Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Animals
2.3. Clinical and Necropsy Assessment
2.4. Histopathology and Morphometry
2.5. qRT-PCR
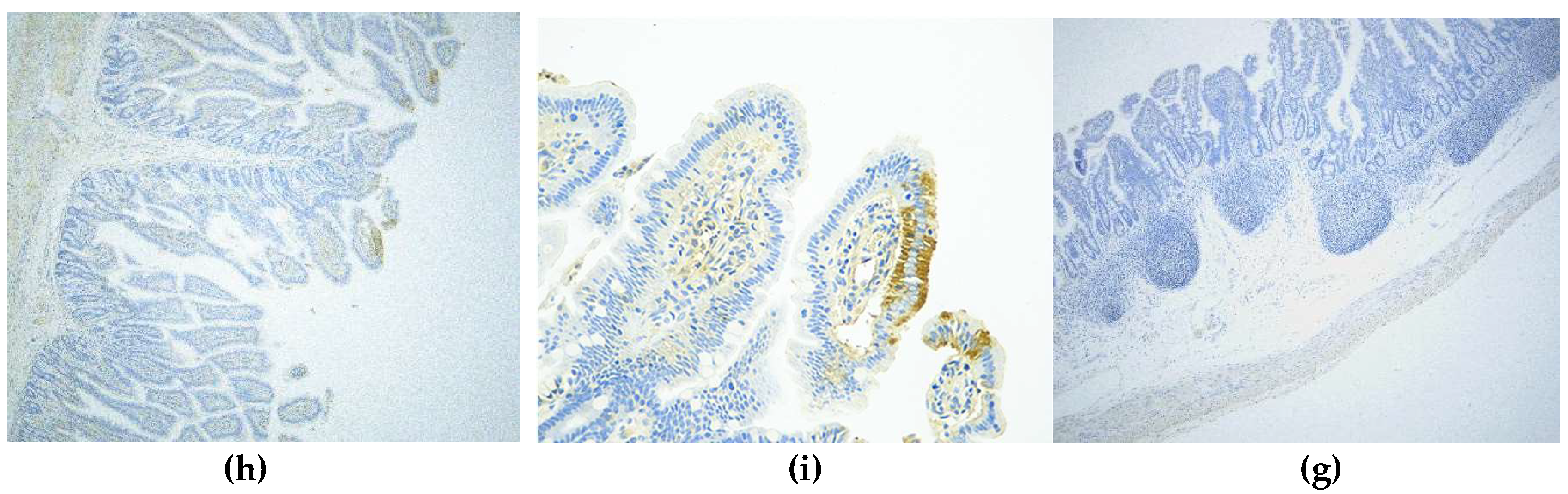
2.6. Immunohistochemistry (IHC)
2.7. Statistical Analysis
3. Results
3.1. Infection of Pigs with PAstV1-GX1 Results in Mild Diarrhea
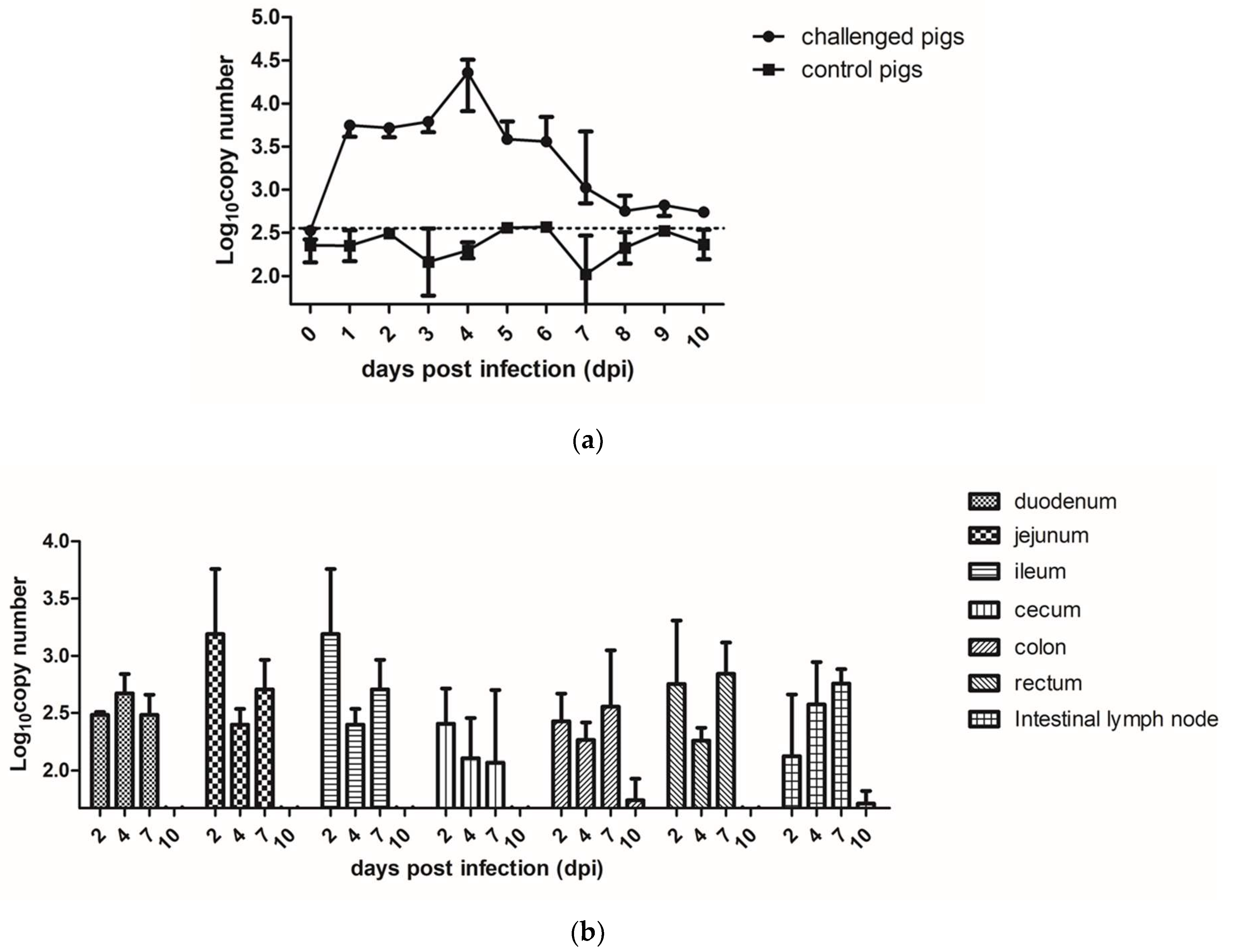
3.2. Fecal Shedding, Viral Load, and Tissue Distribution in PAstV-Inoculated Pigs
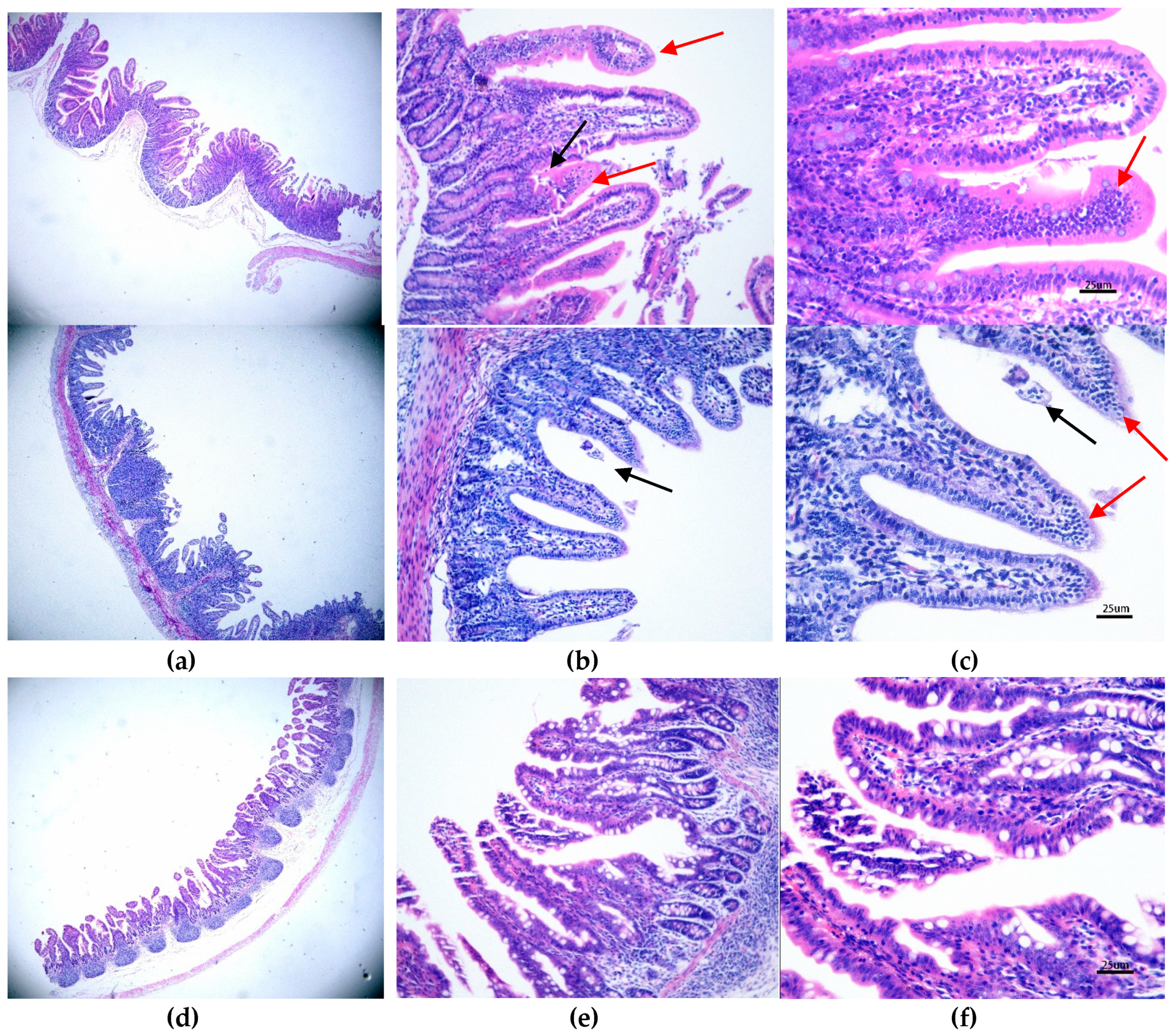
3.3. Histopathological and Morphometric Changes Induced by PAstV1-GX1 Infection
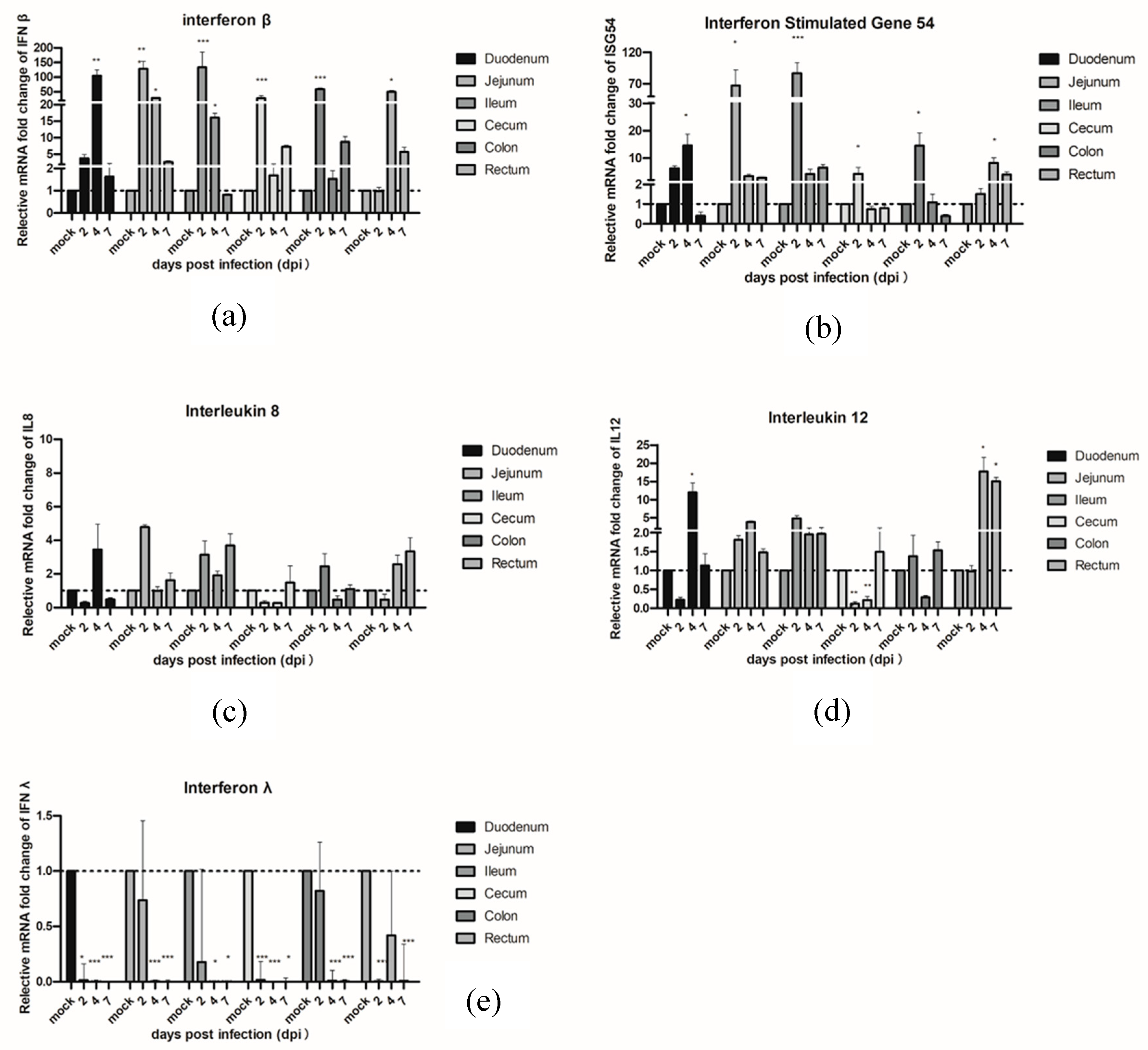
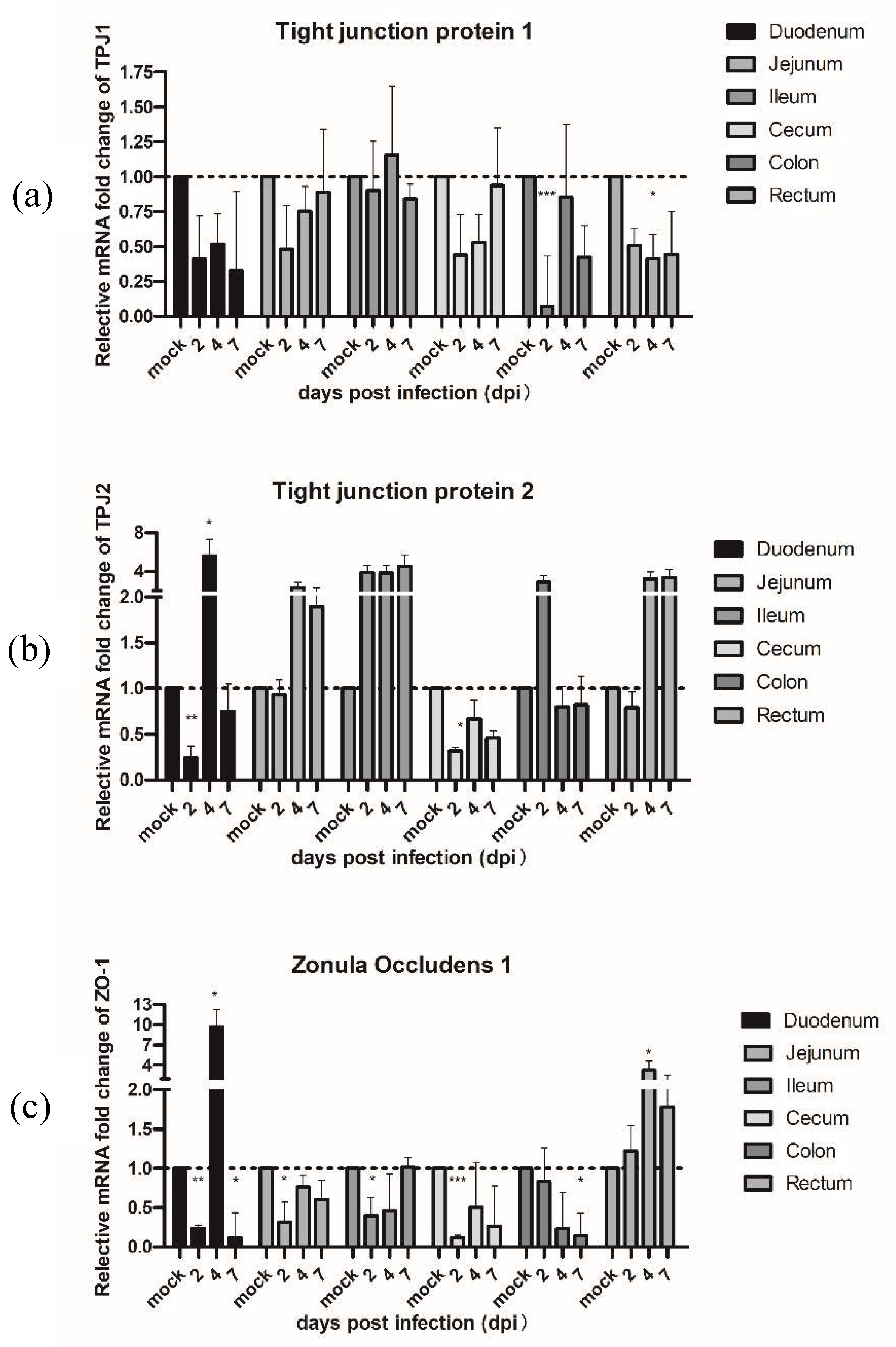
3.4. The Effects of PAstV on Transcriptional Activation of Inflammatory Cytokines and Permeable Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benedictis, P.D.; Schultz-Cherry, S.; Burnham, A.; Cattoli, G. Astrovirus infections in humans and animals–Molecular biology, genetic diversity, and interspecies transmissions. Infect. Genet. Evol. 2011, 11, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G.; Pankovics, P.; Delwart, E.; Boros, Á. Identification of a novel astrovirus in domestic sheep in Hungary. Arch. Virol. 2011, 156, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.C. Detection by electron microscopy of caliciviruses, astroviruses and rotavirus-like particles in the faeces of piglets with diarrhoea. Vet. Rec. 1980, 107, 532–533. [Google Scholar] [PubMed]
- Brnić, D.; Jemeršić, L.; Keros, T.; Prpić, J. High prevalence and genetic heterogeneity of porcine astroviruses in domestic pigs. Vet. J. 2014, 202, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yin, W.; Zhou, Y.; Li, B.; Ai, L.; Pan, M.; Guo, W. Molecular detection of Porcine astrovirus in Sichuan Province, China. Virol. J. 2016, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.T.; Giménez-Lirola, L.G.; Gerber, P.F.; Jiang, Y.H.; Halbur, P.G.; Opriessnig, T. Identification and characterization of novel porcine astroviruses (PAstVs) with high prevalence and frequent co-infection of individual pigs with multiple PAstV types. J. Gen. Virol. 2013, 94, 570. [Google Scholar] [CrossRef]
