Tilapia Lake Virus Does Not Hemagglutinate Avian and Piscine Erythrocytes and NH4Cl Does Not Inhibit Viral Replication In Vitro

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Virus Titrations
2.3. Verification of Virus Species Used for the Hemagglutination Assays
2.4. Collection and Preparation of Erythrocytes
2.5. Hemagglutination Assay
2.6. Evaluating the Effect of Ammonium Chloride on TiLV Replication in E-11 Cells
3. Results
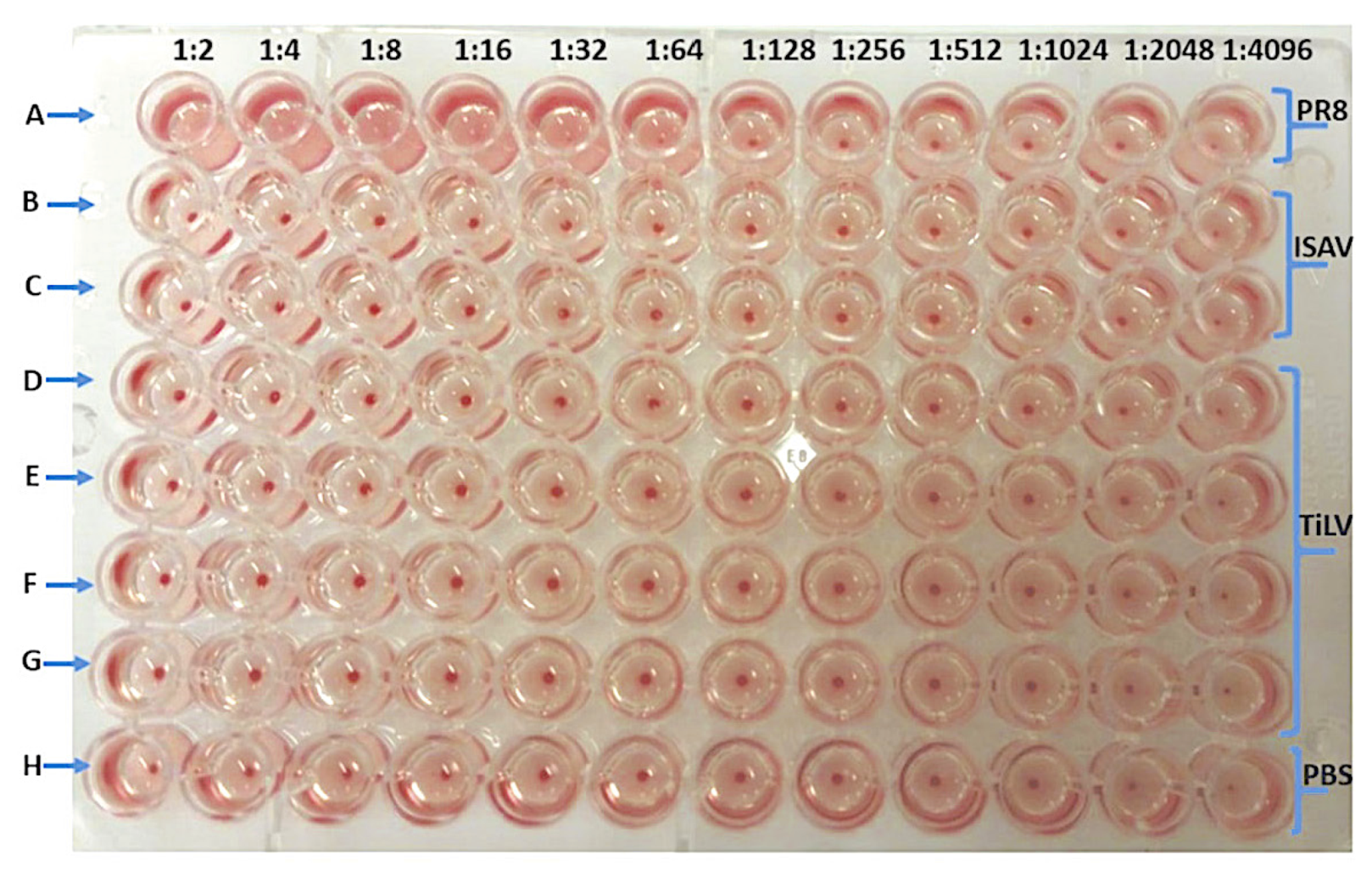
3.1. Verification of the Viruses Used in the Hemagglutination Assay
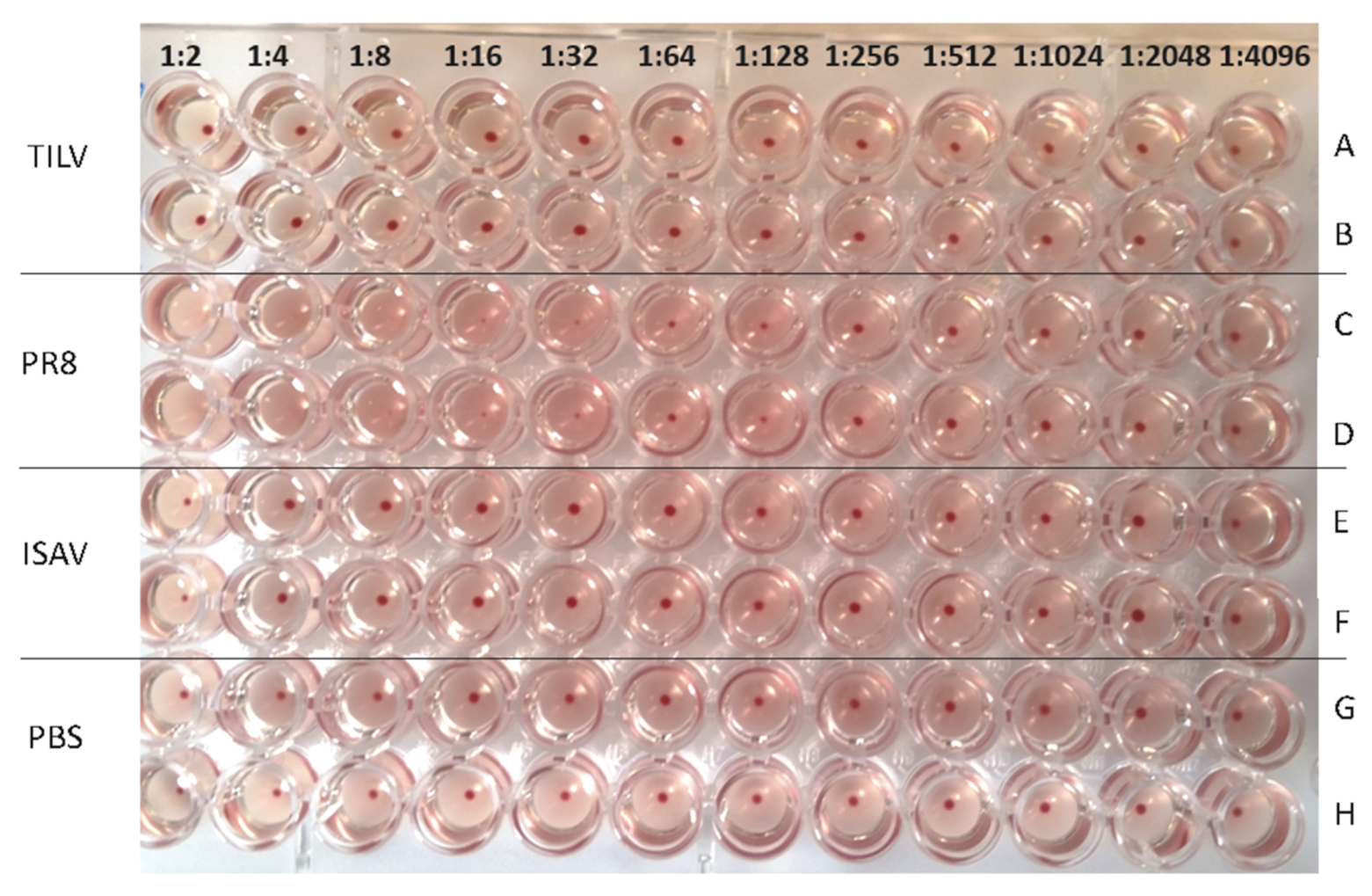
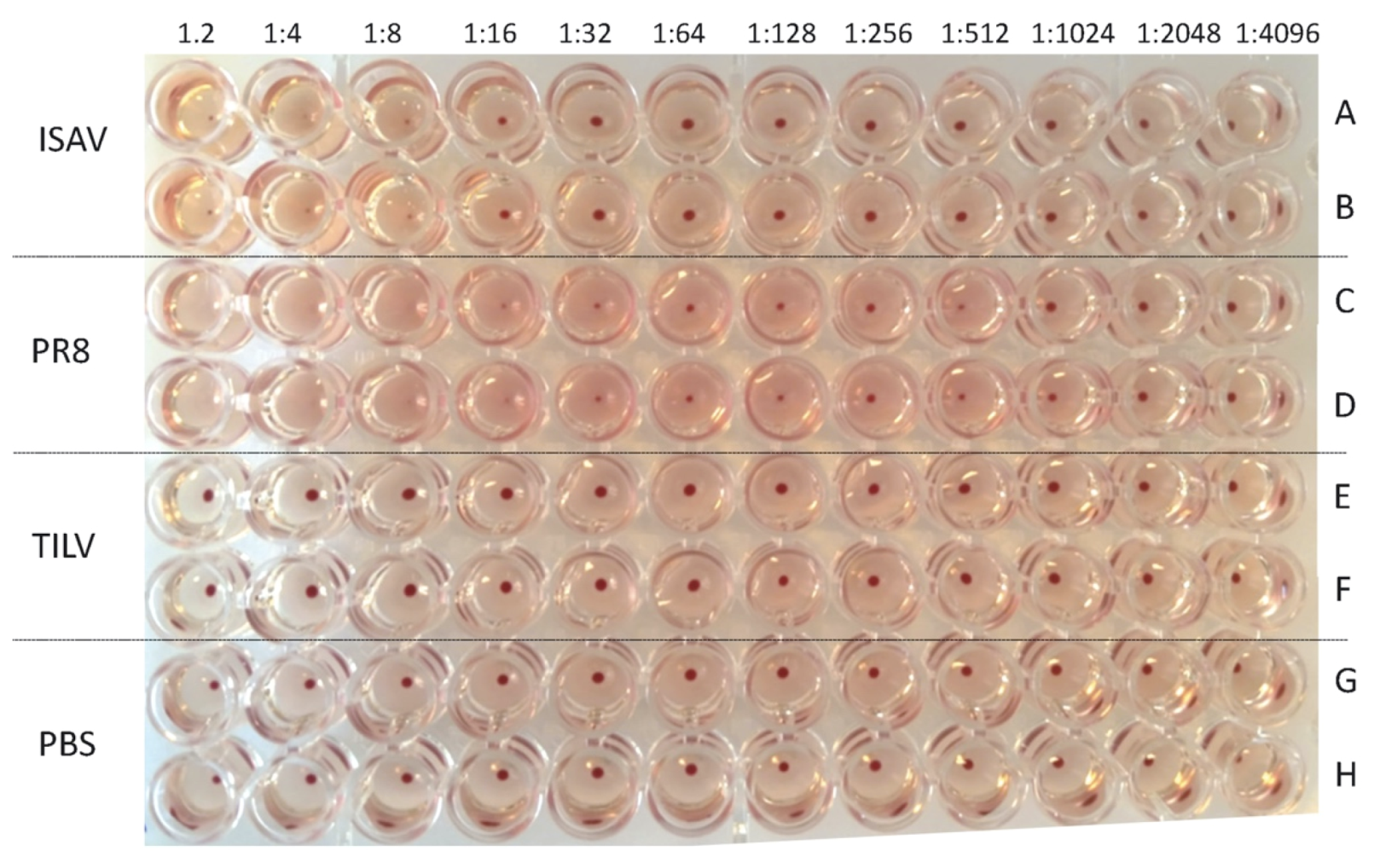
3.2. TiLV Hemagglutination Test Using Avian Erythrocytes
3.3. TiLV Hemagglutination Test Using Piscine Erythrocytes
