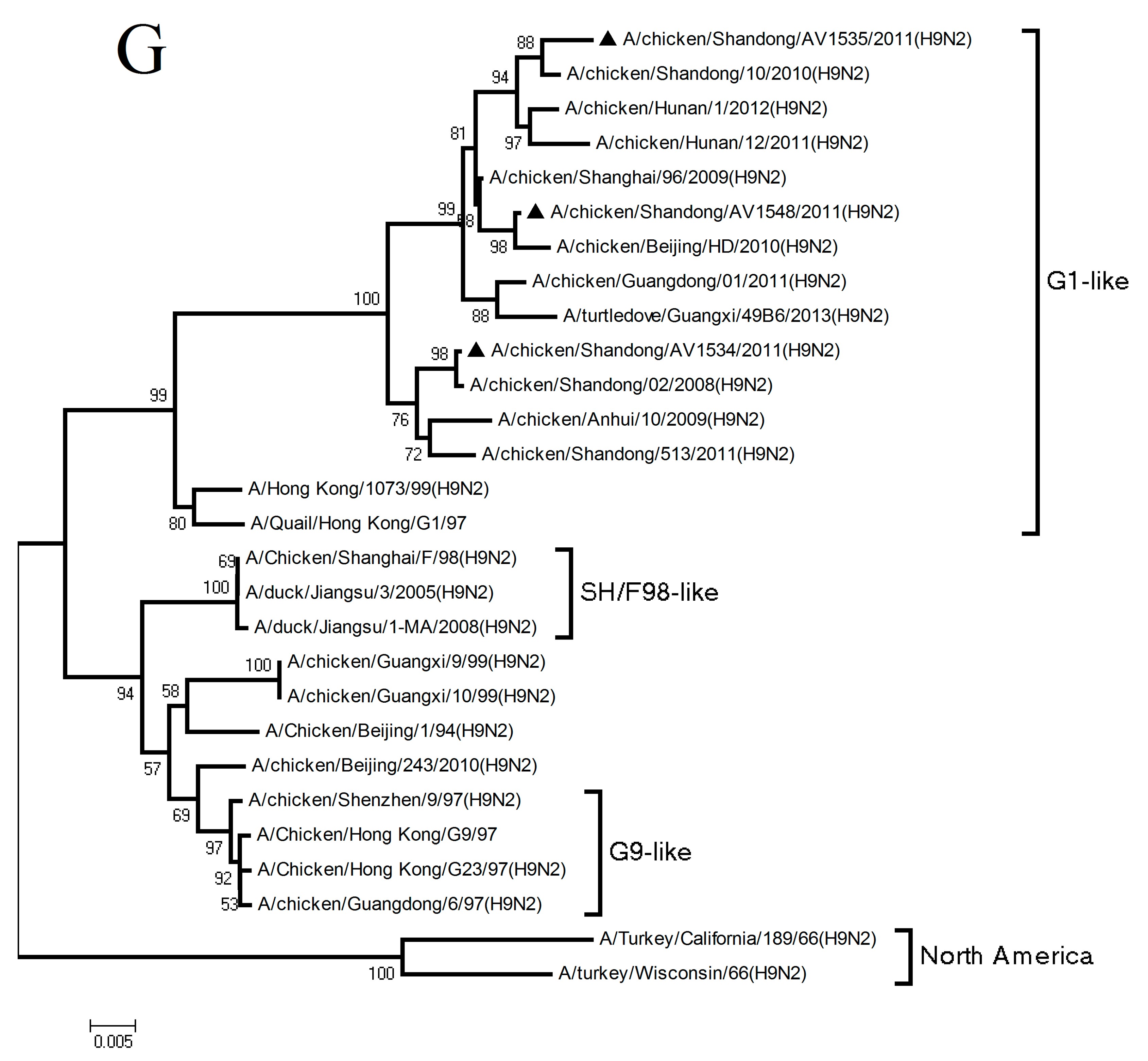
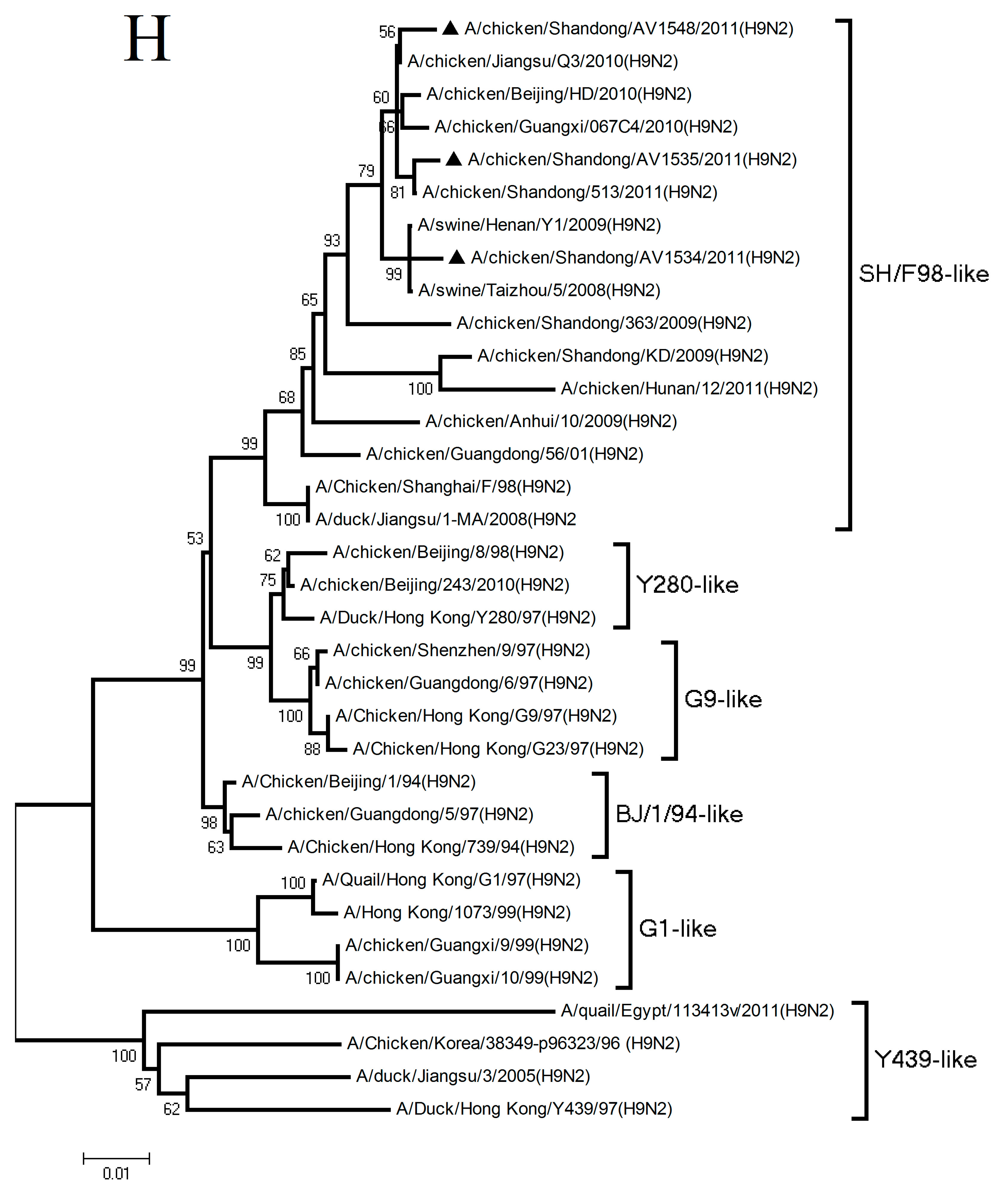
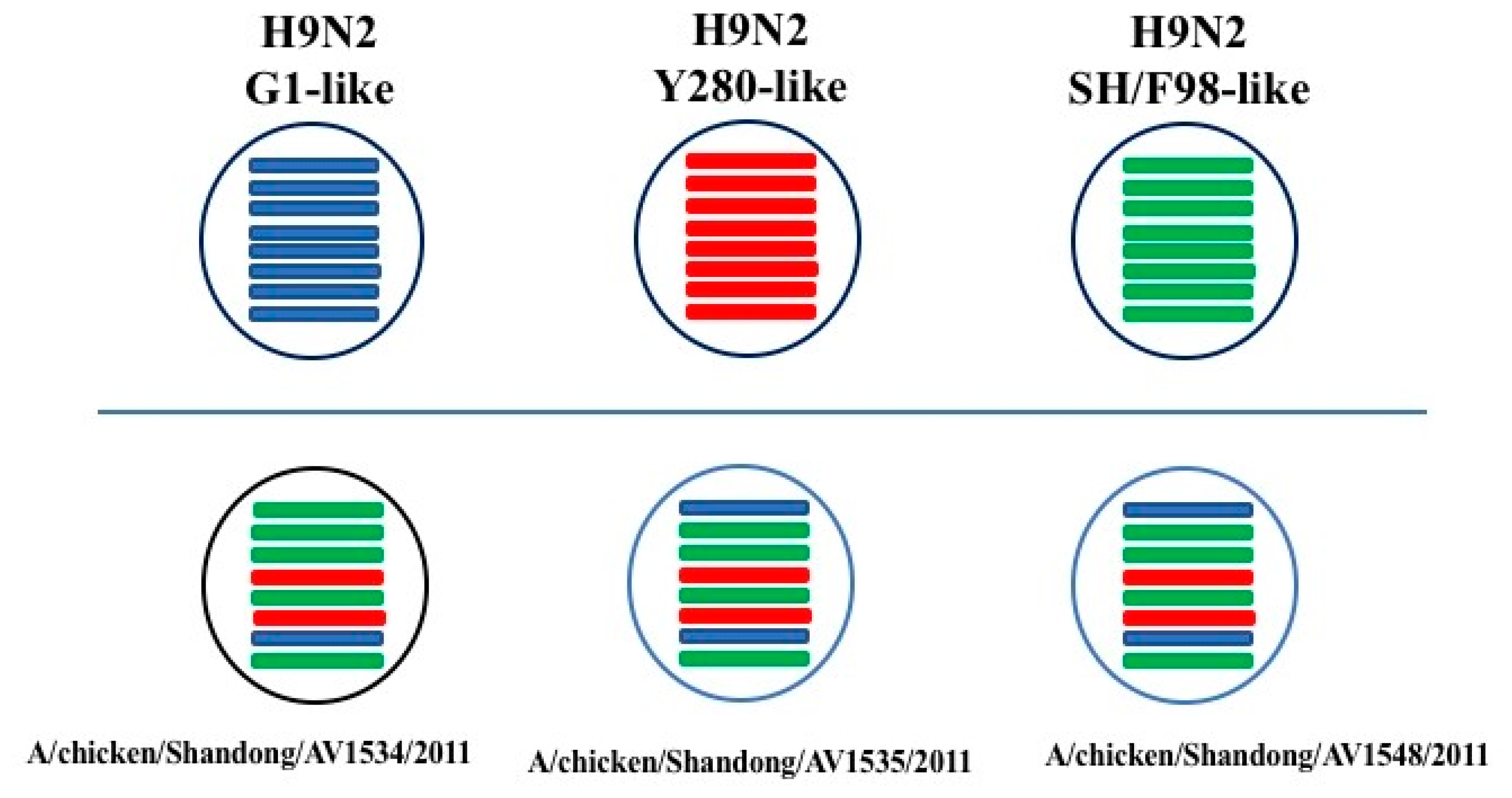
3.1. Phylogenetic Analysis of the Three H9N2 Viruses
3.2. Molecular Characterization
3.3. Pathogenicity and Transmission of These H9N2 Viruses in Chickens
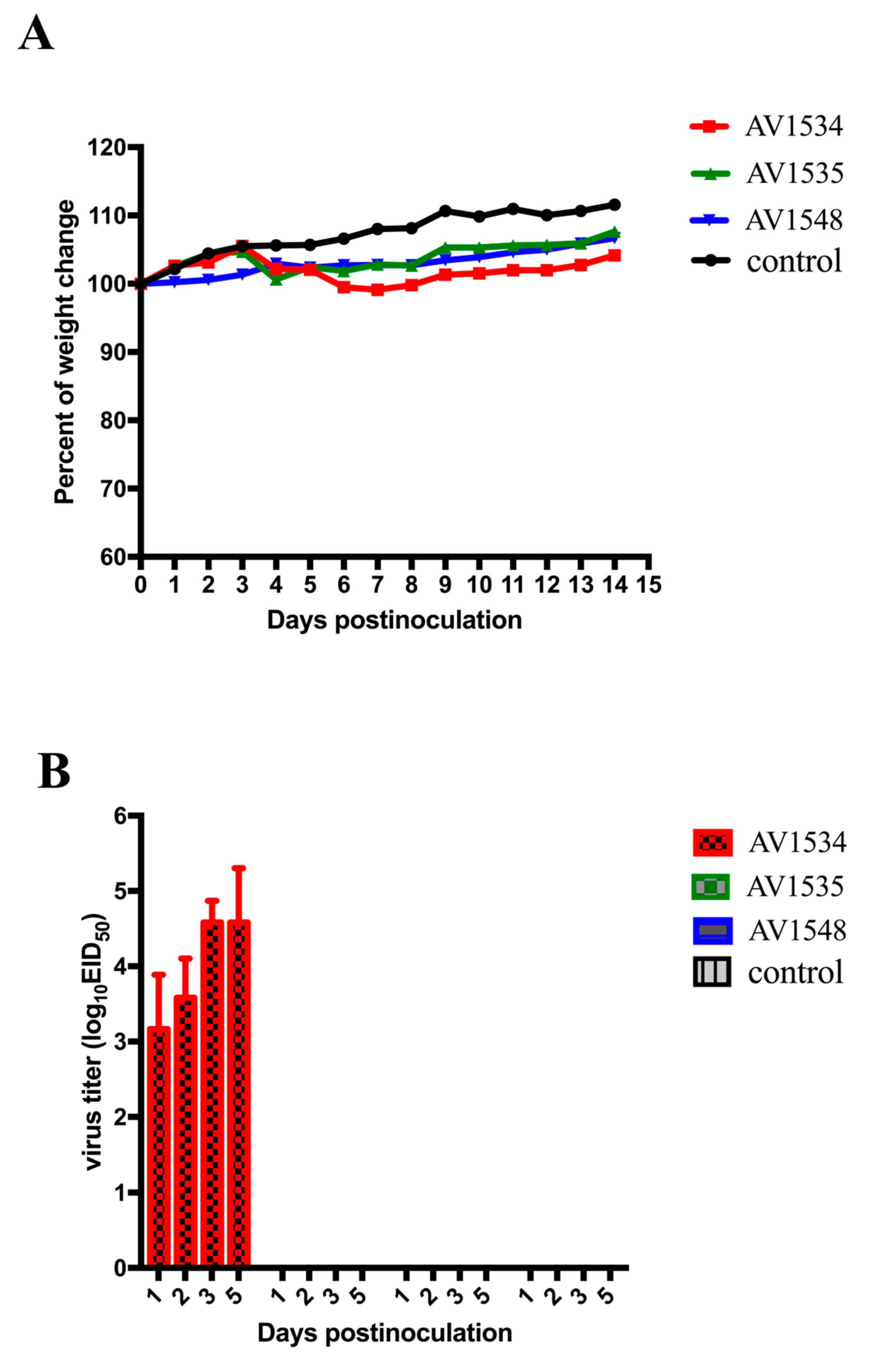
3.4. Pathogenicity of These H9N2 Viruses in Mice
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guo, Y.J.; Krauss, S.; Senne, D.A.; Mo, I.P.; Lo, K.S.; Xiong, X.P.; Norwood, M.; Shortridge, K.F.; Webster, R.G.; Guan, Y. Characterization of the pathogenicity of members of the newly established H9N2 influenza virus lineages in Asia. Virology 2000, 267, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homme, P.J.; Easterday, B.C.; Anderson, D.P. Avian influenza virus infections. II. Experimental epizootiology of influenza A-turkey-Wisconsin-1966 virus in turkeys. Avian Dis. 1970, 14, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Shortridge, K.F. Pandemic influenza: A zoonosis? Semin. Respir. Infect. 1992, 7, 11–25. [Google Scholar] [PubMed]
- Xu, K.M.; Li, K.S.; Smith, G.J.; Li, J.W.; Tai, H.; Zhang, J.X.; Webster, R.G.; Peiris, J.S.; Chen, H.; Guan, Y. Evolution and molecular epidemiology of H9N2 influenza A viruses from quail in southern China, 2000 to 2005. J. Virol. 2007, 81, 2635–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusch, E.A.; Suarez, D.L. The Multifaceted Zoonotic Risk of H9N2 Avian Influenza. Vet. Sci. 2018, 5, 82. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.; Xu, L.; Wang, X.; Liu, X. Current situation of H9N2 subtype avian influenza in China. Vet. Res. 2017, 48, 49. