Enhanced Ability of Oligomeric Nanobodies Targeting MERS Coronavirus Receptor-Binding Domain

 ,
,
Abstract
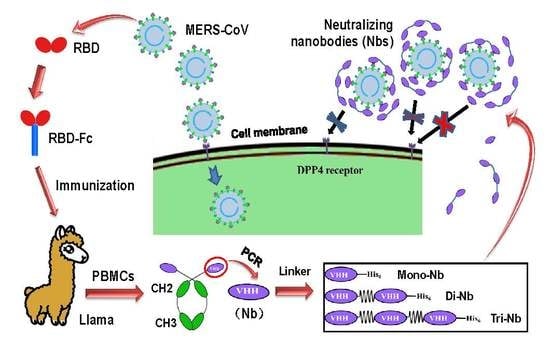
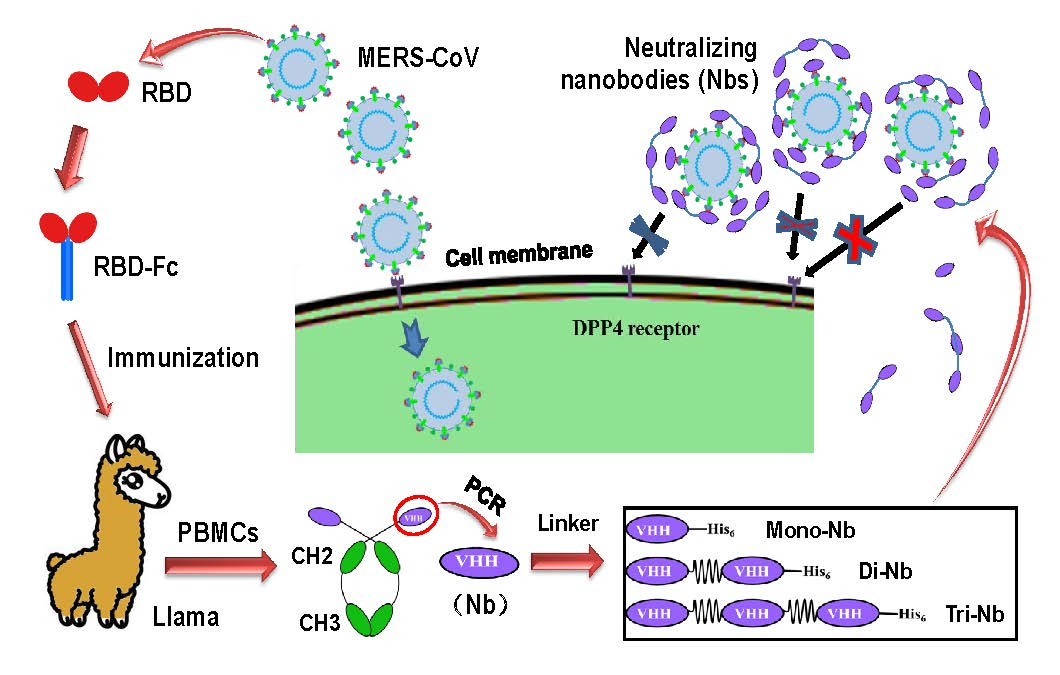
:
1. Introduction
2. Materials and Methods
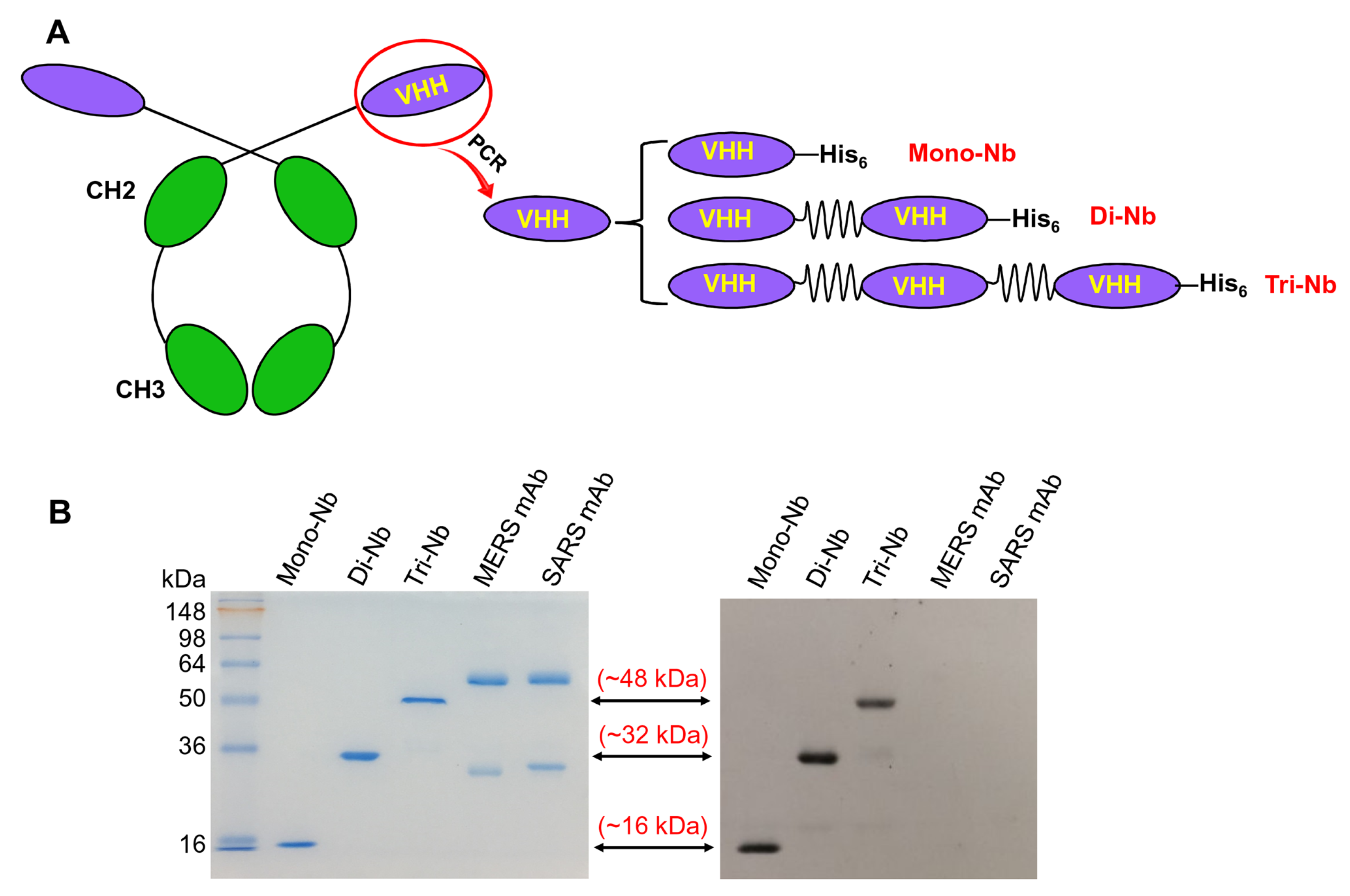
2.1. Construction and Expression of MERS-CoV RBD-Specific Dimeric and Trimeric Nbs
2.2. SDS-PAGE and Western Blot
2.3. ELISA
2.4. Surface Plasmon Resonance (SPR)
2.5. Flow Cytometry
2.6. MERS-CoV Micro-Neutralization Assay
2.7. MERS Pseudovirus Neutralization Assay
2.8. Detection of Nb Stability
2.9. Statistical Analysis
3. Results
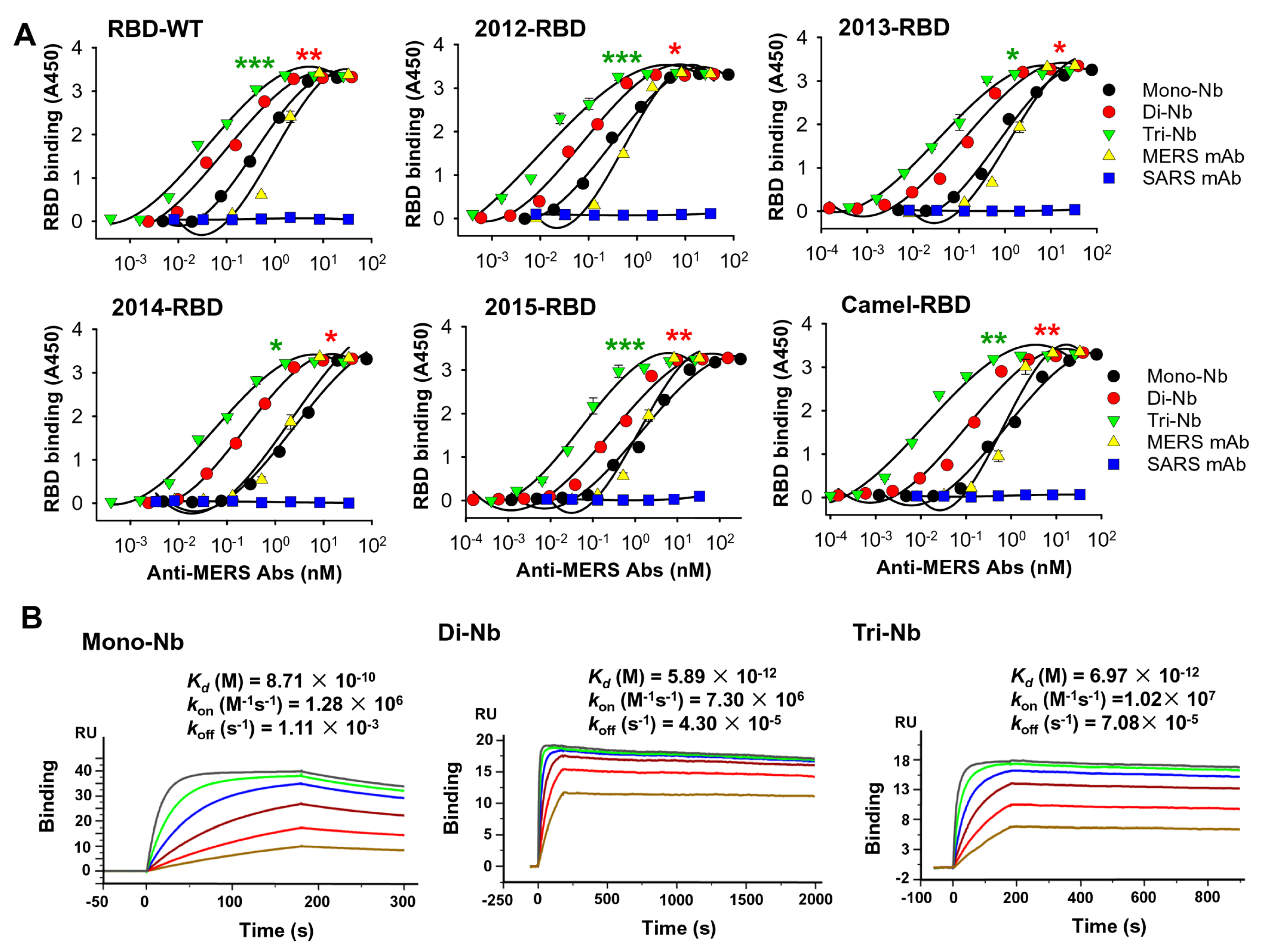
3.1. Construction and Characterization of MERS-CoV RBD-Targeting Dimeric and Trimeric Nbs
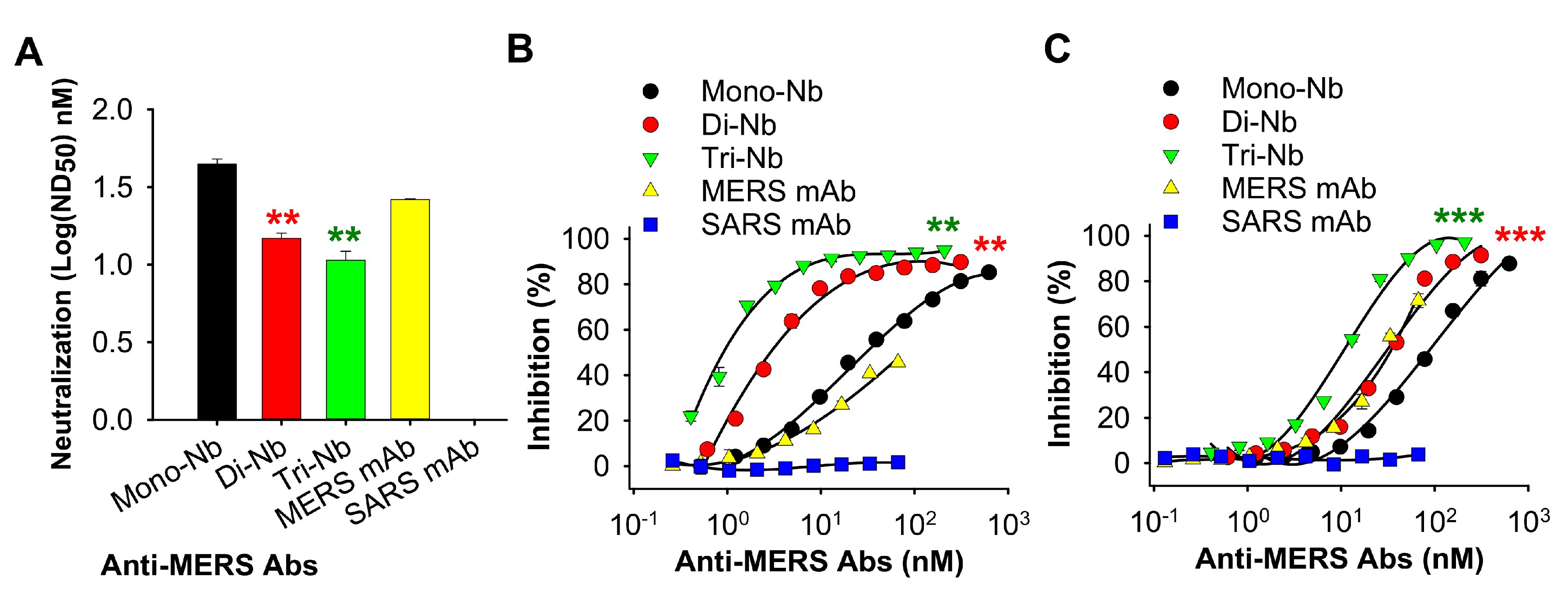
3.2. MERS-CoV RBD-Targeting Dimeric and Trimeric Nbs Exhibited Superior Binding toward MERS-CoV RBD, Neutralization of MERS-CoV Infection, and Inhibition of RBD–hDPP4 Binding
3.3. MERS-CoV RBD-Targeting Nbs Maintain Strong pH, Protease, Chemical, and Thermal Stability
3.4. MERS-CoV RBD-Targeting Dimeric and Trimeric Nbs Had Significantly Elevated Cross-Neutralizing Activity Against Multiple Heterologous MERS-CoV Isolates
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Zaki, A.M.; Van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, L.; Liu, C.; Wang, L.; Ma, C.; Tang, J.; Baric, R.S.; Jiang, S.; Li, F. Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus. Proc. Natl. Acad. Sci. USA 2014, 111, 12516–12521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munster, V.J.; Adney, D.R.; Van Doremalen, N.; Brown, V.R.; Miazgowicz, K.L.; Milne-Price, S.; Bushmaker, T.; Rosenke, R.; Scott, D.; Hawkinson, A.; et al. Replication and shedding of MERS-CoV in Jamaican fruit bats (Artibeus jamaicensis). Sci. Rep. 2016, 6, 21878. [Google Scholar] [CrossRef] [Green Version]
- Anthony, S.J.; Gilardi, K.; Menachery, V.D.; Goldstein, T.; Ssebide, B.; Mbabazi, R.; Navarrete-Macias, I.; Liang, E.; Wells, H.; Hicks, A.; et al. Further evidence for bats as the evolutionary source of Middle East respiratory syndrome coronavirus. MBio 2017, 8, e00373-17. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Wong, A.C.P.; Lau, T.C.K.; Woo, P.C.Y. Molecular evolution of MERS Coronavirus: Dromedaries as a recent intermediate host or long-time animal reservoir? Int. J. Mol. Sci. 2017, 18, 2138. [Google Scholar] [CrossRef] [PubMed]