3.4. Evaluating the Effect of Ammonium Chloride on TiLV Replication in E-11 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Eyngor, M.; Zamostiano, R.; Kembou Tsofack, J.E.; Berkowitz, A.; Bercovier, H.; Tinman, S.; Lev, M.; Hurvitz, A.; Galeotti, M.; Bacharach, E.; et al. Identification of a novel RNA virus lethal to tilapia. J. Clin. Microbiol. 2014, 52, 4137–4146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, H.; Kabuusu, R.; Beltran, S.; Reyes, E.; Lince, J.; Del Pozo, J. Syncytial hepatitis of farmed tilapia, Oreochromis niloticus (L.): A case report. J. Fish Dis. 2014, 37, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Mugimba, K.; Chengula, A.; Wamala, S.; Mwega, E.; Kasanga, C.; Byarugaba, D.; Mdegela, R.; Tal, S.; Bornstein, B.; Dishon, A. Detection of tilapia lake virus (Ti LV) infection by PCR in farmed and wild Nile tilapia (Oreochromis niloticus) from Lake Victoria. J. Fish. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, K.; Ferguson, H.W.; Kabuusu, R.; Waltzek, T.B. Genome Sequence of Tilapia Lake Virus Associated with Syncytial Hepatitis of Tilapia in an Ecuadorian Aquaculture Facility. Microbiol. Resour. Announc. 2019, 8, e00084-19. [Google Scholar] [CrossRef] [Green Version]
- Fathi, M.; Dickson, C.; Dickson, M.; Leschen, W.; Baily, J.; Muir, F.; Ulrich, K.; Weidmann, M. Identification of Tilapia Lake Virus in Egypt in Nile tilapia affected by ‘summer mortality’syndrome. Aquaculture 2017, 473, 430–432. [Google Scholar] [CrossRef]
- Bacharach, E.; Mishra, N.; Briese, T.; Zody, M.C.; Kembou Tsofack, J.E.; Zamostiano, R.; Berkowitz, A.; Ng, J.; Nitido, A.; Corvelo, A.; et al. Characterization of a Novel Orthomyxo-like Virus Causing Mass Die-Offs of Tilapia. MBio 2016, 7, e00431-16. [Google Scholar] [CrossRef] [Green Version]
- Yamkasem, J.; Tattiyapong, P.; Kamlangdee, A.; Surachetpong, W. Evidence of potential vertical transmission of tilapia lake virus. J. Fish Dis. 2019. [Google Scholar] [CrossRef]
- Nicholson, P.; Fathi, M.; Fischer, A.; Mohan, C.; Schieck, E.; Mishra, N.; Heinimann, A.; Frey, J.; Wieland, B.; Jores, J. Detection of Tilapia Lake Virus in Egyptian fish farms experiencing high mortalities in 2015. J. Fish Dis. 2017, 40, 1925–1928. [Google Scholar] [CrossRef]
- Jansen, M.D.; Mohan, C.V. Tilapia Lake Virus (TiLV): Literature Review. WorldFish: Penang, Malaysia, 2017. Available online: https://www.worldfishcenter.org/content/tilapia-lake-virus-tilv-literature-review (accessed on 11 December 2019).
