High-Pressure Response of Amyloid Folds


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Sensitivity to Pressure of the α-Helical recPrP Fold
3. Sensitivity to Pressure of the β-Sheet-Rich recPrP Assemblies
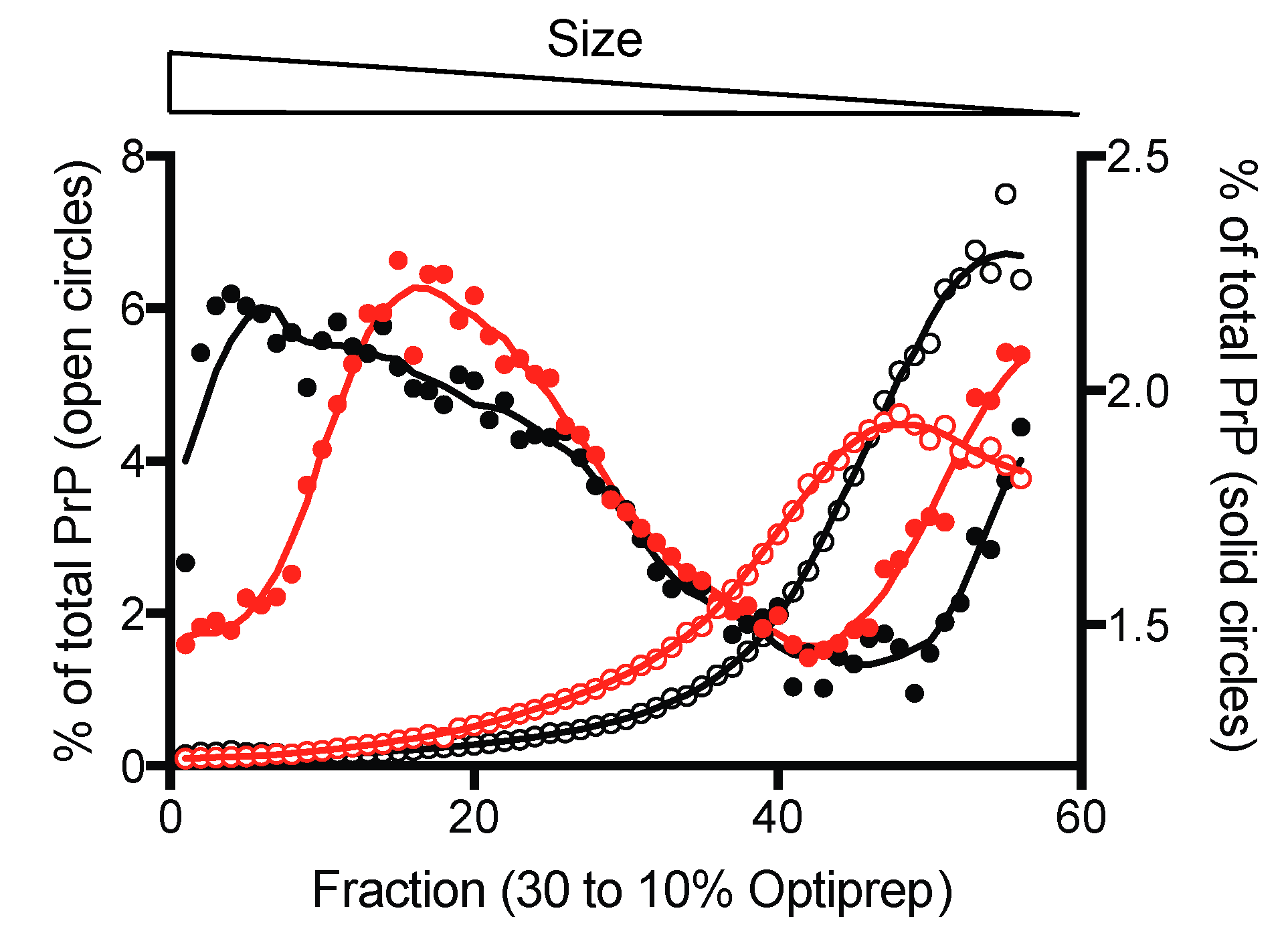
3.1. Oligomers
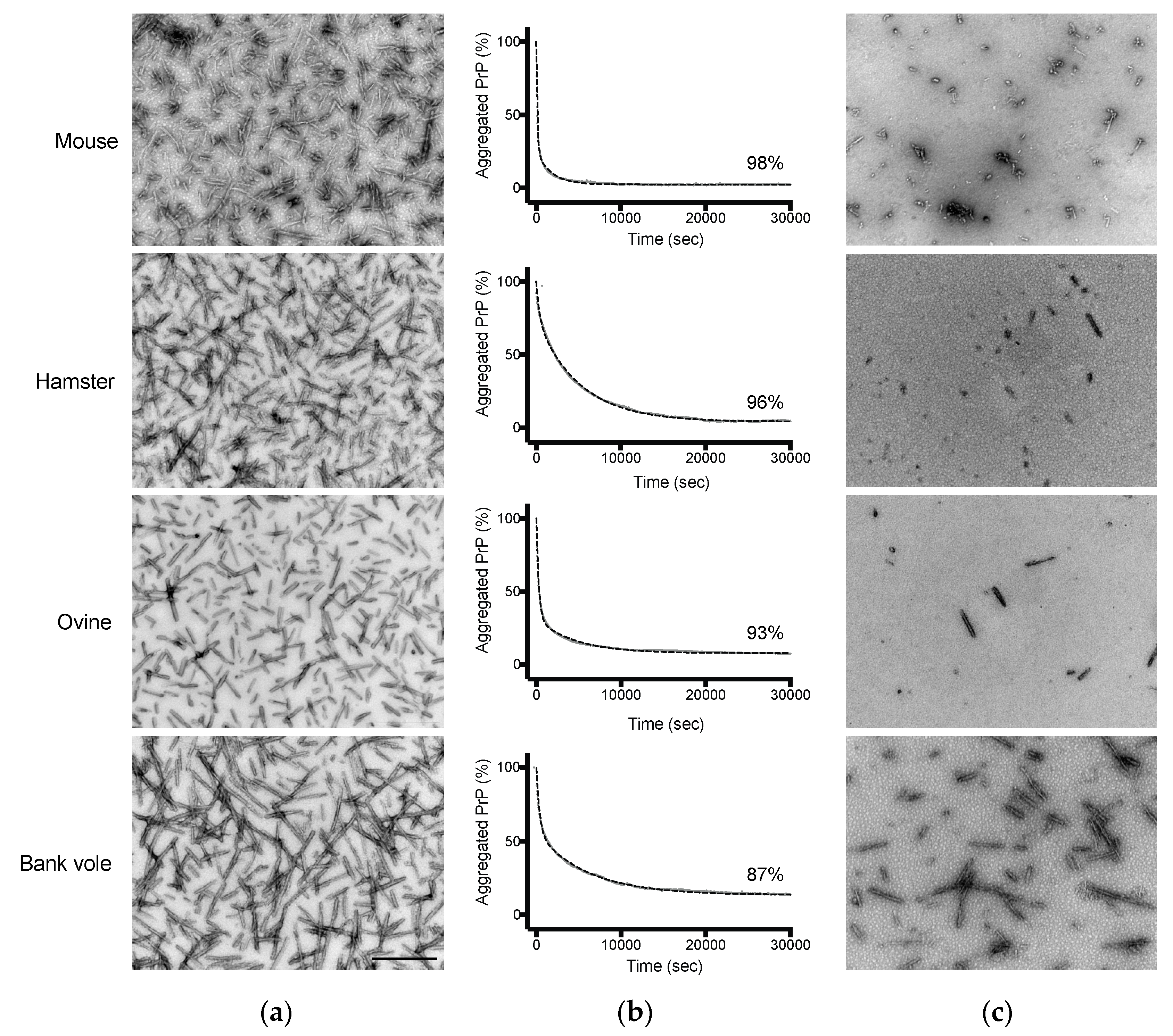
3.2. Amyloid Fibrils
4. Outstanding Questions
5. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jahn, T.R.; Radford, S.E. Folding versus aggregation: Polypeptide conformations on competing pathways. Arch. Biochem. Biophys. 2008, 469, 100–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, J.W. Towards an understanding of amyloidogenesis. Nat. Struct. Biol. 2002, 9, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Soto, C.; Pritzkow, S. Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Annamalai, K.; Guhrs, K.H.; Koehler, R.; Schmidt, M.; Michel, H.; Loos, C.; Gaffney, P.M.; Sigurdson, C.J.; Hegenbart, U.; Schonland, S.; Fandrich, M. Polymorphism of Amyloid Fibrils In Vivo. Angew. Chem. Int. Ed. Engl. 2016, 55, 4822–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legname, G.; Nguyen, H.O.; Peretz, D.; Cohen, F.E.; DeArmond, S.J.; Prusiner, S.B. Continuum of prion protein structures enciphers a multitude of prion isolate-specified phenotypes. Proc. Natl. Acad. Sci. USA 2006, 103, 19105–19110. [Google Scholar] [CrossRef] [PubMed]
- Safar, J.; Wille, H.; Itri, V.; Groth, D.; Serban, H.; Torchia, M.; Cohen, F.E.; Prusiner, S.B. Eight prion strains have PrP(Sc) molecules with different conformations. Nat. Med. 1998, 4, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Telling, G.C.; Parchi, P.; DeArmond, S.J.; Cortelli, P.; Montagna, P.; Gabizon, R.; Mastrianni, J.; Lugaresi, E.; Gambetti, P.; Prusiner, S.B. Evidence for the conformation of the pathologic isoform of the prion protein enciphering and propagating prion diversity. Science 1996, 274, 2079–2082. [Google Scholar] [CrossRef] [PubMed]