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.L.Z.Z.; Chen, W. The study of avian influenza: I. The isolation and preliminary serological identification of avian influenza virus in chicken. Chin. J. Vet. Med. 1994, 20, 3–5. [Google Scholar]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular characterization of H9N2 influenza viruses: Were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Tang, Y.; Liu, X.; Liu, W.; Zhang, X.; Liu, H.; Peng, D.; Gao, S.; Wu, Y.; Zhang, L.; et al. A novel genotype H9N2 influenza virus possessing human H5N1 internal genomes has been circulating in poultry in eastern China since 1998. J. Virol. 2009, 83, 8428–8438. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.M.; Smith, G.J.; Bahl, J.; Duan, L.; Tai, H.; Vijaykrishna, D.; Wang, J.; Zhang, J.X.; Li, K.S.; Fan, X.H.; et al. The genesis and evolution of H9N2 influenza viruses in poultry from southern China, 2000 to 2005. J. Virol. 2007, 81, 10389–10401. [Google Scholar] [CrossRef] [Green Version]
- Ramos, I.; Krammer, F.; Hai, R.; Aguilera, D.; Bernal-Rubio, D.; Steel, J.; Garcia-Sastre, A.; Fernandez-Sesma, A. H7N9 influenza viruses interact preferentially with alpha2,3-linked sialic acids and bind weakly to alpha2,6-linked sialic acids. J. Gen. Virol. 2013, 94, 2417–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Liu, J. H9N2 influenza virus in China: A cause of concern. Protein Cell 2015, 6, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, K.; Panei, C.J.; Shishido, M.; Noguchi, D.; Pearce, J.; Kang, H.M.; Jeong, O.M.; Lee, Y.J.; Nakanishi, K.; Ashizawa, T. SYBR green-based real-time reverse transcription-PCR for typing and subtyping of all hemagglutinin and neuraminidase genes of avian influenza viruses and comparison to standard serological subtyping tests. J. Clin. Microbiol. 2012, 50, 37–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE Avian Influenza. Available online: http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.03.04_AI.pdf (accessed on 18 February 2019).