- van Doremalen, N.; Hijazeen, Z.S.; Holloway, P.; Al, O.B.; McDowell, C.; Adney, D.; Talafha, H.A.; Guitian, J.; Steel, J.; Amarin, N.; et al. High prevalence of Middle East respiratory coronavirus in young dromedary camels in Jordan. Vector Borne Zoonotic Dis. 2017, 17, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Madani, T.A.; Azhar, E.I.; Hashem, A.M. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 371, 1360. [Google Scholar] [PubMed]
- Hemida, M.G.; Elmoslemany, A.; Al-Hizab, F.; Alnaeem, A.; Almathen, F.; Faye, B.; Chu, D.K.; Perera, R.A.; Peiris, M. Dromedary camels and the transmission of Middle East respiratory syndrome coronavirus (MERS-CoV). Transbound. Emerg. Dis. 2015, 64, 344–353. [Google Scholar] [CrossRef]
- Hunter, J.C.; Nguyen, D.; Aden, B.; Al, B.Z.; Al, D.W.; Abu, E.K.; Khudair, A.; Al, M.M.; El, S.F.; Imambaccus, H.; et al. Transmission of Middle East respiratory syndrome coronavirus infections in healthcare settings, Abu Dhabi. Emerg. Infect. Dis. 2016, 22, 647–656. [Google Scholar] [CrossRef]
- Alhakeem, R.F.; Midgley, C.M.; Assiri, A.M.; Alessa, M.; Al, H.H.; Saeed, A.B.; Almasri, M.M.; Lu, X.; Abedi, G.R.; Abdalla, O.; et al. Exposures among MERS case-patients, Saudi Arabia, January-February 2016. Emerg. Infect. Dis. 2016, 22, 2020–2022. [Google Scholar] [CrossRef]
- Chen, X.; Chughtai, A.A.; Dyda, A.; MacIntyre, C.R. Comparative epidemiology of Middle East respiratory syndrome coronavirus (MERS-CoV) in Saudi Arabia and South Korea. Emerg. Microbes Infect. 2017, 6, e51. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y. An outbreak of Middle East respiratory syndrome coronavirus infection in South Korea, 2015. Yonsei Med. J. 2015, 56, 1174–1176. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Wong, N.S. Probable transmission chains of Middle East respiratory syndrome coronavirus and the multiple generations of secondary infection in South Korea. Int. J. Infect. Dis. 2015, 38, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Jiang, S.; Du, L. Prospects for a MERS-CoV spike vaccine. Expert Rev. Vaccines 2018, 17, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Tai, W.; Zhou, Y.; Jiang, S. Vaccines for the prevention against the threat of MERS-CoV. Expert Rev. Vaccines 2016, 15, 1123–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Hu, Y.; Wang, Q.; Qi, J.; Gao, F.; Li, Y.; Zhang, Y.; Zhang, W.; Yuan, Y.; Bao, J.; et al. Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26. Nature 2013, 500, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Muller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Q.; Zhu, Y.; Chan, K.H.; Qin, L.; Li, Y.; Wang, Q.; Chan, J.F.; Du, L.; Yu, F.; et al. Structure-based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor. Nat. Commun. 2014, 5, 3067. [Google Scholar] [CrossRef] [Green Version]