- Jucker, M.; Walker, L.C. Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature 2013, 501, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazit, E. The “Correctly Folded” state of proteins: Is it a metastable state? Angew. Chem. Int. Ed. Engl. 2002, 41, 257–259. [Google Scholar] [CrossRef]
- Close, W.; Neumann, M.; Schmidt, A.; Hora, M.; Annamalai, K.; Schmidt, M.; Reif, B.; Schmidt, V.; Grigorieff, N.; Fandrich, M. Physical basis of amyloid fibril polymorphism. Nat. Commun. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Igel-Egalon, A.; Moudjou, M.; Martin, D.; Busley, A.; Knapple, T.; Herzog, L.; Reine, F.; Lepejova, N.; Richard, C.A.; Beringue, V.; Rezaei, H. Reversible unfolding of infectious prion assemblies reveals the existence of an oligomeric elementary brick. PLoS Pathog. 2017, 13, e1006557. [Google Scholar] [CrossRef] [PubMed]
- Balny, C.; Masson, P.; Heremans, K. High pressure effects on biological macromolecules: From structural changes to alteration of cellular processes. Biochim. Biophys. Acta. 2002, 1595, 3–10. [Google Scholar] [CrossRef]
- Heremans, K. High pressure effects on proteins and other biomolecules. Annu. Rev. Biophys. Bioeng. 1982, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Randolph, T.W.; Seefeldt, M.; Carpenter, J.F. High hydrostatic pressure as a tool to study protein aggregation and amyloidosis. Biochim. Biophys. Acta 2002, 1595, 224–234. [Google Scholar] [CrossRef]
- Silva, J.L.; Foguel, D.; Royer, C.A. Pressure provides new insights into protein folding, dynamics and structure. Trends Biochem. Sci. 2001, 26, 612–618. [Google Scholar] [CrossRef]