- Thakur, A.K.; Fezio, W.L. A computer program for estimating LD50 and its confidence limits using modified Behrens-Reed-Muench cumulant method. Drug Chem. Toxicol. 1981, 4, 297–305. [Google Scholar] [CrossRef]
- Song, Y.; Cui, J.; Song, H.; Ye, J.; Zhao, Z.; Wu, S.; Xu, C.; Jiao, P.; Liao, M. New reassortant H5N8 highly pathogenic avian influenza virus from waterfowl in Southern China. Front. Microbiol. 2015, 6, 1170. [Google Scholar] [CrossRef]
- Shen, H.Q.; Yan, Z.Q.; Zeng, F.G.; Liao, C.T.; Zhou, Q.F.; Qin, J.P.; Xie, Q.M.; Bi, Y.Z.; Chen, F. Isolation and phylogenetic analysis of hemagglutinin gene of H9N2 influenza viruses from chickens in South China from 2012 to 2013. J. Vet. Sci. 2015, 16, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Yan, Z.Q.; Liu, J.; Ji, J.; Chang, S.; Liu, D.; Qin, J.P.; Ma, J.Y.; Bi, Y.Z.; Xie, Q.M. Phylogenetic analysis of hemagglutinin genes of 40 H9N2 subtype avian influenza viruses isolated from poultry in China from 2010 to 2011. Virus Genes 2012, 45, 69–75. [Google Scholar] [CrossRef]
- Su, X.; Xie, Q.; Liao, C.; Yan, Z.; Chen, W.; Bi, Y.; Chen, F. Sequence and phylogenetic analysis of hemagglutinin genes of H9N2 influenza viruses isolated from chicken in China from 2013 to 2015. J. Integr. Agric. 2016, 15, 2604–2612. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Qi, W.; Li, X.; He, J.; Jiao, P.; Zhang, C.; Liu, G.Q.; Liao, M. A single E627K mutation in the PB2 protein of H9N2 avian influenza virus increases virulence by inducing higher glucocorticoids (GCs) level. PLoS ONE 2012, 7, e38233. [Google Scholar] [CrossRef]
- Hatta, M.; Gao, P.; Halfmann, P.; Kawaoka, Y. Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science 2001, 293, 1840–1842. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, G.; Dauber, B.; Wolff, T.; Planz, O.; Klenk, H.D.; Stech, J. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. USA 2005, 102, 18590–18595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.; Ma, W.; Sun, N.; Huang, L.; Li, Y.; Zeng, Z.; Wen, Y.; Zhang, Z.; Li, H.; Li, Q.; et al. PB2-588 V promotes the mammalian adaptation of H10N8, H7N9 and H9N2 avian influenza viruses. Sci. Rep. 2016, 6, 19474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholtissek, C.; Quack, G.; Klenk, H.D.; Webster, R.G. How to overcome resistance of influenza A viruses against adamantane derivatives. Antiviral. Res. 1998, 37, 83–95. [Google Scholar] [CrossRef]
- Obenauer, J.C.; Denson, J.; Mehta, P.K.; Su, X.; Mukatira, S.; Finkelstein, D.B.; Xu, X.; Wang, J.; Ma, J.; Fan, Y.; et al. Large-scale sequence analysis of avian influenza isolates. Science 2006, 311, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.K.; Ozaki, H.; Webby, R.J.; Webster, R.G.; Peiris, J.S.; Poon, L.; Butt, C.; Leung, Y.H.; Guan, Y. Continuing evolution of H9N2 influenza viruses in Southeastern China. J. Virol. 2004, 78, 8609–8614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Xu, G.; Zhang, G.; Wen, C.; Anwar, F.; Wang, S.; Lemmon, G.; Wang, J.; Carter, R.; Wang, M.; et al. Antigenic evolution of H9N2 chicken influenza viruses isolated in China during 2009–2013 and selection of a candidate vaccine strain with broad cross-reactivity. Vet. Microbiol. 2016, 182, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Teng, Q.; Xu, D.; Shen, W.; Liu, Q.; Rong, G.; Li, X.; Yan, L.; Yang, J.; Chen, H.; Yu, H.; et al. A Single Mutation at Position 190 in Hemagglutinin Enhances Binding Affinity for Human Type Sialic Acid Receptor and Replication of H9N2 Avian Influenza Virus in Mice. J. Virol. 2016, 90, 9806–9825. [Google Scholar] [CrossRef] [Green Version]
- Li, K.S.; Xu, K.M.; Peiris, J.S.; Poon, L.L.; Yu, K.Z.; Yuen, K.Y.; Shortridge, K.F.; Webster, R.G.; Guan, Y. Characterization of H9 subtype influenza viruses from the ducks of southern China: A candidate for the next influenza pandemic in humans? J. Virol. 2003, 77, 6988–6994. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Kahn, R.E.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2008, 3, 158–166. [Google Scholar] [CrossRef]