- Li, F. Receptor recognition mechanisms of coronaviruses: A decade of structural studies. J. Virol. 2015, 89, 1954–1964. [Google Scholar] [CrossRef]
- Chen, Y.; Rajashankar, K.R.; Yang, Y.; Agnihothram, S.S.; Liu, C.; Lin, Y.L.; Baric, R.S.; Li, F. Crystal structure of the receptor-binding domain from newly emerged Middle East respiratory syndrome coronavirus. J. Virol. 2013, 87, 10777–10783. [Google Scholar] [CrossRef]
- Ma, C.; Wang, L.; Tao, X.; Zhang, N.; Yang, Y.; Tseng, C.T.; Li, F.; Zhou, Y.; Jiang, S.; Du, L. Searching for an ideal vaccine candidate among different MERS coronavirus receptor-binding fragments—The importance of immunofocusing in subunit vaccine design. Vaccine 2014, 32, 6170–6176. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Tang, J.; Lu, L.; Jiang, S.; Du, L. Receptor-binding domain-based subunit vaccines against MERS-CoV. Virus Res. 2015, 202, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Tai, W.; Yang, Y.; Zhao, G.; Zhu, Q.; Sun, S.; Liu, C.; Tao, X.; Tseng, C.K.; Perlman, S.; et al. Introduction of neutralizing immunogenicity index to the rational design of MERS coronavirus subunit vaccines. Nat. Commun. 2016, 7, 13473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, W.; Wang, Y.; Fett, C.A.; Zhao, G.; Li, F.; Perlman, S.; Jiang, S.; Zhou, Y.; Du, L. Recombinant receptor-binding domains of multiple Middle East respiratory syndrome coronaviruses (MERS-CoVs) induce cross-neutralizing antibodies against divergent human and camel MERS-CoVs and antibody escape mutants. J. Virol. 2017, 91, e01651-16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Channappanavar, R.; Ma, C.; Wang, L.; Tang, J.; Garron, T.; Tao, X.; Tasneem, S.; Lu, L.; Tseng, C.T.; et al. Identification of an ideal adjuvant for receptor-binding domain-based subunit vaccines against Middle East respiratory syndrome coronavirus. Cell. Mol. Immunol. 