- Winter, R.; Dzwolak, W. Exploring the temperature-pressure configurational landscape of biomolecules: From lipid membranes to proteins. Philos. Trans. A Math. Phys. Eng. Sci. 2005, 363, 537–563. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.; Caro, J.A.; Norberto, D.R.; Barthe, P.; Roumestand, C.; Schlessman, J.L.; Garcia, A.E.; Garcia-Moreno, B.E.; Royer, C.A. Cavities determine the pressure unfolding of proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 6945–6950. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, G.A.; Marques, M.A.; Cruzeiro-Silva, C.; Cordeiro, Y.; Schuabb, C.; Moraes, A.H.; Winter, R.; Oschkinat, H.; Foguel, D.; Freitas, M.S.; et al. Structural basis for the dissociation of alpha-synuclein fibrils triggered by pressure perturbation of the hydrophobic core. Sci. Rep. 2016, 6, 37990. [Google Scholar] [CrossRef] [PubMed]
- El Moustaine, D.; Perrier, V.; Acquatella-Tran Van Ba, I.; Meersman, F.; Ostapchenko, V.G.; Baskakov, I.V.; Lange, R.; Torrent, J. Amyloid features and neuronal toxicity of mature prion fibrils are highly sensitive to high pressure. J. Biol. Chem. 2011, 286, 13448–13459. [Google Scholar] [CrossRef] [PubMed]
- Ferrao-Gonzales, A.D.; Palmieri, L.; Valory, M.; Silva, J.L.; Lashuel, H.; Kelly, J.W.; Foguel, D. Hydration and packing are crucial to amyloidogenesis as revealed by pressure studies on transthyretin variants that either protect or worsen amyloid disease. J. Mol. Biol. 2003, 328, 963–974. [Google Scholar] [CrossRef]
- Foguel, D.; Suarez, M.C.; Ferrao-Gonzales, A.D.; Porto, T.C.; Palmieri, L.; Einsiedler, C.M.; Andrade, L.R.; Lashuel, H.A.; Lansbury, P.T.; Kelly, J.W.; Silva, J.L. Dissociation of amyloid fibrils of alpha-synuclein and transthyretin by pressure reveals their reversible nature and the formation of water-excluded cavities. Proc. Natl. Acad. Sci. USA 2003, 100, 9831–9836. [Google Scholar] [CrossRef] [PubMed]