- Li, C.; Yu, K.; Tian, G.; Yu, D.; Liu, L.; Jing, B.; Ping, J.; Chen, H. Evolution of H9N2 influenza viruses from domestic poultry in Mainland China. Virology 2005, 340, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Wang, W.; Yang, X.; Su, H.; Fan, J.; Zhu, R.; Wang, S.; Shi, H.; Liu, X. Evolution of H9N2 avian influenza virus in embryonated chicken eggs with or without homologous vaccine antibodies. BMC Vet. Res. 2018, 14, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, J.; Wang, S.; Yin, Y.; Zhang, G.; Carter, R.A.; Wang, J.; Xu, G.; Sun, H.; Wang, M.; Wen, C.; et al. Evolution of the H9N2 influenza genotype that facilitated the genesis of the novel H7N9 virus. Proc. Natl. Acad. Sci. USA 2015, 112, 548–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuoka, Y.; Swayne, D.E.; Thomas, C.; Rameix-Welti, M.A.; Naffakh, N.; Warnes, C.; Altholtz, M.; Donis, R.; Subbarao, K. Neuraminidase stalk length and additional glycosylation of the hemagglutinin influence the virulence of influenza H5N1 viruses for mice. J. Virol. 2009, 83, 4704–4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Tan, Y.; Wei, K.; Sun, H.; Shi, Y.; Pu, J.; Yang, H.; Gao, G.F.; Yin, Y.; Feng, W.; et al. Amino acid 316 of hemagglutinin and the neuraminidase stalk length influence virulence of H9N2 influenza virus in chickens and mice. J. Virol. 2013, 87, 2963–2968. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.; Xiao, H.; Chen, Q.; Wu, Y.; Fu, L.; Quan, C.; Wong, G.; Liu, J.; Haywood, J.; Liu, Y.; et al. Changes in the Length of the Neuraminidase Stalk Region Impact H7N9 Virulence in Mice. J. Virol. 2016, 90, 2142–2149. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Yu, Z.; Hu, Y.; Tu, J.; Zou, W.; Peng, Y.; Zhu, J.; Li, Y.; Zhang, A.; Yu, Z.; et al. The special neuraminidase stalk-motif responsible for increased virulence and pathogenesis of H5N1 influenza A virus. PLoS ONE 2009, 4, e6277. [Google Scholar] [CrossRef]