2016, 13, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Yang, Y.; Zhou, Y.; Lu, L.; Li, F.; Jiang, S. MERS-CoV spike protein: A key target for antivirals. Expert Opin. Ther. Targets 2017, 21, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Sun, S.; Xiao, H.; Feng, J.; Guo, Y.; Tai, W.; Wang, Y.; Du, L.; Zhao, G.; Zhou, Y. Single-dose treatment with a humanized neutralizing antibody affords full protection of a human transgenic mouse model from lethal Middle East respiratory syndrome (MERS)-coronavirus infection. Antiviral Res. 2016, 132, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Zhao, G.; Yang, Y.; Qiu, H.; Wang, L.; Kou, Z.; Tao, X.; Yu, H.; Sun, S.; Tseng, C.T.; et al. A conformation-dependent neutralizing monoclonal antibody specifically targeting receptor-binding domain in Middle East respiratory syndrome coronavirus spike protein. J. Virol. 2014, 88, 7045–7053. [Google Scholar] [CrossRef]
- Corti, D.; Zhao, J.; Pedotti, M.; Simonelli, L.; Agnihothram, S.; Fett, C.; Fernandez-Rodriguez, B.; Foglierini, M.; Agatic, G.; Vanzetta, F.; et al. Prophylactic and postexposure efficacy of a potent human monoclonal antibody against MERS coronavirus. Proc. Natl Acad. Sci. USA 2015, 112, 10473–10478. [Google Scholar] [CrossRef]
- Ying, T.; Prabakaran, P.; Du, L.; Shi, W.; Feng, Y.; Wang, Y.; Wang, L.; Li, W.; Jiang, S.; Dimitrov, D.S.; et al. Junctional and allele-specific residues are critical for MERS-CoV neutralization by an exceptionally potent germline-like antibody. Nat. Commun. 2015, 6, 8223. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Wang, N.; Zuo, T.; Shi, X.; Poon, K.M.; Wu, Y.; Gao, F.; Li, D.; Wang, R.; Guo, J.; et al. Potent neutralization of MERS-CoV by human neutralizing monoclonal antibodies to the viral spike glycoprotein. Sci. Transl. Med. 2014, 6, 234ra59. [Google Scholar] [CrossRef] [PubMed]
- Lowe, D.; Dudgeon, K.; Rouet, R.; Schofield, P.; Jermutus, L.; Christ, D. Aggregation, stability, and formulation of human antibody therapeutics. Adv. Protein Chem. Struct. Biol. 2011, 84, 41–61. [Google Scholar] [PubMed]
- Rouet, R.; Lowe, D.; Christ, D. Stability engineering of the human antibody repertoire. FEBS Lett. 2014, 588, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Chames, P.; Van, R.M.; Weiss, E.; Baty, D. Therapeutic antibodies: Successes, limitations and hopes for the future. Br. J. Pharmacol. 2009, 157, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Nanobodies: Natural single-domain antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [PubMed]