- Zhou, W.; Ullman, K.; Chowdry, V.; Reining, M.; Benyeda, Z.; Baule, C.; Juremalm, M.; Wallgren, P.; Schwarz, L.; Zhou, E. Molecular investigations on the prevalence and viral load of enteric viruses in pigs from five European countries. Vet. Microbiol. 2016, 182, 75–81. [Google Scholar] [CrossRef]
- Lan, D.; Ji, W.; Shan, T.; Cui, L.; Yang, Z.; Yuan, C.; Hua, X. Molecular characterization of a porcine astrovirus strain in China. Arch. Virol. 2011, 156, 1869–1875. [Google Scholar] [CrossRef]
- Ashley, C.R.; Caul, E.O.; Paver, W.K. Astrovirus-associated gastroenteritis in children. Lancet 1975, 306, 559. [Google Scholar] [CrossRef] [Green Version]
- Cortez, V.; Meliopoulos, V.A.; Karlsson, E.A.; Hargest, V.; Johnson, C.; Schultz-Cherry, S. Astrovirus biology and pathogenesis. Annu. Rev. Virol. 2017, 4, 327–348. [Google Scholar] [CrossRef]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendenhall, I.H.; Smith, G.J.; Vijaykrishna, D. Ecological drivers of virus evolution: Astrovirus as a case study. J. Virol. 2015, 89, 6978–6981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, E.A.; Small, C.T.; Freiden, P.; Feeroz, M.M.; Matsen, T.F.; San, S.; Hasan, M.K.; Wang, D.; Jonesengel, L.; Schultzcherry, S. Non-human primates harbor diverse mammalian and avian astroviruses including those associated with human infections. PLoS Pathog. 2015, 11, e1005225. [Google Scholar] [CrossRef] [PubMed]
- Pankovics, P.; Boros, A.; Kiss, T.; Delwart, E.; Reuter, G. Detection of a mammalian-like astrovirus in bird, European roller (Coracias garrulus). Infect. Genet. Evol. 2015, 34, 114–121. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Le, B.M.; Holtz, L.R.; Storch, G.A.; Wang, D. Detection of newly described astrovirus MLB1 in stool samples from children. Emerg. Infect. Dis. 2009, 15, 441–444. [Google Scholar] [CrossRef] [Green Version]
- Rivera, R.; Nollens, H.H.; Venn-Watson, S.; Gulland, F.M.; Wellehan, F.X., Jr. Characterization of phylogenetically diverse astroviruses of marine mammals. J. Gen. Virol. 2010, 91, 166. [Google Scholar] [CrossRef]
- Johnson, C.; Hargest, V.; Cortez, V.; Meliopoulos, V.A.; Stacey, S.-C. Astrovirus pathogenesis. Viruses 2017, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Indik, S.; Valíček, L.; Šmíd, B.; Dvořáková, H.; Rodák, L. Isolation and partial characterization of a novel porcine astrovirus. Vet. Microbiol. 2006, 117, 276–283. [Google Scholar] [CrossRef]