- Del-Pozo, J.; Mishra, N.; Kabuusu, R.; Cheetham, S.; Eldar, A.; Bacharach, E.; Lipkin, W.; Ferguson, H. Syncytial hepatitis of tilapia (Oreochromis niloticus L.) is associated with orthomyxovirus-like virions in hepatocytes. Vet. Pathol. 2017, 54, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Tsofack, J.E.K.; Zamostiano, R.; Watted, S.; Berkowitz, A.; Rosenbluth, E.; Mishra, N.; Briese, T.; Lipkin, W.I.; Kabuusu, R.M.; Ferguson, H. Detection of Tilapia Lake Virus (TiLV) in Clinical Samples by Culturing and Nested RT-PCR. J. Clin. Microbiol. 2016, 55, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Surachetpong, W.; Janetanakit, T.; Nonthabenjawan, N.; Tattiyapong, P.; Sirikanchana, K.; Amonsin, A. Outbreaks of Tilapia Lake Virus Infection, Thailand, 2015–2016. Emerg. Infect. Dis. 2017, 23, 1031–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, B.; Pradhan, P.; Swaminathan, T.; Sood, N.; Paria, P.; Das, A.; Verma, D.; Kumar, R.; Yadav, M.; Dev, A.J. Emergence of tilapia lake virus associated with mortalities of farmed Nile tilapia Oreochromis niloticus (Linnaeus 1758) in India. Aquaculture 2018, 484, 168–174. [Google Scholar] [CrossRef]
- Wagner, R.; Feldmann, A.; Wolff, T.; Pleschka, S.; Garten, W.; Klenk, H.-D. The Role of Hemagglutinin and Neuraminidase in Influenza Virus Pathogenicity. In Structure-Function Relationships of Human Pathogenic Viruses; Holzenburg, A., Bogner, E., Eds.; Springer: Boston, MA, USA, 2002; pp. 331–345. [Google Scholar]
- Isin, B.; Doruker, P.; Bahar, I. Functional motions of influenza virus hemagglutinin: A structure-based analytical approach. Biophys. J. 2002, 82, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Al-Majhdi, F.N. Structure of the Sialic Acid Binding Site in Influenza A Virus: Hemagglutinin. J. Biol. Sci. 2007, 7, 113–122. [Google Scholar]
- Sriwilaijaroen, N.; Suzuki, Y. Molecular basis of the structure and function of H1 hemagglutinin of influenza virus. Proc. Jpn. Acad. Ser. B Bphys. Biol. Sci. 2012, 88, 226–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toennessen, R.; Lauscher, A.; Rimstad, E. Comparative aspects of infectious salmon anemia virus, an orthomyxovirus of fish, to influenza viruses. Indian J. Microbiol. 2009, 49, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Weis, W.; Brown, J.H.; Cusack, S.; Paulson, J.C.; Skehel, J.J.; Wiley, D.C. Structure of the influenza virus haemagglutinin complexed with its receptor, sialic acid. Nature 1988, 333, 426–431. [Google Scholar] [CrossRef]
- Skehel, J.J.; Wiley, D.C. Receptor Binding and Membrane Fusion in Virus Entry: The Influenza Hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Varki, A. Multiple changes in sialic acid biology during human evolution. Glycoconj. J. 2009, 26, 231–245. [Google Scholar] [CrossRef]
- Viswanathan, K.; Chandrasekaran, A.; Srinivasan, A.; Raman, R.; Sasisekharan, V.; Sasisekharan, R. Glycans as receptors for influenza pathogenesis. Glycoconj. J. 2010, 27, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Falk, K.; Namork, E.; Rimstad, E.; Mjaaland, S.; Dannevig, B.H. Characterization of infectious salmon anemia virus, an orthomyxo-like virus isolated from Atlantic salmon (Salmo salar L.). J. Virol. 1997, 71, 9016–9023. [Google Scholar] [PubMed]