- Siontorou, C.G. Nanobodies as novel agents for disease diagnosis and therapy. Int. J. Nanomed. 2013, 8, 4215–4227. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, J.; Alzogaray, V.; Reyelt, J.; Unger, M.; Juarez, K.; Urrutia, M.; Cauerhff, A.; Danquah, W.; Rissiek, B.; Scheuplein, F.; et al. Single domain antibodies: Promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009, 198, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Hmila, I.; Abdallah, R.B.; Saerens, D.; Benlasfar, Z.; Conrath, K.; Ayeb, M.E.; Muyldermans, S.; Bouhaouala-Zahar, B. VHH, bivalent domains and chimeric Heavy chain-only antibodies with high neutralizing efficacy for scorpion toxin AahI’. Mol. Immunol. 2008, 45, 3847–3856. [Google Scholar] [CrossRef]
- Els, C.K.; Lauwereys, M.; Wyns, L.; Muyldermans, S. Camel single-domain antibodies as modular building units in bispecific and bivalent antibody constructs. J. Biol. Chem. 2001, 276, 7346–7350. [Google Scholar]
- Laursen, N.S.; Friesen, R.H.E.; Zhu, X.; Jongeneelen, M.; Blokland, S.; Vermond, J.; van Eijgen, A.; Tang, C.; van Diepen, H.; Obmolova, G.; et al. Universal protection against influenza infection by a multidomain antibody to influenza hemagglutinin. Science 2018, 362, 598–602. [Google Scholar] [CrossRef]
- Zhao, G.; He, L.; Sun, S.; Qiu, H.; Tai, W.; Chen, J.; Li, J.; Chen, Y.; Guo, Y.; Wang, Y.; et al. A novel nanobody targeting Middle East respiratory syndrome coronavirus (MERS-CoV) receptor-binding domain has potent cross-neutralizing activity and protective efficacy against MERS-CoV. J. Virol. 2018, 92, e00837-18. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.; Zhao, G.; Sun, S.; Guo, Y.; Wang, Y.; Tao, X.; Tseng, C.K.; Li, F.; Jiang, S.; Du, L.; et al. A recombinant receptor-binding domain of MERS-CoV in trimeric form protects human dipeptidyl peptidase 4 (hDPP4) transgenic mice from MERS-CoV infection. Virology 2016, 499, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, W.; He, L.; Wang, Y.; Sun, S.; Zhao, G.; Luo, C.; Li, P.; Zhao, H.; Fremont, D.H.; Li, F.; et al. Critical neutralizing fragment of Zika virus EDIII elicits cross-neutralization and protection against divergent Zika viruses. Emerg. Microbes Infect. 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lu, H.; Siddiqui, P.; Zhou, Y.; Jiang, S. Receptor-binding domain of severe acute respiratory syndrome coronavirus spike protein contains multiple conformation-dependent epitopes that induce highly potent neutralizing antibodies. J. Immunol. 2005, 174, 4908–4915. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tai, W.; Yang, J.; Zhao, G.; Sun, S.; Tseng, C.K.; Jiang, S.; Zhou, Y.; Du, L.; Gao, J. Receptor-binding domain of MERS-CoV with optimal immunogen dosage and immunization interval protects human transgenic mice from MERS-CoV infection. Hum. Vaccin. Immunother. 