- Mor, S.K.; Chander, Y.; Marthaler, D.; Patnayak, D.P.; Goyal, S.M. Detection and molecular characterization of Porcine astrovirus strains associated with swine diarrhea. J. Vet. Diagn. Investig. 2012, 24, 1064–1067. [Google Scholar] [CrossRef] [Green Version]
- Shan, T.; Li, L.; Simmonds, P.; Wang, C.; Moeser, A.; Delwart, E. The fecal virome of pigs on a high-density farm. J. Virol. 2011, 85, 11697. [Google Scholar] [CrossRef] [Green Version]
- Brnić, D.; Prpić, J.; Keros, T.; Roić, B.; Starešina, V.; Jemeršić, L. Porcine astrovirus viremia and high genetic variability in pigs on large holdings in Croatia. Infect. Genet. Evol. 2013, 14, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Arruda, B.; Arruda, P.; Hensch, M.; Chen, Q.; Zheng, Y.; Yang, C.; Irh, G.; Ferreyra, F.M.; Gauger, P.; Schwartz, K. Porcine astrovirus type 3 in central nervous system of swine with polioencephalomyelitis. Emerg. Infect. Dis. 2017, 23, 2097–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boros, Á.; Albert, M.; Pankovics, P.; Bíró, H.; Pesavento, P.A.; Phan, T.G.; Delwart, E.; Reuter, G. Outbreaks of neuroinvasive astrovirus associated with encephalomyelitis, weakness, and paralysis among weaned pigs, Hungary. Emerg. Infect. Dis. 2017, 23, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Behling-kelly, E.; Schultz-cherry, S.; Koci, M.; Kelley, L.; Larsen, D.; Brown, C. Localization of astrovirus in experimentally infected turkeys as determined by in situ hybridization. Vet. Pathol. 2002, 39, 595–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Wang, F.; Shi, J.; Zheng, L.; Wang, X.; Zhang, D. Molecular characterization of a duck hepatitis virus 3-like astrovirus. Vet. Microbiol. 2014, 170, 39–47. [Google Scholar] [CrossRef]
- Cordey, S.; Vu, D.L.; Schibler, M.; L’Huillier, A.G.; Brito, F.; Docquier, M.; Posfay-Barbe, K.M.; Petty, T.J.; Turin, L.; Zdobnov, E.M. Astrovirus mlb2, a new gastroenteric virus associated with meningitis and disseminated infection. Emerg. Infect. Dis. 2016, 22, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.R.; Morfopoulou, S.; Hubb, J.; Emmett, W.A.; Ip, W.; Shah, D.; Brooks, T.; Paine, S.M.L.; Anderson, G.; Virasami, A. Astrovirus va1/hmo-c: An increasingly recognized neurotropic pathogen in immunocompromised patients. Clin. Infect. Dis. 2015, 60, 881–888. [Google Scholar] [CrossRef]
