Preexisting Virus-Specific T Lymphocytes-Mediated Enhancement of Adenovirus Infections to Human Blood CD14+ Cells



Abstract
:
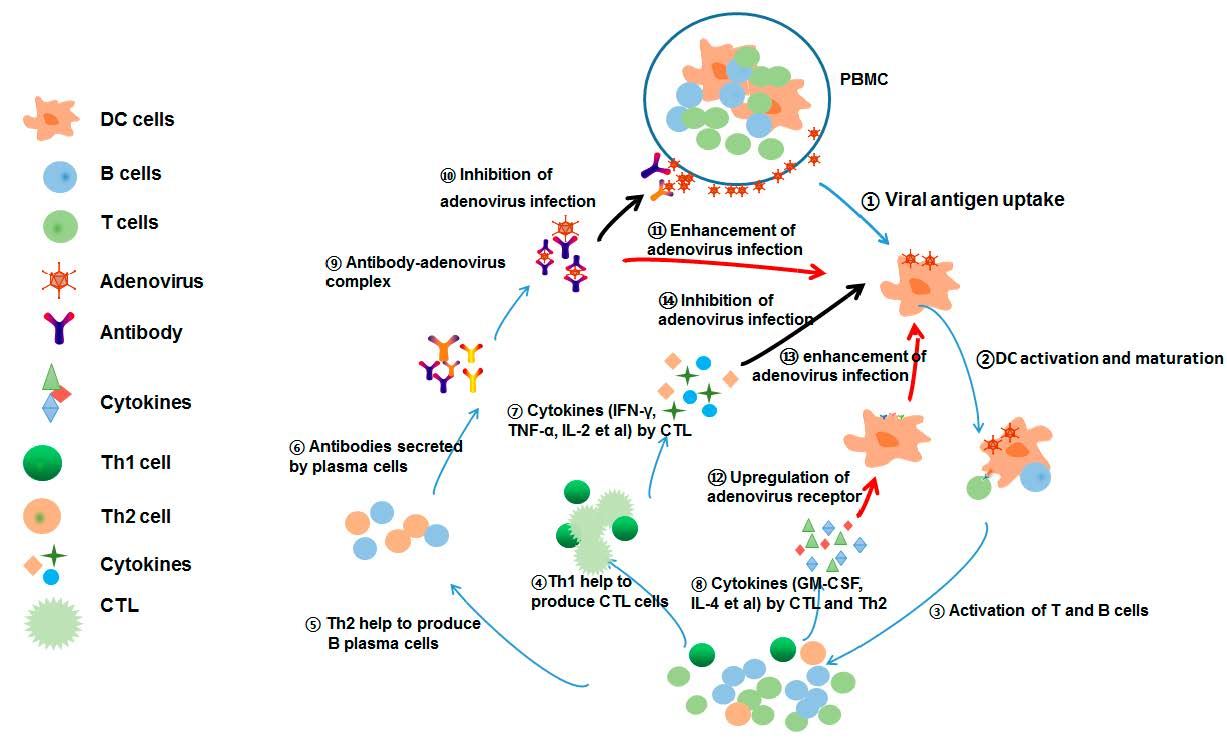{kind=link}
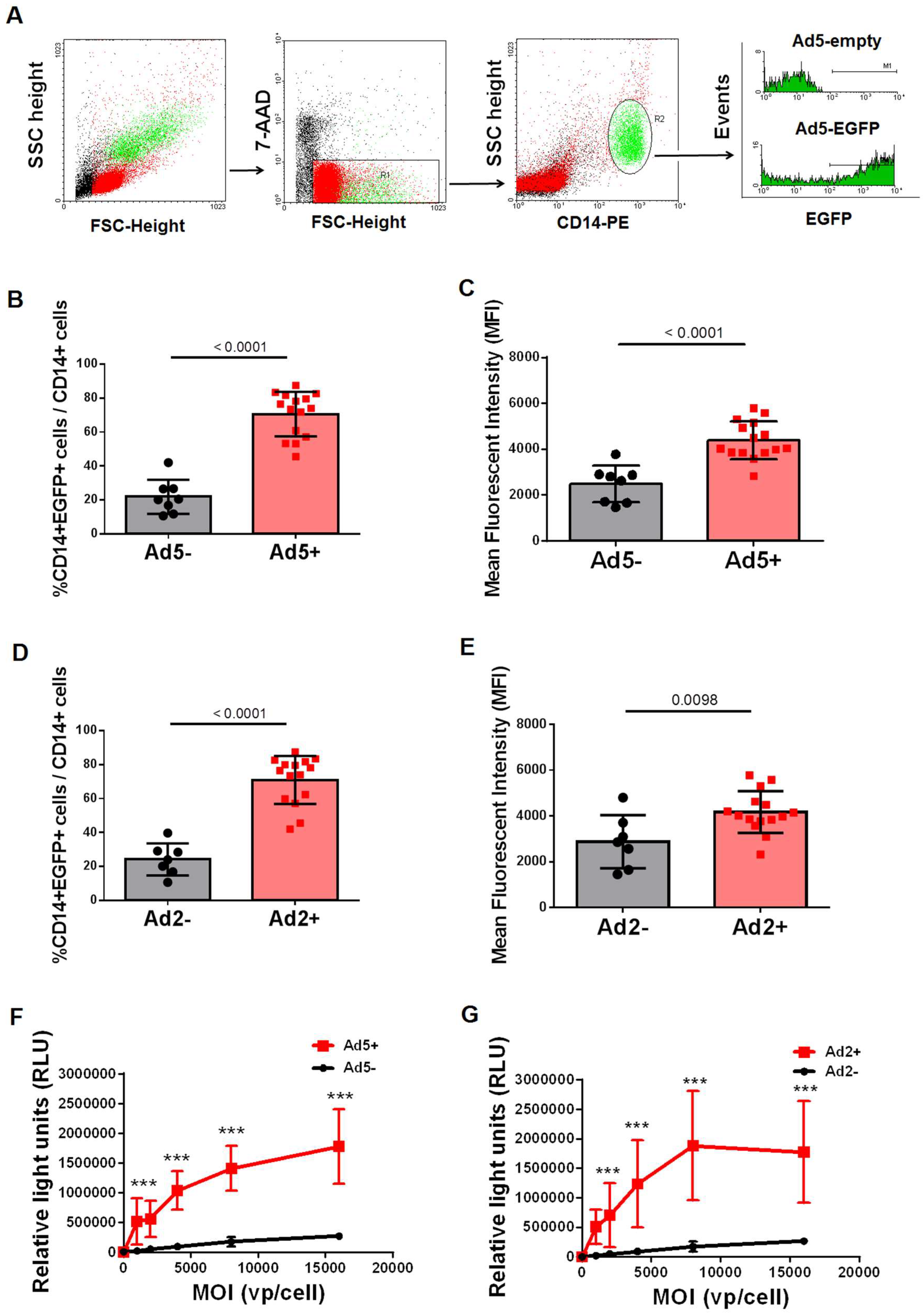{kind=link}
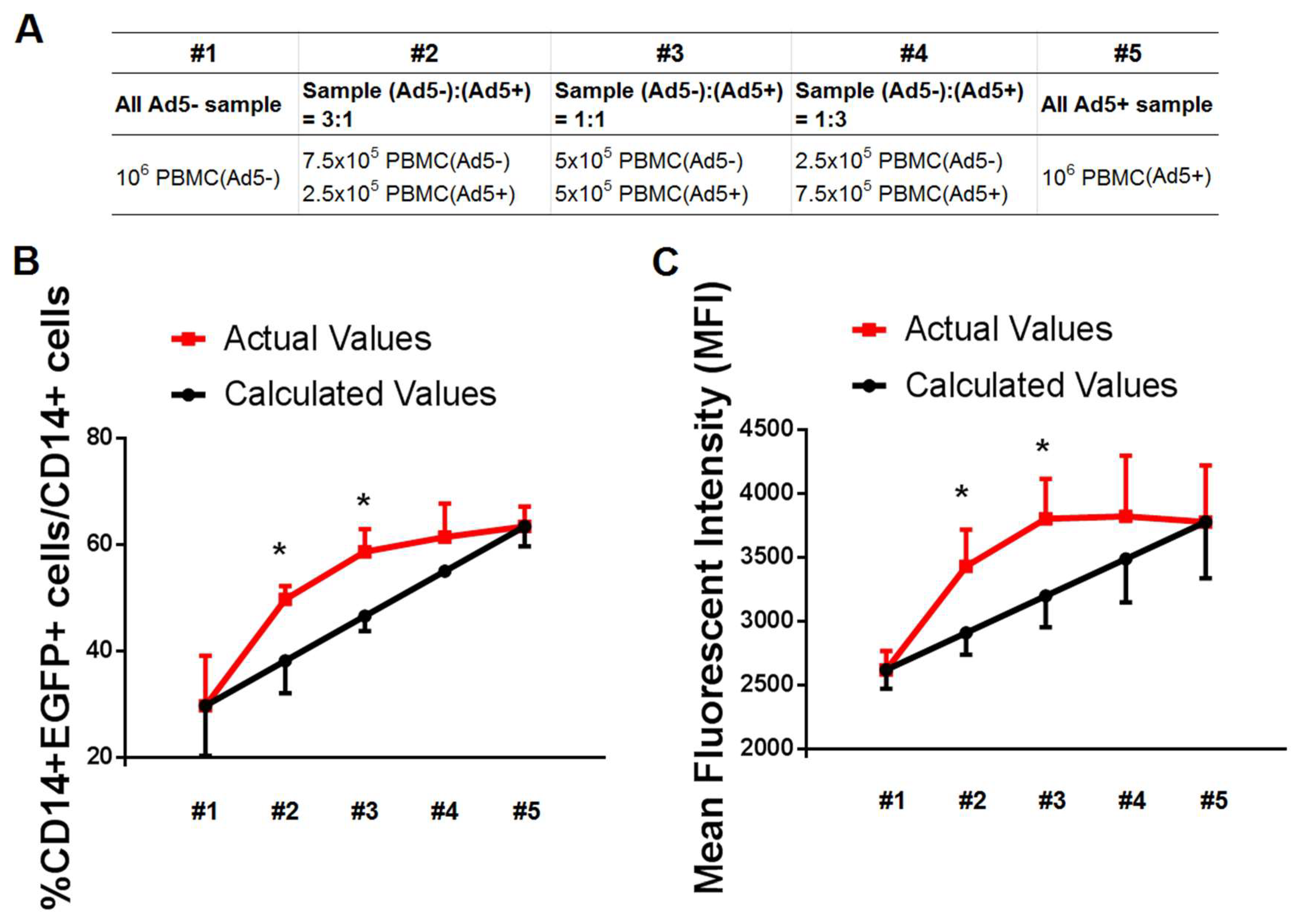{kind=link}
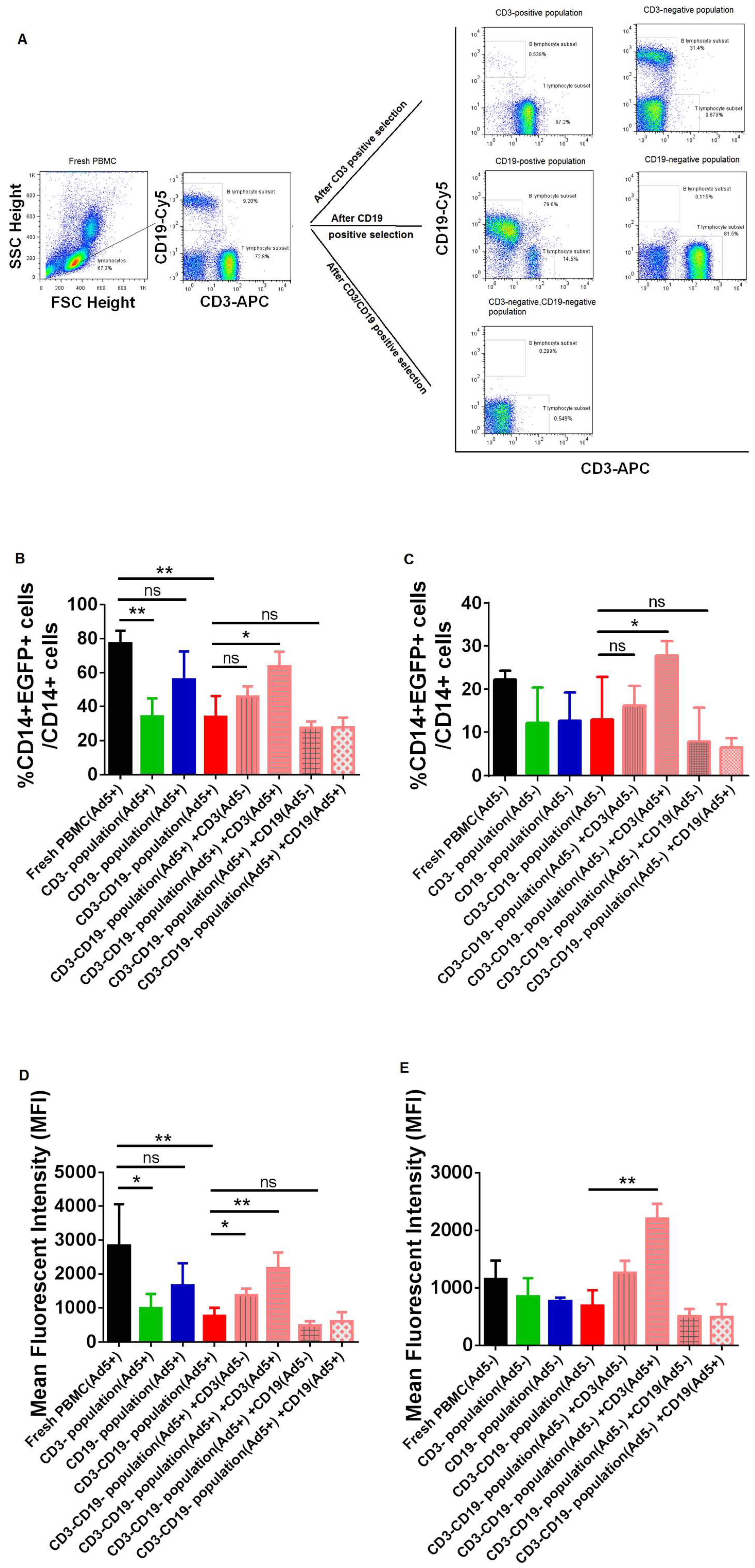{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Viruses, Cells and Ethic Statement
2.2. PBMC Isolation, Adenovirus Infection and Analysis for Infectivity
2.3. Sorting of Different Cell Subsets to Detect the Infectivity for Adenovirus
2.4. Quantitative PCR
2.5. Assay for SEAP-Based Ad Neutralizing Antibody
2.6. IFN-γ ELISPOT Assays
2.7. Incubation with Cytokines during Adenovirus Infection
2.8. Blockade with Anti-Cytokines Antibodies during Adenovirus Infection
2.9. Data Analysis
3. Results
3.1. CD14+ Monocytes from Ad-Seropositive Individuals Exhibited an Increased Susceptibility to Ad Infection
3.2. Secretory Factors by PBMC Samples of Ad-Seropositive Individuals Contributed to Promoting the Efficacy of Ad Infection
3.3. The Efficacy of Adenovirus Infection into CD14+ Monocytes were Significantly Affected by Removing or Adding CD3+ T Lymphocytes from PBMC Samples of Ad-Seropositive Individuals
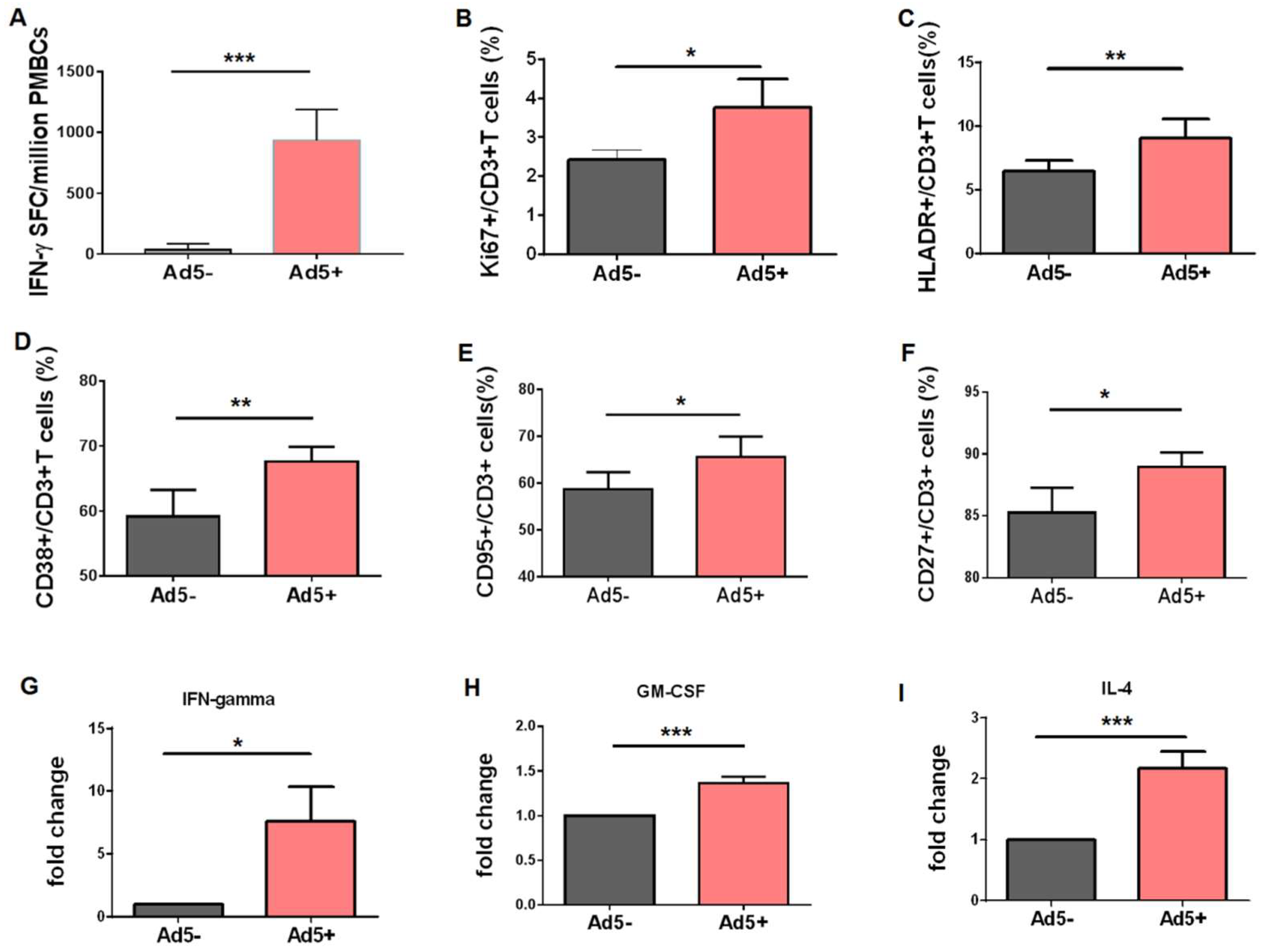
3.4. Preexisting Virus-Specific CD3+ T Lymphocytes were More Effectively Activated after Encountering Adenovirus Stimulus
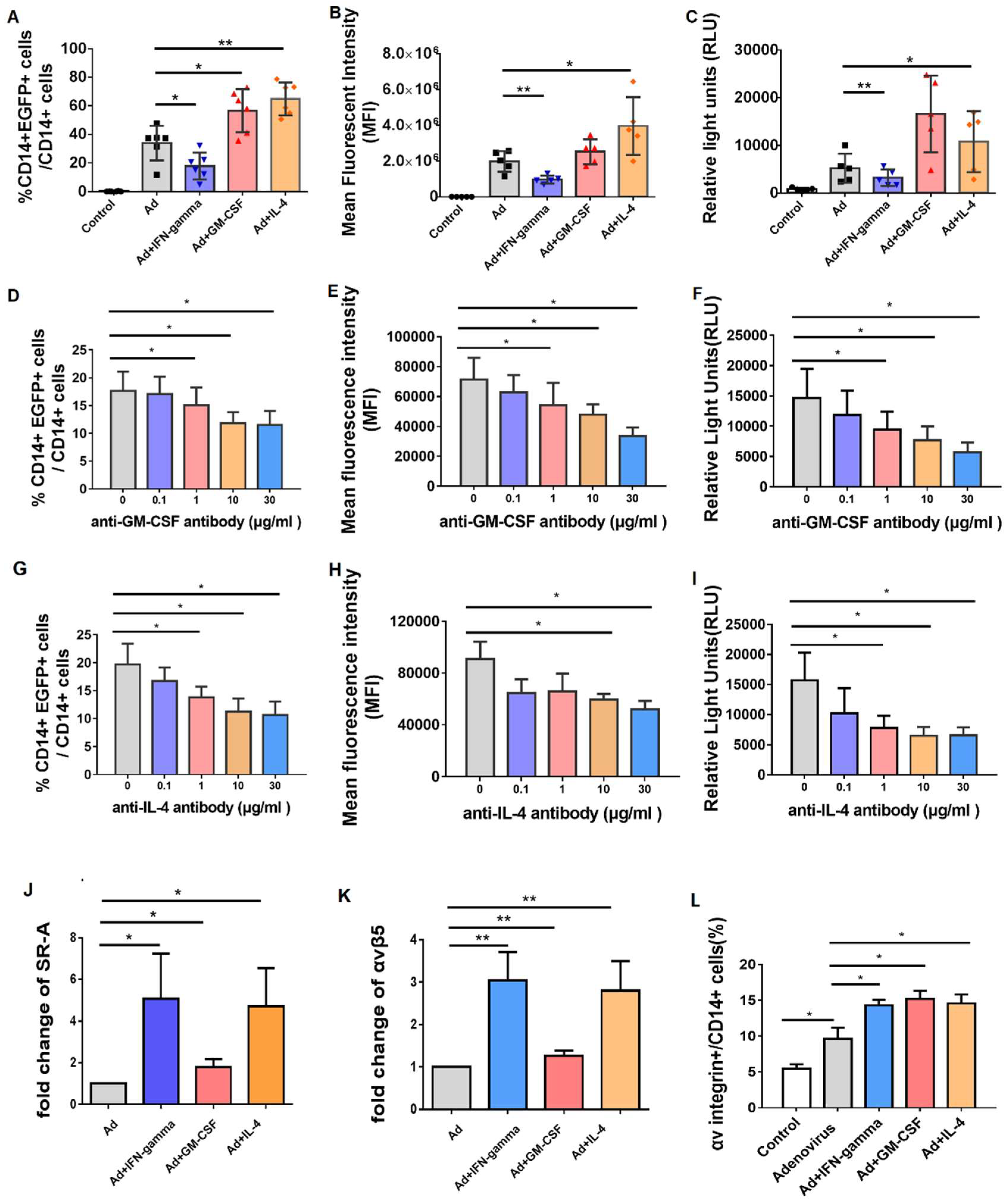
3.5. Incubation of GM-CSF and IL-4 Promoted the Efficacy of Adenovirus Infection by Upregulating SR-A and Integrin Receptor
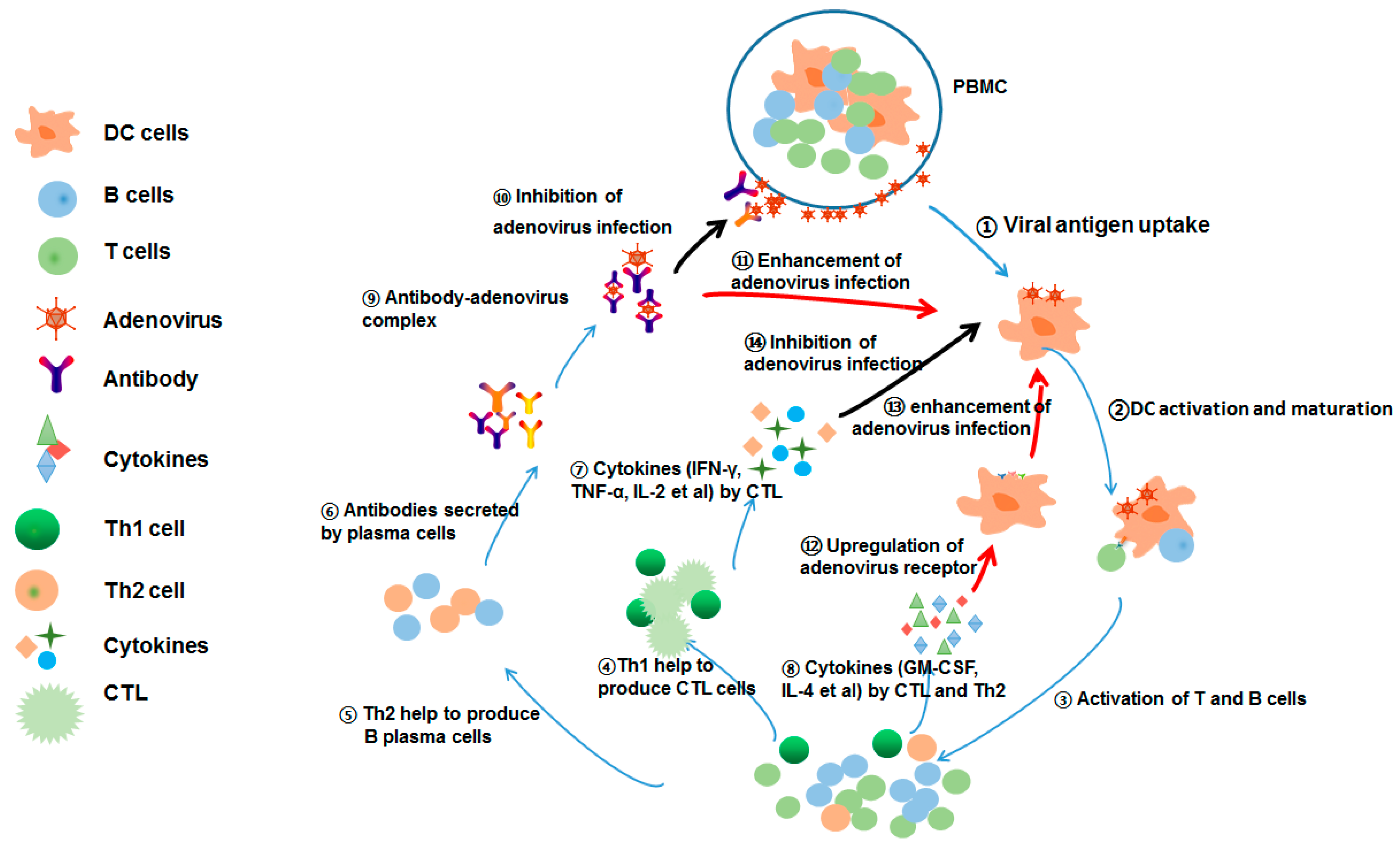
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McEwan, W.A.; Tam, J.C.; Watkinson, R.E.; Bidgood, S.R.; Mallery, D.L.; James, L.C. Intracellular antibody-bound pathogens stimulate immune signaling via the fc receptor trim21. Nat. Immunol. 