Class A Scavenger Receptors Are Used by Frog Virus 3 During Its Cellular Entry


Abstract
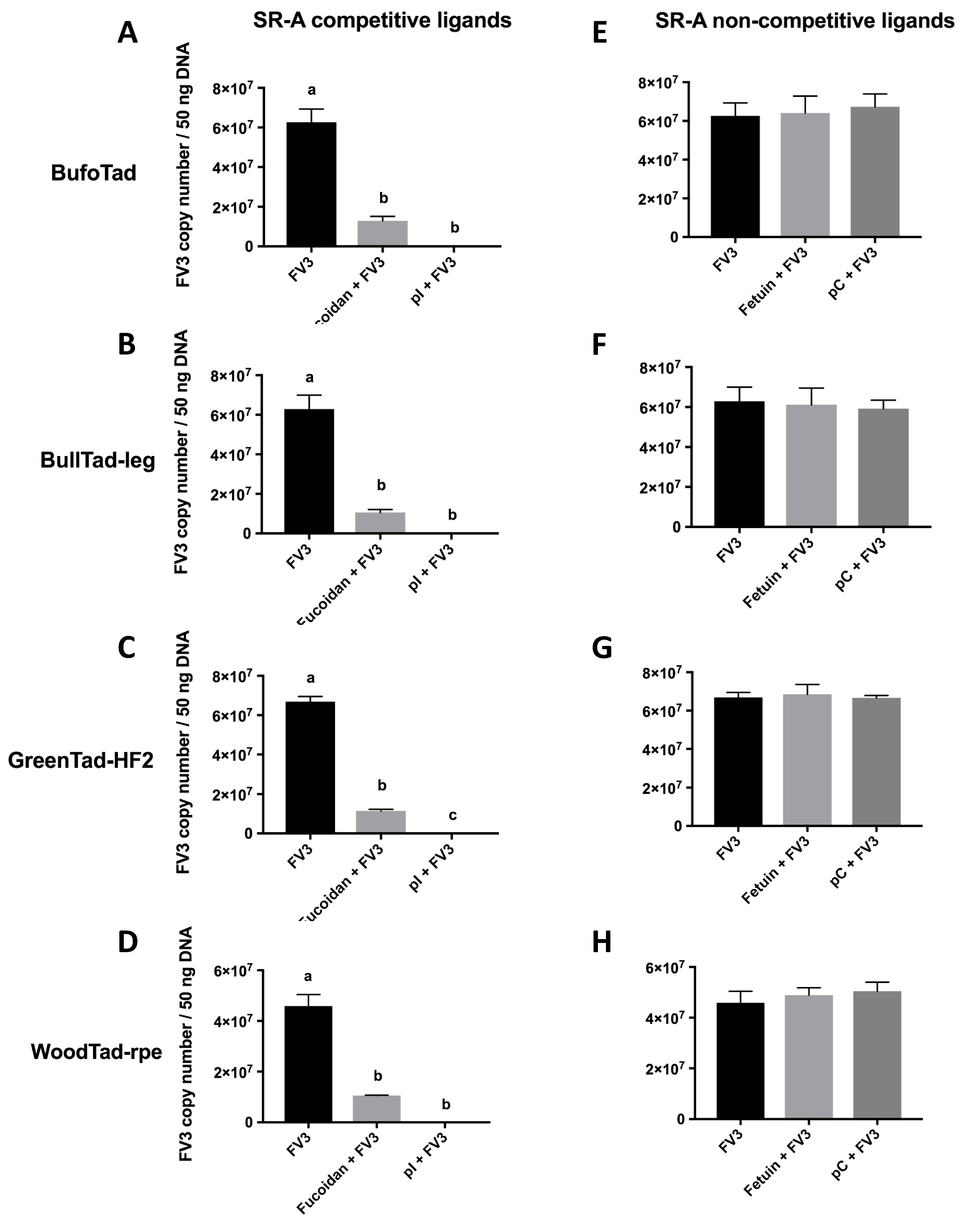
:1. Introduction
2. Materials and Methods
2.1. Cell Culture Media
2.2. Cell Lines and Primary Macrophage Cultures
2.3. Routine Cell Line Maintenance
2.4. FV3 Stocks
2.5. SR-A Ligand Binding Blocking Assay
2.6. Induced Cell-Surface Expression of hSR-AI of hSR-AI-A549 Cells and FV3 Interaction
2.7. Assessment of FV3 Copy Number by Absolute qPCR
2.8. Statistical Analysis
3. Results and Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Price, S.J.; Ariel, E.; Maclaine, A.; Rosa, G.M.; Gray, M.J.; Brunner, J.L.; Garner, T.W.J. From fish to frogs and beyond: Impact and host range of emergent ranaviruses. Virology 2017, 511, 272–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffus, A.L.J.; Waltzek, T.B.; Stöhr, A.C.; Allender, M.C.; Gotesman, M.; Whittington, R.J.; Hick, P.; Hines, M.K.; Marschang, R.E. Distribution and Host Range of Ranaviruses. In Ranaviruses: Lethal Pathogens of Ecothermic Vertebrates; Gray, M.J., Chinchar, V.G., Eds.; Springer: Cham, Switzerland, 2015; pp. 9–57. [Google Scholar] [Green Version]
- Granoff, A.; Came, P.E.; Rafferty, K.A., Jr. The isolation and properties of viruses from Rana pipiens: Their possible relationship to the renal adenocarcinoma of the leopard frog. Ann. N. Y. Acad. Sci. 1965, 126, 237–255. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.L.; Rajeev, S.; Gray, M.J.; Baldwin, C.A. Frog virus 3 infection, cultured American bullfrogs. Emerg. Infect. Dis. 2007, 13, 342–343. [Google Scholar] [CrossRef] [PubMed]
- Grayfer, L.; Robert, J. Divergent antiviral roles of amphibian (Xenopus laevis) macrophages elicited by colony-stimulating factor-1 and interleukin-34. J. Leukoc. Biol. 2014, 96, 1143–1153. [Google Scholar] [CrossRef]
- Grayfer, L.; Robert, J. Distinct functional roles of amphibian (Xenopus laevis) colony-stimulating factor-1- and interleukin-34-derived macrophages. J. Leukoc. Biol. 2015, 98, 641–649. [Google Scholar] [CrossRef]
- Yaparla, A.; Popovic, M.; Grayfer, L. Differentiation-dependent antiviral capacities of amphibian (Xenopus laevis) macrophages. J. Biol. Chem. 2018, 293, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Darlington, R.W.; Granoff, A.; Breeze, D.C. Viruses and renal carcinoma of Rana pipiens: II. Ultrastructural studies and sequential development of virus isolated from normal and tumor tissue. Virology 1966, 29, 149–156. [Google Scholar] [CrossRef]
- Granoff, A.; Came, P.E.; Breeze, D.C. Viruses and renal carcinoma of Rana pipiens. I. The isolation and properties of virus from normal and tumor tissue. Virology 1966, 29, 133–148. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Yu, K.H.; Jancovich, J.K. The molecular biology of frog virus 3 and other iridoviruses infecting cold-blooded vertebrates. Viruses 2011, 3, 1959–1985. [Google Scholar] [CrossRef]