- Selimovic-Hamza, S.; Boujon, C.L.; Hilbe, M.; Oevermann, A.; Seuberlich, T. Frequency and pathological phenotype of bovine astrovirus ch13/neuros1 infection in neurologically-diseased cattle: Towards assessment of causality. Viruses 2017, 9, 12. [Google Scholar] [CrossRef]
- Blomström, A.L.; Widén, F.; Hammer, A.S.; Belák, S.; Berg, M. Detection of a novel astrovirus in brain tissue of mink suffering from shaking mink syndrome by use of viral metagenomics. J. Clin. Microbiol. 2010, 48, 4392–4396. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Fang, Q.; Liu, H.; Ji, C.; Chen, Y.; Ouyang, K.; Wei, Z.; Huang, W. Construction of a reverse genetic system for porcine astrovirus. Arch. Virol. 2018, 163, 1511–1518. [Google Scholar] [CrossRef]
- McCarty, K.S., Jr.; Miller, L.S.; Cox, E.B.; Konrath, J.; McCarty, K.S., Sr. Estrogen receptor analyses. Correlation of biochemical and immunohistochemical methods using monoclonal antireceptor antibodies. Arch. Pathol. Lab. Med. 1985, 109, 716–721. [Google Scholar] [PubMed]
- Luo, Z.; Roi, S.; Dastor, M.; Gallice, E.; Laurin, M.A.; L’Homme, Y. Multiple novel and prevalent astroviruses in pigs. Vet. Microbiol. 2011, 149, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Monini, M.; Bartolo, I.D.; Ianiro, G.; Angeloni, G.; Magistrali, C.F.; Ostanello, F.; Ruggeri, F.M. Detection and molecular characterization of zoonotic viruses in swine fecal samples in Italian pig herds. Arch. Virol. 2015, 160, 2547–2556. [Google Scholar] [CrossRef] [PubMed]
- Machnowska, P.; Ellerbroek, L.; Johne, R. Detection and characterization of potentially zoonotic viruses in faeces of pigs at slaughter in Germany. Vet. Microbiol. 2014, 168, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Kumthip, K.; Khamrin, P.; Saikruang, W.; Kongkaew, A.; Vachirachewin, R.; Ushijima, H.; Maneekarn, N. Detection and genetic characterization of porcine astroviruses in piglets with and without diarrhea in Thailand. Arch. Virol. 2018, 163, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Jeoung, H.Y.; Park, H.R.; Lim, J.A.; Song, J.Y.; An, D.J. Phylogenetic analysis of porcine astrovirus in domestic pigs and wild boars in South Korea. Virus Genes 2013, 46, 175–181. [Google Scholar] [CrossRef]