2013, 14, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Barry, M.; Bleackley, R.C. Cytotoxic t lymphocytes: All roads lead to death. Nat. Rev. Immunol. 2002, 2, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Pan, E.; Feng, F.; Li, P.; Yang, Q.; Ma, X.; Wu, C.; Zhao, J.; Yan, H.; Chen, R.; Chen, L.; et al. Immune protection of siv challenge by pd-1 blockade during vaccination in rhesus monkeys. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Hensley, L.; McKnight, K.L.; Hu, F.; Madden, V.; Ping, L.; Jeong, S.H.; Walker, C.; Lanford, R.E.; Lemon, S.M. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature 2013, 496, 367–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Q.T.; Ren, X.; Zhang, R.; Lee, I.H.; Hurley, J.H. Hiv-1 nef hijacks clathrin coats by stabilizing ap-1:Arf1 polygons. Science 2015, 350, aac5137. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Goraya, M.U.; Maarouf, M.; Huang, S.; Chen, J.L. Host immune response to influenza a virus infection. Front. Immunol. 2018, 9, 320. [Google Scholar] [CrossRef] [PubMed]
- Sebina, I.; Pepper, M. Humoral immune responses to infection: Common mechanisms and unique strategies to combat pathogen immune evasion tactics. Curr. Opin. Immunol. 2018, 51, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Pelegrin, M.; Naranjo-Gomez, M.; Piechaczyk, M. Antiviral monoclonal antibodies: Can they be more than simple neutralizing agents? Trends. Microbiol. 2015, 23, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.; Foo, S.S.; Bruzzone, R.; Dinh, L.V.; King, N.J.; Mahalingam, S. Fc receptors in antibody-dependent enhancement of viral infections. Immunol. Rev. 2015, 268, 340–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khandia, R.; Munjal, A.; Dhama, K.; Karthik, K.; Tiwari, R.; Malik, Y.S.; Singh, R.K.; Chaicumpa, W. Modulation of dengue/zika virus pathogenicity by antibody-dependent enhancement and strategies to protect against enhancement in zika virus infection. Front. Immunol. 