- Braunwald, J.; Tripier, F.; Kirn, A. Comparison of the properties of enveloped and naked frog virus 3 (FV3) particles. J. Gen. Virol. 1979, 45, 673–682. [Google Scholar] [CrossRef]
- Braunwald, J.; Nonnenmacher, H.; Tripier-Darcy, F. Ultrastructural and biochemical study of frog virus 3 uptake by BHK-21 cells. J. Gen. Virol. 1985, 66, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.C. Frog virus 3 replication: Electron microscope observations on the sequence of infection in chick embryo fibroblasts. J. Gen. Virol. 1975, 26, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Houts, G.E.; Gravell, M.; Granoff, A. Electron microscopic observations on early events of frog virus 3 replication. Virology 1974, 58, 589–594. [Google Scholar] [CrossRef]
- Gendrault, J.L.; Steffan, A.M.; Bingen, A.; Kirn, A. Penetration and uncoating of frog virus 3 (FV3) in cultured rat Kupffer cells. Virology 1981, 112, 375–384. [Google Scholar] [CrossRef]
- Eaton, H.E.; Metcalf, J.; Penny, E.; Tcherepanov, V.; Upton, C.; Brunetti, C.R. Comparative genomic analysis of the family Iridoviridae: Re-annotating and defining the core set of iridovirus genes. Virol. J. 2007, 4, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, X.; Wang, S.; Yu, Y.; Ni, S.; Qin, Q. Soft-shelled turtle iridovirus enters cells via cholesterol-dependent, clathrin-mediated endocytosis as well as macropinocytosis. Arch. Virol. 2018, 163, 3023–3033. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huang, X.; Huang, Y.; Hao, X.; Xu, H.; Cai, M.; Wang, H.; Qin, Q. Entry of a novel marine DNA virus, Singapore grouper iridovirus, into host cells occurs via clathrin-mediated endocytosis and macropinocytosis in a pH-dependent manner. J. Virol. 2014, 88, 13047–13063. [Google Scholar] [CrossRef]
- Guo, C.J.; Liu, D.; Wu, Y.Y.; Yang, X.B.; Yang, L.S.; Mi, S.; Huang, Y.X.; Luo, Y.W.; Jia, K.T.; Liu, Z.Y.; et al. Entry of Tiger Frog Virus (an Iridovirus) into HepG2 Cells via a pH-Dependent, Atypical, Caveola-Mediated Endocytosis Pathway. J. Virol. 2011, 85, 6416–6426. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.J.; Wu, Y.Y.; Yang, L.S.; Yang, X.B.; He, J.; Mi, S.; Jia, K.T.; Weng, S.P.; Yu, X.Q.; He, J.G. Infectious spleen and kidney necrosis virus (a fish iridovirus) enters Mandarin fish fry cells via caveola-dependent endocytosis. J. Virol. 2012, 86, 2621–2631. [Google Scholar] [CrossRef]
- Clark, H.F.; Karzon, D.T. Terrapene heart (TH-1), a continuous cell line from the heart of the box turtle Terrapene carolina. Exp. Cell Res. 1967, 48, 263–268. [Google Scholar] [CrossRef]