- Xiao, C.T.; Luo, Z.; Lv, S.L.; Opriessnig, T.; Li, R.C.; Yu, X.L. Identification and characterization of multiple porcine astrovirus genotypes in Hunan province, China. Arch. Virol. 2016, 162, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Snodgrass, D.R.; Gray, E.W. Detection and transmission of 30 nm virus particles (astroviruses) in faeces of lambs with diarrhoea. Arch. Virol. 1977, 55, 287–291. [Google Scholar] [CrossRef]
- Tzipori, S.; Menzies, J.D.; Gray, E.W. Detection of astrovirus in the faeces of red deer. Vet. Rec. 1981, 108, 286. [Google Scholar] [CrossRef]
- Woode, G.N.; Bridger, J.C. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J. Med Microbiol. 1978, 11, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Bosch, A.; Guix, S.; Pintó, R.M. Epidemiology of Human Astroviruses; Springer: New York, NY, USA, 2012. [Google Scholar] [CrossRef]
- Méndez-Toss, M.; Griffin, D.D.; Calva, J.; Contreras, J.F.; Puerto, F.I.; Mota, F.; Guiscafré, H.; Cedillo, R.; Muñoz, O.; Herrera, I. Prevalence and genetic diversity of human astroviruses in Mexican children with symptomatic and asymptomatic infections. J. Clin. Microbiol. 2004, 42, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koci, M.D.; Moser, L.A.; Kelley, L.A.; Diane, L.; Brown, C.C.; Stacey, S.C. Astrovirus induces diarrhea in the absence of inflammation and cell death. J. Virol. 2003, 77, 11798–11808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, C.C.; Loh, J.T.; Zhao, G.; Duan, E.; Freiden, P.; Heath, R.J.; Schultz-Cherry, S.; Stappenbeck, T.S.; Wang, D.; Huang, H.V. Adaptive immunity restricts replication of novel murine astroviruses. J. Virol. 2012, 86, 12262–12270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.L.; Cordey, S.; Brito, F.; Kaiser, L. Novel human astroviruses: Novel human diseases? J. Clin. Virol. 2016, 82, 56–63. [Google Scholar] [CrossRef]