2017, 13, 1615–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, T.C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Biacchesi, S.; Skiadopoulos, M.H.; Yang, L.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Rapid human metapneumovirus microneutralization assay based on green fluorescent protein expression. J. Virol. Methods 2005, 128, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Gaiotto, T.; Hufton, S.E. Cross-neutralising nanobodies bind to a conserved pocket in the hemagglutinin stem region identified using yeast display and deep mutational scanning. PLoS ONE 2016, 11, e0164296. [Google Scholar] [CrossRef]
- Ardekani, L.S.; Gargari, S.L.; Rasooli, I.; Bazl, M.R.; Mohammadi, M.; Ebrahimizadeh, W.; Bakherad, H.; Zare, H. A novel nanobody against urease activity of Helicobacter pylori. Int. J. Infect. Dis. 2013, 17, e723–e728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussack, G.; Hirama, T.; Ding, W.; Mackenzie, R.; Tanha, J. Engineered single-domain antibodies with high protease resistance and thermal stability. PLoS ONE 2011, 6, e28218. [Google Scholar] [CrossRef] [PubMed]
- AlDeghaither, D.; Smaglo, B.G.; Weiner, L.M. Beyond peptides and mAbs--current status and future perspectives for biotherapeutics with novel constructs. J. Clin. Pharmacol. 2015, S4–S20. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, L.I.; De, F.M.; Hultberg, A.; Verrips, T.; Temperton, N.; Weiss, R.A.; Vandevelde, W.; Schepens, B.; Vanlandschoot, P.; Saelens, X. Nanobodies with in vitro neutralizing activity protect mice against H5N1 influenza virus infection. J. Infect. Dis. 2011, 203, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Bourgeois, J.P.; Celli, S.; Glacial, F.; Le Sourd, A.M.; Mecheri, S.; Weksler, B.; Romero, I.; Couraud, P.O.; Rougeon, F.; et al. Cell-penetrating anti-GFAP VHH and corresponding fluorescent fusion protein VHH-GFP spontaneously cross the blood-brain barrier and specifically recognize astrocytes: Application to brain imaging. FASEB J. 2012, 26, 3969–3979. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Yin, K.; Mi, L.; Ma, M.; Liu, Y.; Li, Y.; Wei, W.; Zhang, Y.; Liu, S. A novel photoelectrochemical immunosensor by integration of nanobody and ZnO nanorods for sensitive detection of nucleoside diphosphatase kinase-A. Anal. Chim. Acta. 2017, 973, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Bannas, P.; Hambach, J.; Koch-Nolte, F. Nanobodies and nanobody-based human heavy chain antibodies as antitumor therapeutics. Front. Immunol. 