- Gravell, M.; Granoff, A. Virus and renal adenocarcinoma of Rana pipiens: IX. The influence of temperature and host cell on replication of frog polyhedral cytoplasmic deoxyribovirus (PCDV). Virology 1970, 41, 596–602. [Google Scholar] [CrossRef]
- Morales, H.D.; Abramowitz, L.; Gertz, J.; Sowa, J.; Vogel, A.; Robert, J. Innate immune responses and permissiveness to ranavirus infection of peritoneal leukocytes in the frog Xenopus laevis. J. Virol. 2010, 84, 4912–4922. [Google Scholar] [CrossRef] [PubMed]
- Pham, P.H.; Huang, Y.J.; Mosser, D.D.; Bols, N.C. Use of cell lines and primary cultures to explore the capacity of rainbow trout to be a host for frog virus 3 (FV3). In Vitro Cell. Dev. Biol. Anim. 2015, 51, 894–904. [Google Scholar] [CrossRef]
- Forzán, M.J.; Jones, K.M.; Ariel, E.; Whittington, R.J.; Wood, J.; Markham, R.J.F.; Daoust, P.Y. Pathogenesis of Frog Virus 3 (Ranavirus, Iridoviridae) Infection in Wood Frogs (Rana sylvatica). Vet. Pathol. 2017, 54, 531–548. [Google Scholar] [CrossRef]
- Platt, N.; Gordon, S. Scavenger receptors: Diverse activities and promiscuous binding of polyanionic ligands. Chem. Biol. 1998, 5, R193–R203. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.J.; Collins, S.E.; Bauer, C.M.; Bowdish, D.M.; Mossman, K.L. An accessory to the ‘Trinity’: SR-As are essential pathogen sensors of extracellular dsRNA, mediating entry and leading to subsequent type I IFN responses. PLoS Pathog. 2010, 6, e1000829. [Google Scholar] [CrossRef]
- Baid, K.; Nellimarla, S.; Huynh, A.; Boulton, S.; Guarné, A.; Melacini, G.; Collins, S.E.; Mossman, K.L. Direct binding and internalization of diverse extracellular nucleic acid species through the collagenous domain of class A scavenger receptors. Immunol. Cell Biol. 2018, 96, 922–934. [Google Scholar] [CrossRef]
- Han, H.J.; Tokino, T.; Nakamura, Y. CSR, a scavenger receptor-like protein with a protective role against cellular damage caused by UV irradiation and oxidative stress. Hum. Mol. Genet. 1998, 7, 1039–1046. [Google Scholar] [CrossRef]
- Krieger, M. The other side of scavenger receptors: Pattern recognition for host defense. Curr. Opin. Lipidol. 1997, 8, 275–280. [Google Scholar] [CrossRef]
- Nakamura, K.; Funakoshi, H.; Miyamoto, K.; Tokunaga, F.; Nakamura, T. Molecular cloning and functional characterization of a human scavenger receptor with C-type lectin (SRCL), a novel member of a scavenger receptor family. Biochem. Biophys. Res. Commun. 2001, 280, 1028–1035. [Google Scholar] [CrossRef]
- Jiang, Y.; Oliver, P.; Davies, K.E.; Platt, N. Identification and characterization of murine SCARA5, a novel class A scavenger receptor that is expressed by populations of epithelial Cells. J. Biol. Chem. 2006, 281, 11834–11845. [Google Scholar] [CrossRef] [PubMed]