- Cossart, P.; Helenius, A. Endocytosis of viruses and bacteria. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villamil Giraldo, A.M.; Appelqvist, H.; Ederth, T.; Ollinger, K. Lysosomotropic agents: Impact on lysosomal membrane permeabilization and cell death. Biochem. Soc. Trans. 2014, 42, 1460–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helenius, A. Virus entry: What has pH got to do with it? Nat. Cell Biol. 2013, 15, 125. [Google Scholar] [CrossRef]
- Munang’andu, H.M.; Sandtro, A.; Mutoloki, S.; Brudeseth, B.E.; Santi, N.; Evensen, O. Immunogenicity and cross protective ability of the central VP2 amino acids of infectious pancreatic necrosis virus in Atlantic salmon (Salmo salar L.). PLoS ONE 2013, 8, e54263. [Google Scholar] [CrossRef] [Green Version]
- Mugimba, K.K.; Tal, S.; Dubey, S.; Mutoloki, S.; Dishon, A.; Evensen, O.; Munang’andu, H.M. Gray (Oreochromis niloticus x O. aureus) and Red (Oreochromis spp.) Tilapia Show Equal Susceptibility and Proinflammatory Cytokine Responses to Experimental Tilapia Lake Virus Infection. Viruses 2019, 11, 893. [Google Scholar] [CrossRef] [Green Version]
- Daum, L.T.; Canas, L.C.; Arulanandam, B.P.; Niemeyer, D.; Valdes, J.J.; Chambers, J.P. Real-time RT-PCR assays for type and subtype detection of influenza A and B viruses. Influenza Respir. Viruses 2007, 1, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Snow, M.; McKay, P.; McBeath, A.J.; Black, J.; Doig, F.; Kerr, R.; Cunningham, C.O.; Nylund, A.; Devold, M. Development, application and validation of a Taqman real-time RT-PCR assay for the detection of infectious salmon anaemia virus (ISAV) in Atlantic salmon (Salmo salar). Dev. Biol. 2006, 126, 133–145. [Google Scholar]
- Conrad, S.K.; Narwold, D.R.; Srinivas, G.B. Detection of Hemagglutinating Viruses; United States Department of Health and Human Services, Ed.; United States Department of Agriculture Animal and Plant Health Inspection Service: Ames, IA, USA, 2018; pp. 1–21.
- Munang’andu, H.M.; Fredriksen, B.N.; Mutoloki, S.; Dalmo, R.A.; Evensen, Ø. The kinetics of CD4+ and CD8+ T-cell gene expression correlate with protection in Atlantic salmon (Salmo salar L.) vaccinated against infectious pancreatic necrosis. Vaccine 2013, 31, 1956–1963. [Google Scholar]
- Munang’andu, H.M.; Fredriksen, B.N.; Mutoloki, S.; Brudeseth, B.; Kuo, T.-Y.; Marjara, I.S.; Dalmo, R.A.; Evensen, Ø. Comparison of vaccine efficacy for different antigen delivery systems for infectious pancreatic necrosis virus vaccines in Atlantic salmon (Salmo salar L.) in a cohabitation challenge model. Vaccine 2012, 30, 4007–4016. [Google Scholar]
- Gollins, S.W.; Porterfield, J.S. The uncoating and infectivity of the flavivirus West Nile on interaction with cells: Effects of pH and ammonium chloride. J. Gen. Virol. 1986, 67, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Ohnishi, S. Uncoating of influenza virus in endosomes. J. Virol. 1984, 51, 497–504. [Google Scholar] [PubMed]
- Dermody, T.; Nibert, M.; Wetzel, J.; Tong, X.; Fields, B.N. Cells and viruses with mutations affecting viral entry are selected during persistent infections of L cells with mammalian reoviruses. J. Virol. 1993, 67, 2055–2063. [Google Scholar] [PubMed]
- Ito, T.; Couceiro, J.N.; Kelm, S.; Baum, L.G.; Krauss, S.; Castrucci, M.R.; Donatelli, I.; Kida, H.; Paulson, J.C.; Webster, R.G.; et al. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J. Virol. 1998, 72, 7367–7373. [Google Scholar] [PubMed]