- Munier, S.; Larcher, T.; Cormier-Aline, F.; Soubieux, D.; Su, B.; Guigand, L.; Labrosse, B.; Cherel, Y.; Quere, P.; Marc, D.; et al. A genetically engineered waterfowl influenza virus with a deletion in the stalk of the neuraminidase has increased virulence for chickens. J. Virol. 2010, 84, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Blumenkrantz, D.; Roberts, K.L.; Shelton, H.; Lycett, S.; Barclay, W.S. The short stalk length of highly pathogenic avian influenza H5N1 virus neuraminidase limits transmission of pandemic H1N1 virus in ferrets. J. Virol. 2013, 87, 10539–10551. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Gene a | Closest Viruses in GenBank | Homology (%) |
|---|---|---|---|
| A/chicken/Shandong/AV1534/2011(H9N2) | HA | A/Chicken/Tianjin/2009(H9N2) | 97.9 |
| NA | A/Chicken/Tianjin/2009(H9N2) | 99.8 | |
| PB2 | A/chicken/Shandong/Li-2/2010(H9N2) | 98.7 | |
| PB1 | A/chicken/Shandong/01/2009(H9N2) | 98.9 | |
| PA | A/chicken/Henan/13/2008(H9N2) | 98.8 | |
| NP | A/chicken/Shandong/zc2/2009(H9N2) | 99.3 | |
| M | A/chicken/Shandong/02/2008(H9N2) | 99.9 | |
| NS | A/swine/Taizhou/5/2008(H9N2) | 99.5 | |
| A/chicken/Shandong/AV1535/2011(H9N2) | HA | A/chicken/Anhui/LJT/2010(H9N2) | 99.1 |
| NA | A/chicken/Beijing/HD/2010(H9N2) | 98.3 | |
| PB2 | A/chicken/Anhui/HF/2010(H9N2) | 98.9 | |
| PB1 | A/Duck/Fujian/1753/2009(H9N2) | 99.2 | |
| PA | A/chicken/Shanghai/96/2009(H9N2) | 98.5 | |
| NP | A/Duck/Fujian/1753/2009(H9N2) | 98.9 | |
| M | A/chicken/Shandong/10/2010(H9N2) | 99.3 | |
| NS | A/chicken/Jiangsu/Q3/2010(H9N2) | 99.4 | |
| A/chicken/Shandong/AV1548/2011(H9N2) | HA | A/chicken/Shandong/H/2009(H9N2) | 99.6 |