2017, 8, 1603. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Shi, X.; Jiang, L.; Zhang, S.; Wang, D.; Tong, P.; Guo, D.; Fu, L.; Cui, Y.; Liu, X.; et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res. 2013, 23, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Van Roy, M.; Ververken, C.; Beirnaert, E.; Hoefman, S.; Kolkman, J.; Vierboom, M.; Breedveld, E.; Hart, B.; Poelmans, S.; Bontinck, L.; et al. The preclinical pharmacology of the high affinity anti-IL-6R Nanobody(R) ALX-0061 supports its clinical development in rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 135. [Google Scholar] [CrossRef]
- Raj, V.S.; Okba, N.M.; Gutierrez-Alvarez, J.; Drabek, D.; van Dieren, B.; Widagdo, W.; Lamers, M.M.; Widjaja, I.; Fernandez-Delgado, R.; Sola, I.; et al. Chimeric camel/human heavy-chain antibodies protect against MERS-CoV infection. Sci. Adv. 2018, 4, eaas9667. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Treatment | ND50 (nM, Mean Value) | ||||
|---|---|---|---|---|---|---|
| Mono-Nb | Di-Nb | Tri-Nb | MERS mAb | SARS mAb | ||
| pH | pH 5.0 | 1.94 | 0.21 | 0.03 | 0.53 | ND |
| pH 7.0 | 2.00 | 0.21 | 0.03 | 0.57 | ND | |
| pH 8.0 | 1.93 | 0.21 | 0.03 | 0.53 | ND | |
| No treatment | 2.00 | 0.23 | 0.03 | 0.56 | ND | |
| Pepsin | 0 µg/mL | 2.23 | 0.29 | 0.04 | 0.56 | ND |
| 25 µg/mL | 2.24 | 0.27 | 0.04 | 1.19 ** | ND | |
| 625 µg/mL | 2.20 | 0.26 | 0.04 | 1.86 ** | ND | |
| No treatment | 2.15 | 0.24 | 0.04 | 0.55 | ND | |
| Urea | 0 mM | 2.27 | 0.25 | 0.04 | 0.56 | ND |
| 50 mM | 2.01 | 0.29 | 0.04 | 0.85 ** | ND | |
| 400 mM | 2.18 | 0.25 | 0.03 | 1.46 ** | ND | |
| No treatment | 2.19 | 0.25 | 0.04 | 0.54 | ND | |
| Temperature | 4 °C | 2.06 | 0.25 | 0.03 | 0.56 | ND |
| 37 °C | 1.80 | 0.22 | 0.04 | 1.32 ** | ND | |
| 60 °C | 2.21 | 0.26 | 0.03 | 2.06 *** | ND | |
| No treatment | 2.17 | 0.26 | 0.04 | 0.58 | ND | |
| Accession No. | Isolate Year | Host | Country | S Protein RBD Mutation(s) | ND50 (nM, Mean Value) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mono-Nb | Di-Nb | Tri-Nb | MERS mAb | SARS mAb | |||||
| AFS88936 | 2012 | Human | Saudi Arabia | — | 2.14 | 0.24 ** | 0.03 ** | 0.57 | ND |
| AGV08379 | 2012 | Human | Saudi Arabia | D509G | 3.39 | 0.19 *** | 0.06 *** | 122 | ND |
| AGV08584 | 2012 | Human | Saudi Arabia | V534A | 6.64 | 0.38 * | 0.08 * | 2.02 | ND |