2018, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.; Bousema, T.; Sauerwein, R.; Drakeley, C. Two-faced immunity? The evidence for antibody enhancement of malaria transmission. Trends Parasitol. 2019, 35, 140–153. [Google Scholar] [CrossRef]
- Halstead, S.B.; Mahalingam, S.; Marovich, M.A.; Ubol, S.; Mosser, D.M. Intrinsic antibody-dependent enhancement of microbial infection in macrophages: Disease regulation by immune complexes. Lancet Infect. Dis. 2010, 10, 712–722. [Google Scholar] [CrossRef]
- Nwanegbo, E.; Vardas, E.; Gao, W.; Whittle, H.; Sun, H.; Rowe, D.; Robbins, P.D.; Gambotto, A. Prevalence of neutralizing antibodies to adenoviral serotypes 5 and 35 in the adult populations of the gambia, south africa, and the united states. Clin. Diagn. Lab. Immunol. 2004, 11, 351–357. [Google Scholar] [CrossRef]
- Dormond, E.; Perrier, M.; Kamen, A. From the first to the third generation adenoviral vector: What parameters are governing the production yield? Biotechnol. Adv. 2009, 27, 133–144. [Google Scholar] [CrossRef]
- Feng, Y.; Li, C.; Hu, P.; Wang, Q.; Zheng, X.; Zhao, Y.; Shi, Y.; Yang, S.; Yi, C.; Feng, Y.; et al. An adenovirus serotype 2-vectored ebolavirus vaccine generates robust antibody and cell-mediated immune responses in mice and rhesus macaques. Emerg. Microbes Infect. 2018, 7, 101. [Google Scholar] [CrossRef]
- Sun, C.; Chen, Z.; Tang, X.; Zhang, Y.; Feng, L.; Du, Y.; Xiao, L.; Liu, L.; Zhu, W.; Chen, L.; et al. Mucosal priming with a replicating-vaccinia virus-based vaccine elicits protective immunity to simian immunodeficiency virus challenge in rhesus monkeys. J. Virol. 2013, 87, 5669–5677. [Google Scholar] [CrossRef]
- Kim, E.H.; Han, G.Y.; Nguyen, H. An adenovirus-vectored influenza vaccine induces durable cross-protective hemagglutinin stalk antibody responses in mice. Viruses. 2017, 9, 234. [Google Scholar] [CrossRef]