- Quan, P.L.; Wagner, T.A.; Briese, T.; Torgerson, T.R.; Hornig, M.; Tashmukhamedova, A.; Firth, C.; Palacios, G.; Baisre-De-Leon, A.; Paddock, C.D. Astrovirus encephalitis in boy with X-linked agammaglobulinemia. Emerg. Infect. Dis. 2010, 16, 918–925. [Google Scholar] [CrossRef]
- Bouzalas, I.G.; Daniel, W.; Julia, W.; Cord, D.G.; Andreas, Z.; Marc, V.; Anna, O.; Rémy, B.; Torsten, S. Neurotropic astrovirus in cattle with nonsuppurative encephalitis in Europe. J. Clin. Microbiol. 2014, 52, 3318–3324. [Google Scholar] [CrossRef] [Green Version]
- Schlottau, K.; Schulze, C.; Bilk, S.; Hanke, D.; Höper, D.; Beer, M.; Hoffmann, B. Detection of a Novel Bovine Astrovirus in a Cow with Encephalitis. Transbound. Emerg. Dis. 2016, 63, 253–259. [Google Scholar] [CrossRef]
- Pfaff, F.; Schlottau, K.; Scholes, S.; Courtenay, A.; Hoffmann, B.; Höper, D.; Beer, M. A novel astrovirus associated with encephalitis and ganglionitis in domestic sheep. Transbound. Emerg. Dis. 2017, 64, 677–682. [Google Scholar] [CrossRef]
- Marvin, S.A.; Huerta, C.T.; Sharp, B.; Freiden, P.; Cline, T.D.; Schultz-Cherry, S. Type I interferon response limits astrovirus replication and protects against increased barrier permeability in vitro and in vivo. J. Virol. 2016, 90, 1988–1996. [Google Scholar] [CrossRef] [Green Version]
- Guix, S.; Perez-Bosque, A.; Miro, L.; Moreto, M.; Bosch, A.; Pinto, R.M. Type I interferon response is delayed in human astrovirus infections. PLoS ONE 2015, 10, e0123087. [Google Scholar] [CrossRef]
- Wang, D.; Fang, L.; Liu, L.; Zhong, H.; Chen, Q.; Luo, R.; Liu, X.; Zhang, Z.; Chen, H.; Xiao, S. Foot-and-mouth disease virus (FMDV) leader proteinase negatively regulates the porcine interferon-lambda1 pathway. Mol. Immunol. 2011, 49, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Meliopoulos, V.A.; Marvin, S.A.; Freiden, P.; Moser, L.A.; Nighot, P.; Ali, R.; Blikslager, A.; Reddivari, M.; Heath, R.J.; Koci, M.D. Oral administration of astrovirus capsid protein is sufficient to induce acute diarrhea in vivo. Mbio 2016, 7, e01494-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, L.A.; Michael, C.; Stacey, S.C. Astrovirus increases epithelial barrier permeability independently of viral replication. J. Virol. 2007, 81, 11937–11945. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| dpi | PAstV-Challenged | Control |