| NA | A/chicken/Shandong/HL/2010(H9N2) | 99.4 | |
| PB2 | A/chicken/Anhui/HF/2010(H9N2) | 99.1 | |
| PB1 | A/chicken/Hunan/2237/2010(H9N2) | 99.1 | |
| PA | A/chicken/Guangdong/1683/2009(H9N2) | 99.1 | |
| NP | A/chicken/Shandong/LY/2008(H9N2) | 98.6 | |
| M | A/chicken/Beijing/HD/2010(H9N2) A/chicken/Shanghai/96/2009(H9N2) | 99.6 | |
| NS | A/chicken/Jiangsu/Q3/2010(H9N2) | 99.5 |
| Strains | Virus Replication on Three Days Post-Inoculation (DPI) (log10EID50/0.1 mL) b | ||||
|---|---|---|---|---|---|
| Lung | Trachea | Spleen | Kidney | Brain | |
| AV1534 | 2.58 ± 1.88 | 2.17 ± 1.16 | 1.67 ± 0.14 | - c | 1.92 ± 0.52 |
| AV1535 | 4.75 ± 1.30 | 3.50 ± 0 | 2.83 ± 0.58 | 2.25 ± 0.66 | 2.25 ± 0.66 |
| AV1548 | 5.42 ± 0.14 | 3.50 ± 0 | 2.17 ± 1.16 | 2.83 ± 0.95 | 1.75 ± 0.43 |
| viruses | Virus Shedding and Mean Infectivity Titer on the Days Post-Inoculation (log10EID50/0.1 mL ± SD a) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 Day | 3 Days | 5 Days | 7 Days | 9 Days | 11 Days | 13 Days | |||||||||
| O b | C c | O | C | O | C | O | C | O | C | O | C | O | C | ||
| AV1534 | infected | 4.13 ± 0.41 (6/6) | 1.83 ± 0.44 (3/6) | 4.33 ± 0.13 (6/6) | 2.71 ± 0.70 (6/6) | 4.42 ± 0.14 (3/3) | 2.08 ± 0.52 (2/3) | 2.75 ± 0.50 (3/3) | 1.83 ± 0.38 (2/3) | ND d (0/3) | ND (0/3) | ND (0/3) | ND (0/3) | ND (0/3) | ND (0/3) |
| contact | 3.08 ± 0.29 (3/3) | ND (0/3) | 3.67 ± 0.14 (3/3) | 1.83 ± 0.58 (1/3) | 4.17 ± 0.38 (3/3) | 2.25 ± 0.43 (3/3) | 1.83 ± 0.58 (1/3) | ND (0/3) | ND (0/3) | ND (0/3) | ND (0/3) | ND (0/3) | ND (0/3) | ND (0/3) | |
| AV1535 | infected | 3.29 ± 0.29 (6/6) | 1.63 ± 0.31 (1/6) | 3.83 ± 0.59 (6/6) | 2.54 ± 0.60 (6/6) | 3.33 ± 1.04 (3/3) | 2.42 ± 0.88 (2/3) | 2.75 ± 0.43 (3/3) | 2.83 ± 1.16 (2/3) | ND (0/3) | 2.83 ± 1.16 (2/3) | ND (0/3) | 2.83 ± 1.16 (2/3) | ND (0/3) | 2.67 ± 1.01 (2/3) |