- Whelan, F.J.; Meehan, C.J.; Golding, G.B.; McConkey, B.J.; Bowdish, D.M. The evolution of the class A scavenger receptors. BMC Evol. Biol. 2012, 12, 227. [Google Scholar] [CrossRef] [PubMed]
- Poynter, S.J.; Weleff, J.; Soares, A.B.; DeWitte-Orr, S.J. Class-A scavenger receptor function and expression in the rainbow trout (Oncorhynchus mykiss) epithelial cell lines RTgutGC and RTgill-W1. Fish Shellfish Immunol. 2015, 44, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Vo, N.T.K.; Moore, L.C.; Leis, E.; DeWitte-Orr, S.J. Class A scavenger receptors mediate extracellular dsRNA sensing, leading to downstream antiviral gene expression in a novel American toad cell line, BufoTad. Dev. Comp. Immunol. 2019, 92, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Monjo, A.L.; Poynter, S.J.; DeWitte-Orr, S.J. CHSE-214: A model for studying extracellular dsRNA sensing in vitro. Fish Shellfish Immunol. 2017, 68, 266–271. [Google Scholar] [CrossRef]
- Semple, S.L.; Vo, N.T.K.; Poynter, S.J.; Li, M.; Heath, D.D.; DeWitte-Orr, S.J.; Dixon, B. Extracellular dsRNA induces a type I interferon response in a novel Chinook salmon cell line. Dev. Comp. Immunol. 2018, 89, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Hirano, S.; Kanno, S. Macrophage Receptor with Collagenous Structure (MARCO) is processed by either macropinocytosis or endocytosis-autophagy pathway. PLoS ONE 2015, 10, e0142062. [Google Scholar] [CrossRef] [PubMed]
- Lunov, O.; Zablotskii, V.; Syrovets, T.; Röcker, C.; Tron, K.; Nienhaus, G.U.; Simmet, T. Modeling receptor-mediated endocytosis of polymer-functionalized iron oxide nanoparticles by human macrophages. Biomaterials 2011, 32, 547–555. [Google Scholar] [CrossRef]
- Zhu, X.D.; Zhuang, Y.; Ben, J.J.; Qian, L.L.; Huang, H.P.; Bai, H.; Sha, J.H.; He, Z.G.; Chen, Q. Caveolae-dependent endocytosis is required for class A macrophage scavenger receptor-mediated apoptosis in macrophages. J. Biol. Chem. 2011, 286, 8231–8239. [Google Scholar] [CrossRef]
- MacLeod, D.T.; Nakatsuji, T.; Yamasaki, K.; Kobzik, L.; Gallo, R.L. HSV-1 exploits the innate immune scavenger receptor MARCO to enhance epithelial adsorption and infection. Nat. Commun. 2013, 4, 1963. [Google Scholar] [CrossRef] [Green Version]
- MacLeod, D.T.; Nakatsuji, T.; Wang, Z.; di Nardo, A.; Gallo, R.L. Vaccinia virus binds to the scavenger receptor MARCO on the surface of keratinocytes. J. Investig. Dermatol. 2015, 135, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Haisma, H.J.; Boesjes, M.; Beerens, A.M.; van der Strate, B.W.; Curiel, D.T.; Plüddemann, A.; Gordon, S.; Bellu, A.R. Scavenger receptor A: A new route for adenovirus 5. Mol. Pharm. 