- Gamblin, S.J.; Skehel, J.J. Influenza hemagglutinin and neuraminidase membrane glycoproteins. J. Biol. Chem. 2010, 285, 28403–28409. [Google Scholar] [CrossRef] [Green Version]
- Herrler, G.; Hausmann, J.; Klenk, H.-D. Sialic acid as receptor determinant of ortho-and paramyxoviruses. In Biology of the Sialic Acids; Springer: Berlin/Heidelberg, Germany, 1995; pp. 315–336. [Google Scholar]
- Zhang, Q.; Wang, Y.; Zheng, Q.; Li, J. Analysis of O-acetylated sialic acids in dried blood spots. Anal. Chem. 2019, 4, 2744–2751. [Google Scholar] [CrossRef]
- Ghosh, S. Sialic acid binding lectins (SABL) from Mollusca, a review and insilico study of SABL from Solen grandis and Limax flavus. Entomol. Zool. Stud. 2017, 5, 1563–1572. [Google Scholar]
- Hanaoka, K.; Pritchett, T.J.; Takasaki, S.; Kochibe, N.; Sabesan, S.; Paulson, J.C.; Kobata, A. 4-O-acetyl-N-acetylneuraminic acid in the N-linked carbohydrate structures of equine and guinea pig alpha 2-macroglobulins, potent inhibitors of influenza virus infection. J. Biol. Chem. 1989, 264, 9842–9849. [Google Scholar]
- Hara, S.; Yamaguchi, M.; Takemori, Y.; Furuhata, K.; Ogura, H.; Nakamura, M. Determination of mono-O-acetylatedN-acetylneuraminic acids in human and rat sera by fluorometric high-performance liquid chromatography. Anal. Biochem. 1989, 179, 162–166. [Google Scholar] [CrossRef]
- Jayo, R.G.; Li, J.; Chen, D.D.Y. Capillary electrophoresis mass spectrometry for the characterization of O-acetylated N-glycans from fish serum. Anal. Chem. 2012, 84, 8756–8762. [Google Scholar] [CrossRef]
- Liu, X.; Afonso, L.; Altman, E.; Johnson, S.; Brown, L.; Li, J. O-acetylation of sialic acids in N-glycans of Atlantic salmon (Salmo salar) serum is altered by handling stress. Proteomics 2008, 8, 2849–2857. [Google Scholar] [CrossRef] [PubMed]
- Sato, C.; Kitajima, K.; Tazawa, I.; Inoue, Y.; Inoue, S.; Troy, F.A. Structural diversity in the alpha 2--> 8-linked polysialic acid chains in salmonid fish egg glycoproteins. Occurrence of poly (Neu5Ac), poly (Neu5Gc), poly (Neu5Ac, Neu5Gc), poly (KDN), and their partially acetylated forms. J. Biol. Chem. 1993, 268, 23675–23684. [Google Scholar] [PubMed]
- Iwersen, M.; Vandamme-Feldhaus, V.; Schauer, R. Enzymatic 4-O-acetylation of N-acetylneuraminic acid in guinea-pig liver. Glycoconj. J. 1998, 15, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Yeşilyurt, B.; Şahar, U.; Deveci, R. Determination of the type and quantity of sialic acid in the egg jelly coat of the sea urchin Paracentrotus lividus using capillary LC-ESI-MS/MS. Mol. Reprod. Dev. 2015, 82, 115–122. [Google Scholar] [CrossRef]
- Spik, G.; Coddeville, B.; Montreuil, J.J.B. Comparative study of the primary structures of sero-, lacto-and ovotransferrin glycans from different species. Biochimie 1988, 70, 1459–1469. [Google Scholar] [CrossRef]
- Schwegmann-Weßels, C.; Herrler, G. Sialic acids as receptor determinants for coronaviruses. Glycoconj. J. 2006, 23, 51–58. [Google Scholar] [CrossRef]
- Cook, J.D.; Sultana, A.; Lee, J.E. Structure of the infectious salmon anemia virus receptor complex illustrates a unique binding strategy for attachment. Proc. Natl. Acad. Sci. USA 2017, 114, E2929–E2936. [Google Scholar] [CrossRef] [Green Version]
- Hellebø, A.; Vilas, U.; Falk, K.; Vlasak, R. Infectious salmon anemia virus specifically binds to and hydrolyzes 4-O-acetylated sialic acids. J. Virol. 2004, 78, 3055–3062. [Google Scholar] [CrossRef] [Green Version]