- Bressy, C.; Hastie, E.; Grdzelishvili, V.Z. Combining oncolytic virotherapy with p53 tumor suppressor gene therapy. Mol. Ther. Oncolytics 2017, 5, 20–40. [Google Scholar] [CrossRef]
- Zhu, F.C.; Wurie, A.H.; Hou, L.H.; Liang, Q.; Li, Y.H.; Russell, J.B.; Wu, S.P.; Li, J.X.; Hu, Y.M.; Guo, Q.; et al. Safety and immunogenicity of a recombinant adenovirus type-5 vector-based ebola vaccine in healthy adults in sierra leone: A single-centre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 389, 621–628. [Google Scholar] [CrossRef]
- Wong, G.; Mendoza, E.J.; Plummer, F.A.; Gao, G.F.; Kobinger, G.P.; Qiu, X. From bench to almost bedside: The long road to a licensed ebola virus vaccine. Expert Opin. Biol. Ther. 2018, 18, 159–173. [Google Scholar] [CrossRef]
- Echavarria, M. Adenoviruses in immunocompromised hosts. Clin. Microbiol. Rev. 2008, 21, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Thorner, A.R.; Vogels, R.; Kaspers, J.; Weverling, G.J.; Holterman, L.; Lemckert, A.A.; Dilraj, A.; McNally, L.M.; Jeena, P.M.; Jepsen, S.; et al. Age dependence of adenovirus-specific neutralizing antibody titers in individuals from sub-saharan africa. J. Clin. Microbiol. 2006, 44, 3781–3783. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Wang, Z.; Dong, J.; Wang, C.; Gu, L.; Sun, C.; Kong, W.; Yu, X. A serological survey of human adenovirus serotype 2 and 5 circulating pediatric populations in changchun, china, 2011. Virol. J. 2012, 9, 287. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Feng, L.; Zhang, Y.; Xiao, L.; Pan, W.; Li, C.; Zhang, L.; Chen, L. Circumventing antivector immunity by using adenovirus-infected blood cells for repeated application of adenovirus-vectored vaccines: Proof of concept in rhesus macaques. J. Virol. 2012, 86, 11031–11042. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Feng, F.; Pan, E.; Fan, X.; Yang, Q.; Guan, M.; Chen, L.; Sun, C. Scavenger receptor-mediated ad5 entry and acldl accumulation in monocytes/macrophages synergistically trigger innate responses against viral infection. Virology. 2018, 519, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhang, L.; Zhang, M.; Liu, Y.; Zhong, M.; Ma, X.; Chen, L. Induction of balance and breadth in the immune response is beneficial for the control of sivmac239 replication in rhesus monkeys. J. Infect. 2010, 60, 371–381. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, Y.; Feng, L.; Pan, W.; Zhang, M.; Hong, Z.; Ma, X.; Chen, X.; Chen, L. Epidemiology of adenovirus type 5 neutralizing antibodies in healthy people and aids patients in guangzhou, southern china. Vaccine 2011, 29, 3837–3841. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, C.; Li, P.; Wu, T.; Zhou, H.; Yang, D.; Liu, Y.; Ma, X.; Song, Z.; Nian, Q.; et al. Vaccine engineering with dual-functional mineral shell: A promising strategy to overcome preexisting immunity. Adv. Mater. 2016, 28, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, C.H.; Roberts, A.I.; Das, J.; Xu, G.; Ren, G.; Zhang, Y.; Zhang, L.; Yuan, Z.R.; Tan, H.S.; et al. Granulocyte-macrophage colony-stimulating factor (gm-csf) and t-cell responses: What we do and don’t know. Cell Res. 2006, 16, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Sallerfors, B. Endogenous production and peripheral blood levels of granulocyte-macrophage (gm-) and granulocyte (g-) colony-stimulating factors. Leuk. Lymphoma 1994, 13, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Puzanov, I.; Kelley, M.C. Talimogene laherparepvec (t-vec) for the treatment of advanced melanoma. Immunotherapy 2015, 7, 611–619. [Google Scholar] [CrossRef]