| AFY13307 | 2012 | Human | UK | L506F | 27.1 | 3.29 *** | 0.37 *** | 67.5 | ND |
| AHI48528 | 2013 | Human | Saudi Arabia | A431P, A482V | 1.10 | 0.10 *** | 0.02 *** | 0.52 | ND |
| AHI48733 | 2013 | Human | Saudi Arabia | A434V | 6.67 | 0.11 * | 0.05 * | 2.11 | ND |
| AHC74088 | 2013 | Human | Qatar | S460F | 2.57 | 0.26 ** | 0.04 ** | 0.50 | ND |
| AKM76239 | 2013 | Human | Oman | V514L | 9.07 | 0.95 * | 0.09 ** | 3.65 | ND |
| AID55090 | 2014 | Human | Saudi Arabia | T424I | 0.81 | 0.07 ** | 0.01 ** | 0.80 | ND |
| AID55087 | 2014 | Human | Saudi Arabia | Q522H | 1.49 | 0.09 *** | 0.02 *** | 0.28 | ND |
| ALX27228 | 2014 | Human | Jordan | E536K | 14.7 | 2.96 ** | 0.61 ** | 5.54 | ND |
| ALJ76277 | 2014 | Human | Saudi Arabia | D537E | 7.30 | 1.49 * | 0.41 * | 3.02 | ND |
| ALJ54518 | 2015 | Human | Saudi Arabia | L507P | 13.1 | 2.76 *** | 0.43 *** | 186 | ND |
| ALB08322 | 2015 | Human | South Korea | D510G | 2.28 | 0.23 * | 0.04 ** | 8.92 | ND |
| ALB08289 | 2015 | Human | South Korea | I529T | 3.77 | 0.24 *** | 0.08 *** | 865 | ND |
| ATG84888 | 2016 | Human | Saudi Arabia | S426R | 12.6 | 1.60 ** | 0.34 *** | 15.6 | ND |
| AHY22545 | 2013 | Camel | Saudi Arabia | K400N | 1.77 | 0.20 *** | 0.02 *** | 1.12 | ND |
| AHY22555 | 2013 | Camel | Saudi Arabia | A520S | 1.11 | 0.17 ** | 0.08 ** | 1.10 | ND |
| ASU90076 | 2015 | Camel | UAE | S460T | 6.84 | 1.26 *** | 0.26 *** | 3.36 | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Tai, W.; Li, J.; Chen, Y.; Gao, Y.; Li, J.; Sun, S.; Zhou, Y.; Du, L.; Zhao, G. Enhanced Ability of Oligomeric Nanobodies Targeting MERS Coronavirus Receptor-Binding Domain. Viruses 2019, 11, 166. https://doi.org/10.3390/v11020166
He L, Tai W, Li J, Chen Y, Gao Y, Li J, Sun S, Zhou Y, Du L, Zhao G. Enhanced Ability of Oligomeric Nanobodies Targeting MERS Coronavirus Receptor-Binding Domain. Viruses. 2019; 11(2):166. https://doi.org/10.3390/v11020166
Chicago/Turabian StyleHe, Lei, Wanbo Tai, Jiangfan Li, Yuehong Chen, Yaning Gao, Junfeng Li, Shihui Sun, Yusen Zhou, Lanying Du, and Guangyu Zhao. 2019. "Enhanced Ability of Oligomeric Nanobodies Targeting MERS Coronavirus Receptor-Binding Domain" Viruses 11, no. 2: 166. https://doi.org/10.3390/v11020166
APA StyleHe, L., Tai, W., Li, J., Chen, Y., Gao, Y., Li, J., Sun, S., Zhou, Y., Du, L., & Zhao, G. (2019). Enhanced Ability of Oligomeric Nanobodies Targeting MERS Coronavirus Receptor-Binding Domain. Viruses, 11(2), 166. https://doi.org/10.3390/v11020166