2009, 6, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Stichling, N.; Suomalainen, M.; Flatt, J.W.; Schmid, M.; Pacesa, M.; Hemmi, S.; Jungraithmayr, W.; Maler, M.D.; Freudenberg, M.A.; Plückthun, A.; et al. Lung macrophage scavenger receptor SR-A6 (MARCO) is an adenovirus type-specific virus entry receptor. PLoS Pathog. 2018, 14, e1006914. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, R.; Montenegro-Miranda, P.S.; Riviere, C.; Schilderink, R.; ten Bloemendaal, L.; van Gorp, J.; Duijst, S.; de Waart, D.R.; Beuers, U.; Haisma, H.J.; et al. Polyinosinic acid blocks adeno-associated virus macrophage endocytosis in vitro and enhances adeno-associated virus liver-directed gene therapy in vivo. Hum. Gene Ther. 2013, 24, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.; Abramowitz, L.; Gantress, J.; Morales, H.D. Xenopus laevis: A possible vector of ranavirus infection? J. Wildl. Dis. 2007, 43, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Yaparla, A.; Wendel, E.S.; Grayfer, L. The unique myelopoiesis strategy of the amphibian Xenopus laevis. Dev. Comp. Immunol. 2016, 63, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Lisser, G.J.; Vo, N.T.K.; DeWitte-Orr, S.J. Delineating the roles of cellular and innate antiviral immune parameters mediating ranavirus susceptibility using rainbow trout cell lines. Virus Res. 2017, 238, 114–123. [Google Scholar] [CrossRef]
- Grayfer, L.; Andino, F.D.J.; Robert, J. The amphibian (Xenopus laevis) type I interferon response to frog virus 3: New insight into ranavirus pathogenicity. J. Virol. 2014, 88, 5766–5777. [Google Scholar] [CrossRef]
- Tweedell, K.; Granoff, A. Viruses and renal carcinoma of Rana pipiens. V. Effect of frog virus 3 on developing frog embryos and larvae. J. Natl. Cancer Inst. 1967, 40, 407–409. [Google Scholar]
- Vo, N.T.K.; Everson, J.; Moore, L.C.; DeWitte-Orr, S.J. Class A scavenger receptor expression and function in eight novel tadpole cell lines from the green frog (Lithobates clamitans) and the wood frog (Lithobates sylvatica). Cytotechnology 2019. submitted. [Google Scholar]
- Zeng, X.T.; Gao, X.C.; Zhang, Q.Y. Rana grylio virus 43R encodes an envelope protein involved in virus entry. Virus Genes 2018, 54, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Chinchar, V.G.; Zhang, Q.Y. Molecular basis of pathogenesis of emerging viruses infecting aquatic animals. Aquacult. Fish. 2018, 3, 1–5. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Gui, J.F. Virus genomes and virus-host interactions in aquaculture animals. Sci. China Life Sci. 2015, 58, 156–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Cell Culture Type | Species | Developmental Stage | Cell Morphology | FV3 Isolate for Infection |