|---|---|---|
| 2 d | Stomach bloat, gastric capillary congestion, small intestine bloat containing yellow foam liquid, mesenteric congestion | Mildly flatulence in individual pigs of the small intestine |
| 4 d | Stomach bloat, gastric capillary congestion, small intestine bloat containing yellow foam liquid, mesenteric congestion, cecum slightly flatulent | No lesions |
| 7 d | Yellow liquid contained in individual pigs of the small intestine | No lesions |
| 10 d | No lesions | No lesions |
| PAstV-Challenged | Control | |
|---|---|---|
| Duodenum | 0.42 ± 0.19 | 0 |
| Jejunum | 1.17 ± 0.24 | 0.73 ± 0.20 |
| Ileum | 0.83 ± 0.21 | 0.36 ± 0.15 |
| cecum | 0.17 ± 0.11 | 0 |
| colon | 0.08 ± 0.08 | 0.09 ± 0.09 |
| rectum | 0 | 0 |
| mesenteric lymph | 0.5 ± 0.15 | 0 |
| spleen | 0 | 0 |
| ung | 0 | 0 |
| kidney | 0 | 0 |
| stomach | 0.5 ± 0.15 | 0.27 ± 0.14 |
| dpi | Duodenum | Jejunum | Ileum | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Challenged | p | Control | Challenged | p | Control | Challenged | p | ||
| 2 | Villous height (μm) | 544.9 ± 15.88 | 447.7 ± 25.82 | 0.038 (*) | 461.6 ± 27.97 | 268.1 ± 27.97 | 0.0001 (***) | 344.9 ± 47.67 | 278.6 ± 47.67 | 0.2328 (NS) |
| Crypt depth (μm) | 147.1 ± 8.644 | 180.7 ± 13.21 | 0.0328 (*) | 147.0 ± 4.974 | 171.5 ± 9.240 | 0.0161 (*) | 121.6 ± 6.506 | 179.5 ± 22.36 | 0.002 (*) | |
| Ratio (μm/μm) | 3.704 ± 1.828 | 2.478 ± 1.955 | 3.140 ± 5.623 | 1.563 ± 0.0003 | 2.836 ± 7.327 | 1.558 ± 1.21 | ||||
| 4 | Villous height (μm) | 513.9 ± 68.77 | 274.1 ± 16.22 | 0.0003 (***) | 445.4 ± 47.62 | 260.9 ± 14.73 | 0.0003 (***) | 333.7 ± 11.59 | 264 ± 14.76 | 0.0001 (***) |
| Crypt depth (μm) | 168.3 ± 5.602 | 190.4 ± 11.12 | 0.1936 (NS) | 162.6 ± 7.897 | 161.3 ± 6.637 | 0.917 (NS) | 122.5 ± 6.575 | 145.6 ± 11.06 | 0.0649 (NS) | |
| Ratio (μm/μm) | 3.053 ± 12.276 | 1.440 ± 1.459 | 2.739 ± 6.03 | 1.617 ± 2.219 | 2.734 ± 1.763 | 1.813 ± 1.335 | ||||
| 7 | Villous height (μm) | 456.5 ± 32.36 | 420.1 ± 24.66 | 0.4361 (NS) | 442.3 ± 29.53 | 451.6 ± 40.22 | 0.8539 (NS) | 380 ± 22.29 | 310.2 ± 22.29 | 0.0728 (NS) |