- Senzer, N.N.; Kaufman, H.L.; Amatruda, T.; Nemunaitis, M.; Reid, T.; Daniels, G.; Gonzalez, R.; Glaspy, J.; Whitman, E.; Harrington, K.; et al. Phase ii clinical trial of a granulocyte-macrophage colony-stimulating factor-encoding, second-generation oncolytic herpesvirus in patients with unresectable metastatic melanoma. J. Clin. Oncol. 2009, 27, 5763–5771. [Google Scholar] [CrossRef]
- Menotti, L.; Avitabile, E.; Gatta, V.; Malatesta, P.; Petrovic, B.; Campadelli-Fiume, G. Hsv as a platform for the generation of retargeted, armed, and reporter-expressing oncolytic viruses. Viruses. 2018, 10, 352. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Endo, R.I.; Nemerow, G.R. Upregulation of integrins alpha v beta 3 and alpha v beta 5 on human monocytes and t lymphocytes facilitates adenovirus-mediated gene delivery. J. Virol. 1995, 69, 2257–2263. [Google Scholar] [PubMed]
- Schaeffer, E.; Flacher, V.; Papageorgiou, V.; Decossas, M.; Fauny, J.D.; Kramer, M.; Mueller, C.G. Dermal cd14(+) dendritic cell and macrophage infection by dengue virus is stimulated by interleukin-4. J. Investig. Dermatol. 2015, 135, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.A.; Dobmeyer, J.M.; Dobmeyer, T.S.; Pape, M.; Ottmann, O.G.; Helm, E.B.; Hoelzer, D.; Rossol, R. Demonstration of the th1 to th2 cytokine shift during the course of hiv-1 infection using cytoplasmic cytokine detection on single cell level by flow cytometry. Aids 1997, 11, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, T.; Hooks, J.J.; Dougherty, S.F.; Oppenheim, J.J. Interleukin 2-mediated immune interferon (ifn-gamma) production by human t cells and t cell subsets. J. Immunol. 1983, 130, 1784–1789. [Google Scholar]
- Gresser, I. Biologic effects of interferons. J. Investig. Dermatol. 1990, 95, 66s–71s. [Google Scholar] [CrossRef]
- Mistchenko, A.S.; Diez, R.A.; Falcoff, R. Inhibitory effect of interferon-gamma on adenovirus replication and late transcription. Biochem. Pharmacol. 1989, 38, 1971–1978. [Google Scholar] [CrossRef]