| contact | 2.92 ± 0.58 (3/3) | 192 ± 0.72 (1/3) | 3.25 ± 0.75 (3/3) | 1.75 ± 0 (3/3) | 2.25 ± 1.09 (2/3) | 2.83 ± 0.95 (3/3) | 2.42 ± 0.14 (3/3) | 2.42 ± 1.01 (2/3) | ND (0/3) | 2.17 ± 1.16 (1/3) | ND (0/3) | 2.17 ± 1.16 (1/3) | ND (0/3) | 1.58 ± 0.14 (1/3) | |
| AV1548 | infected | 3.67 ± 0.34 (6/6) | 1.58 ± 0.13 (2/6) | 4.42 ± 0.13 (6/6) | 2.25 ± 0.67 (4/6) | 4.50 ± 0 (3/3) | 2.17 ± 0.58 (2/3) | 2.42 ± 0.88 (3/3) | 2.33 ± 0.88 (2/3) | ND (0/3) | 1.83 ± 0.58 (1/3) | ND (0/3) | 1.75 ± 0.43 (1/3) | ND (0/3) | ND (0/3) |
| contact | 3.50 ± 0 (3/3) | 1.92 ± 0.72 (1/3) | 3.92 ± 0.52 (3/3) | 2.17 ± 0.58 (2/3) | 4.08 ± 0.29 (3/3) | 2.50 ± 0 (3/3) | 3.42 ± 0.14 (3/3) | 2.33 ± 0.14 (3/3) | 2.17 ± 0.58 (2/3) | ND (0/3) | 1.75 ± 0.43 (1/3) | ND (0/3) | ND (0/3) | ND (0/3) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Zhang, Y.; Chen, L.; Zhang, B.; Zhang, M.; Wang, J.; Jiang, Y.; Yang, C.; Jiang, T. Genetic Characteristics and Pathogenicity Analysis in Chickens and Mice of Three H9N2 Avian Influenza Viruses. Viruses 2019, 11, 1127. https://doi.org/10.3390/v11121127
Song Y, Zhang Y, Chen L, Zhang B, Zhang M, Wang J, Jiang Y, Yang C, Jiang T. Genetic Characteristics and Pathogenicity Analysis in Chickens and Mice of Three H9N2 Avian Influenza Viruses. Viruses. 2019; 11(12):1127. https://doi.org/10.3390/v11121127
Chicago/Turabian StyleSong, Yafen, Yong Zhang, Ling Chen, Bing Zhang, Min Zhang, Jingwen Wang, Ying Jiang, Chenghuai Yang, and Taozhen Jiang. 2019. "Genetic Characteristics and Pathogenicity Analysis in Chickens and Mice of Three H9N2 Avian Influenza Viruses" Viruses 11, no. 12: 1127. https://doi.org/10.3390/v11121127
APA StyleSong, Y., Zhang, Y., Chen, L., Zhang, B., Zhang, M., Wang, J., Jiang, Y., Yang, C., & Jiang, T. (2019). Genetic Characteristics and Pathogenicity Analysis in Chickens and Mice of Three H9N2 Avian Influenza Viruses. Viruses, 11(12), 1127. https://doi.org/10.3390/v11121127