| Crypt depth (μm) | 176.7 ± 5.904 | 182.0 ± 14.98 | 0.8042 (NS) | 162.6 ± 15.35 | 177.6 ± 11.22 | 0.4573 (NS) | 157.3 ± 6.014 | 138.8 ± 11.52 | 0.1380 (NS) | |
| Ratio (μm/μm) | 2.583 ± 5.481 | 2.308 ± 1.646 | 2.720 ± 1.924 | 2.543 ± 3.585 | 2.416 ± 3.706 | 2.235 ± 1.935 | ||||
| 10 | Villous height (μm) | 510.1 ± 9.831 | 485.4 ± 10.76 | 0.1181 (NS) | 312.6 ± 16.88 | 336 ± 17.5 | 0.3513 (NS) | 394.9 ± 8.937 | 367.8 ± 21.71 | 0.3243 (NS) |
| Crypt depth (μm) | 188.4 ± 14.14 | 211.7 ± 14.12 | 0.5219 (NS) | 194.3 ± 7.024 | 192.4 ± 8.213 | 0.8751 (NS) | 158 ± 16.27 | 162.6 ± 8.119 | 0.7906 (NS) | |
| Ratio (μm/μm) | 2.571 ± 0.695 | 2.293 ± 0.762 | 1.609 ± 2.403 | 1.746 ± 2.131 | 2.429 ± 0.549 | 2.262 ± 2.674 | ||||
| dpi | Number of Sections | Duodenum | Jejunum | Ileum | Cecum | Colon | Rectum | Mesenteric Lymph |
|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 6.67 ± 0.67 | 8.33 ± 2.33 | 9.00 ± 1.73 | 1.67 ± 0.33 | 2.00 ± 1.00 | 2.00 ± 1.00 | 7.33 ± 1.67 |
| 4 | 4 | 8.50 ± 1.66 | 6.25 ± 1.32 | 3.25 ± 0.75 | 1.25 ± 0.25 | 2.25 ± 1.32 | 1.13 ± 0.48 | 2.00 ± 1.35 |
| 7 | 3 | 5.00 ± 2.08 | 4.67 ± 1.33 | 2.33 ± 0.67 | 2.33 ± 0.88 | 1.67 ± 0.88 | 1.33 ± 0.67 | 1.33 ± 0.33 |
| 10 | 3 | 4.00 ± 2.65 | 2.00 ± 1.16 | 1.67 ± 1.20 | 1.00 ± 0.58 | 0.67 ± 0.33 | 0.67 ± 0.33 | 0.67 ± 0.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Q.; Wang, C.; Liu, H.; Wu, Q.; Liang, S.; Cen, M.; Dong, Q.; Wei, Y.; Chen, Y.; Ouyang, K.; et al. Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses 2019, 11, 1156. https://doi.org/10.3390/v11121156
Fang Q, Wang C, Liu H, Wu Q, Liang S, Cen M, Dong Q, Wei Y, Chen Y, Ouyang K, et al. Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses. 2019; 11(12):1156. https://doi.org/10.3390/v11121156
Chicago/Turabian StyleFang, Qingli, Cui Wang, Huan Liu, Qingping Wu, Siying Liang, Minli Cen, Qinting Dong, Yingyi Wei, Ying Chen, Kang Ouyang, and et al. 2019. "Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China" Viruses 11, no. 12: 1156. https://doi.org/10.3390/v11121156
APA StyleFang, Q., Wang, C., Liu, H., Wu, Q., Liang, S., Cen, M., Dong, Q., Wei, Y., Chen, Y., Ouyang, K., Wei, Z., & Huang, W. (2019). Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses, 11(12), 1156. https://doi.org/10.3390/v11121156