- Mistchenko, A.S.; Falcoff, R. Recombinant human interferon-gamma inhibits adenovirus multiplication in vitro. J. Gen. Virol. 1987, 68 Pt 3, 941–944. [Google Scholar] [CrossRef]
- Liu, Q.; Hamblin, M.R. Macrophage-targeted photodynamic therapy: Scavenger receptor expression and activation state. Int. J. Immunopathol. Pharmacol. 2005, 18, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Grewal, T.; Priceputu, E.; Davignon, J.; Bernier, L. Identification of a gamma-interferon-responsive element in the promoter of the human macrophage scavenger receptor a gene. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.K.; Dmitriev, I.P.; Kashentseva, E.A.; Raes, G.; Li, L.; Kim, S.W.; Lu, Z.H.; Arbeit, J.M.; Fleming, T.P.; Kaliberov, S.A.; et al. Development of an adenovirus vector vaccine platform for targeting dendritic cells. Cancer Gene Ther. 2018, 25, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; Gao, W.; Alber, S.; Trichel, A.; Murphey-Corb, M.; Watkins, S.C.; Gambotto, A.; Barratt-Boyes, S.M. Adenovirus-transduced dendritic cells injected into skin or lymph node prime potent simian immunodeficiency virus-specific t cell immunity in monkeys. J. Immunol. 2003, 171, 6875–6882. [Google Scholar] [CrossRef] [PubMed]
- De Rose, R.; Fernandez, C.S.; Loh, L.; Peut, V.; Mason, R.D.; Alcantara, S.; Reece, J.; Kent, S.J. Delivery of immunotherapy with peptide-pulsed blood in macaques. Virology 2008, 378, 201–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, M.; Onion, D.; Green, N.K.; Aslan, K.; Rajaratnam, R.; Bazan-Peregrino, M.; Phipps, S.; Hale, S.; Mautner, V.; Seymour, L.W.; et al. Adenovirus type 5 interactions with human blood cells may compromise systemic delivery. Mol. Ther. 2006, 14, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Mayne, G.C.; Borowicz, R.A.; Greeneklee, K.V.; Finlay-Jones, J.J.; Williams, K.A.; Hart, P.H. Centrifugation facilitates transduction of green fluorescent protein in human monocytes and macrophages by adenovirus at low multiplicity of infection. J. Immunol. Methods 2003, 278, 45–56. [Google Scholar] [CrossRef]