- Aamelfot, M.; Dale, O.B.; Weli, S.C.; Koppang, E.O.; Falk, K. Expression of the infectious salmon anemia virus receptor on atlantic salmon endothelial cells correlates with the cell tropism of the virus. J. Virol. 2012, 86, 10571–10578. [Google Scholar] [CrossRef] [Green Version]
- Weissenhorn, W.; Dessen, A.; Calder, L.; Harrison, S.; Skehel, J.; Wiley, D.C. Structural basis for membrane fusion by enveloped viruses. Mol. Membr. Biol. 1999, 16, 3–9. [Google Scholar] [CrossRef]
- Leung, J.Y.-S.; Ng, M.M.-L.; Chu, J.J.H. Replication of Alphaviruses: A Review on the Entry Process of Alphaviruses into Cells. Adv. Virol. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, A.H.; Takahiro, U. The effect of ammonium chloride on the multiplication of herpes simplex virus type 1 in Vero cells. Virus Res. 1989, 13, 271–281. [Google Scholar] [CrossRef]
- Canning, W.M.; Fields, B.N. Ammonium chloride prevents lytic growth of reovirus and helps to establish persistent infection in mouse L cells. Science 1983, 219, 987–988. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Hingley, S.T.; Simmons, G.; Yu, C.; Das Sarma, J.; Bates, P.; Weiss, S.R. Endosomal Proteolysis by Cathepsins Is Necessary for Murine Coronavirus Mouse Hepatitis Virus Type 2 Spike-Mediated Entry. J. Virol. 2006, 80, 5768–5776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlin, K.S.; Reggio, H.; Helenius, A.; Simons, K. Infectious entry pathway of influenza virus in a canine kidney cell line. J. Cell Biol. 1981, 91, 601–613. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | Expected Product (bp) | Virus/Segment | Reference |
|---|---|---|---|---|
| TiLV-SG3-F4 | TCCAGATCACCCTTCCTACTT | 109 | TiLV/3 | This study |
| TiLV-SG3-R4 | ATCCCAAGCAATCGGCTAAT | |||
| Flu A-F | TAACCGAGGTCGAAACGTA | 195 | PR8/7 | [29] |
| Flu A-R | GCACGGTGAGCGTGAA | |||
| H520-F | CTACACAGCAGGATGCAGATGT | 104 | ISAV/8 | [30] |
| H534-R | CAGGATGCCGGAAGTCGAT |
| Viruses | Hemagglutination Titre (Dilution Factor/50 μL) of Avian and Piscine Erythrocytes | ||
|---|---|---|---|
| Turkey RBCs | Tilapia RBCs | Atlantic Salmon RBCs | |
| PR8 | 1:128 | 1:64 | 1:512 |
| ISAV | 0 | 1:4 | 1:16 |
| TiLV | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chengula, A.A.; Mutoloki, S.; Evensen, Ø.; Munang’andu, H.M. Tilapia Lake Virus Does Not Hemagglutinate Avian and Piscine Erythrocytes and NH4Cl Does Not Inhibit Viral Replication In Vitro. Viruses 2019, 11, 1152. https://doi.org/10.3390/v11121152
Chengula AA, Mutoloki S, Evensen Ø, Munang’andu HM. Tilapia Lake Virus Does Not Hemagglutinate Avian and Piscine Erythrocytes and NH4Cl Does Not Inhibit Viral Replication In Vitro. Viruses. 2019; 11(12):1152. https://doi.org/10.3390/v11121152
Chicago/Turabian StyleChengula, Augustino Alfred, Stephen Mutoloki, Øystein Evensen, and Hetron Mweemba Munang’andu. 2019. "Tilapia Lake Virus Does Not Hemagglutinate Avian and Piscine Erythrocytes and NH4Cl Does Not Inhibit Viral Replication In Vitro" Viruses 11, no. 12: 1152. https://doi.org/10.3390/v11121152
APA StyleChengula, A. A., Mutoloki, S., Evensen, Ø., & Munang’andu, H. M. (2019). Tilapia Lake Virus Does Not Hemagglutinate Avian and Piscine Erythrocytes and NH4Cl Does Not Inhibit Viral Replication In Vitro. Viruses, 11(12), 1152. https://doi.org/10.3390/v11121152