|---|---|---|---|---|---|
| BufoTad | Cell line | American toad (Anaxyrus americanus) | Tadpole (Gosner stage 35-38) | Endothelial-like | EPC-derived isolate |
| BullTad-leg | Cell line | Bullfrog (Lithobates catesbeianus) | Tadpole (Gosner stage 40-41) | Fibroblastic | EPC-derived isolate |
| GreenTad-HF2 | Cell line | Green frog (Lithobates clamitans) | Tadpole (Gosner stage 25-27) | Fibroblastic | EPC-derived isolate |
| WoodTad-rpe | Cell line | Wood frog (Lithobates sylvaticus) | Tadpole (Gosner stage 43-45) | Epithelial | EPC-derived isolate |
| Bone marrow rXlCSF-1-derived macrophages (Mϕs) | Primary culture | African clawed frog (Xenopus laevis) | Adult | Macrophage | Baby hamster kidney (BHK-21)-derived isolate |
| Bone marrow rXlIL-34-derived Mϕs | Primary culture | African clawed frog (Xenopus laevis) | Adult | Macrophage | BHK-21-derived isolate |
| Class A scavenger receptor (SR-A)-null variant A549 cell line with an inducible human SR-AI | Cell line | Human (homo sapiens) | n/a | Epithelial | EPC-derived isolate |
| Cell Cultures | SR-A Gene Expression | ||||
|---|---|---|---|---|---|
| SR-AI | MARCO | SCARA3 | SCARA4 | SCARA5 | |
| BufoTad | positive | negative | unknown | unknown | unknown |
| BullTad-leg | positive | negative | positive | positive | positive |
| GreenTad-HF2 | positive | negative | positive | positive | positive |
| WoodTad-rpe | negative | negative | positive | positive | positive |
| X. laevis bone marrow rXlCSF-1-derived Mϕs | negative | positive | positive | positive | negative |
| X. laevis bone marrow rXlIL-34-derived Mϕs | negative | positive | positive | positive | negative |
| SR-A-null variant A549 cell line with an inducible human SR-AI | negative but inducible | negative | negative | negative | negative |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vo, N.T.K.; Guerreiro, M.; Yaparla, A.; Grayfer, L.; DeWitte-Orr, S.J. Class A Scavenger Receptors Are Used by Frog Virus 3 During Its Cellular Entry. Viruses 2019, 11, 93. https://doi.org/10.3390/v11020093
Vo NTK, Guerreiro M, Yaparla A, Grayfer L, DeWitte-Orr SJ. Class A Scavenger Receptors Are Used by Frog Virus 3 During Its Cellular Entry. Viruses. 2019; 11(2):93. https://doi.org/10.3390/v11020093
Chicago/Turabian StyleVo, Nguyen T. K., Matthew Guerreiro, Amulya Yaparla, Leon Grayfer, and Stephanie J. DeWitte-Orr. 2019. "Class A Scavenger Receptors Are Used by Frog Virus 3 During Its Cellular Entry" Viruses 11, no. 2: 93. https://doi.org/10.3390/v11020093
APA StyleVo, N. T. K., Guerreiro, M., Yaparla, A., Grayfer, L., & DeWitte-Orr, S. J. (2019). Class A Scavenger Receptors Are Used by Frog Virus 3 During Its Cellular Entry. Viruses, 11(2), 93. https://doi.org/10.3390/v11020093