- Nishimura, N.; Nishioka, Y.; Shinohara, T.; Sone, S. Enhanced efficiency by centrifugal manipulation of adenovirus-mediated interleukin 12 gene transduction into human monocyte-derived dendritic cells. Hum. Gene Ther. 2001, 12, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Son, D.; Choi, T.; Yeo, H.; Kim, J.; Han, K. The effect of centrifugation condition on mature adipocytes and adipose stem cell viability. Ann. Plast. Surg. 2014, 72, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Tavakolinejad, A.; Rabbani, M.; Janmaleki, M. Effects of hypergravity on adipose-derived stem cell morphology, mechanical property and proliferation. Biochem. Biophys. Res. Commun. 2015, 464, 473–479. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, F.; Zhao, J.; Li, P.; Li, R.; Chen, L.; Sun, C. Preexisting Virus-Specific T Lymphocytes-Mediated Enhancement of Adenovirus Infections to Human Blood CD14+ Cells. Viruses 2019, 11, 154. https://doi.org/10.3390/v11020154
Feng F, Zhao J, Li P, Li R, Chen L, Sun C. Preexisting Virus-Specific T Lymphocytes-Mediated Enhancement of Adenovirus Infections to Human Blood CD14+ Cells. Viruses. 2019; 11(2):154. https://doi.org/10.3390/v11020154
Chicago/Turabian StyleFeng, Fengling, Jin Zhao, Pingchao Li, Ruiting Li, Ling Chen, and Caijun Sun. 2019. "Preexisting Virus-Specific T Lymphocytes-Mediated Enhancement of Adenovirus Infections to Human Blood CD14+ Cells" Viruses 11, no. 2: 154. https://doi.org/10.3390/v11020154
APA StyleFeng, F., Zhao, J., Li, P., Li, R., Chen, L., & Sun, C. (2019). Preexisting Virus-Specific T Lymphocytes-Mediated Enhancement of Adenovirus Infections to Human Blood CD14+ Cells. Viruses, 11(2), 154. https://doi.org/10.3390/v11020154