- Rezaei-Ghaleh, N.; Amininasab, M.; Kumar, S.; Walter, J.; Zweckstetter, M. Phosphorylation modifies the molecular stability of beta-amyloid deposits. Nat. Commun. 2016, 7, 11359. [Google Scholar] [CrossRef] [PubMed]
- Torrent, J.; Lange, R.; Rezaei, H. The Volumetric Diversity of Misfolded Prion Protein Oligomers Revealed by Pressure Dissociation. J. Biol. Chem. 2015, 290, 20417–20426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrent, J.; Alvarez-Martinez, M.T.; Heitz, F.; Liautard, J.P.; Balny, C.; Lange, R. Alternative prion structural changes revealed by high pressure. Biochemistry 2003, 42, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Makarava, N.; Savtchenko, R.; Baskakov, I.V. Purification and Fibrillation of Full-Length Recombinant PrP. Methods Mol. Biol. 2017, 1658, 3–22. [Google Scholar] [PubMed]
- Eghiaian, F.; Daubenfeld, T.; Quenet, Y.; van Audenhaege, M.; Bouin, A.P.; van der Rest, G.; Grosclaude, J.; Rezaei, H. Diversity in prion protein oligomerization pathways results from domain expansion as revealed by hydrogen/deuterium exchange and disulfide linkage. Proc. Natl. Acad. Sci. USA 2007, 104, 7414–7419. [Google Scholar] [CrossRef] [PubMed]
- Armiento, A.; Moireau, P.; Martin, D.; Lepejova, N.; Doumic, M.; Rezaei, H. The mechanism of monomer transfer between two structurally distinct PrP oligomers. PLoS ONE 2017, 12, e0180538. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.M.; Surewicz, W.K. Fibril conformation as the basis of species- and strain-dependent seeding specificity of mammalian prion amyloids. Cell 2005, 121, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Tixador, P.; Herzog, L.; Reine, F.; Jaumain, E.; Chapuis, J.; Le Dur, A.; Laude, H.; Beringue, V. The physical relationship between infectivity and prion protein aggregates is strain-dependent. PLoS Pathog. 2010, 6, e1000859. [Google Scholar] [CrossRef] [PubMed]
- Laferriere, F.; Tixador, P.; Moudjou, M.; Chapuis, J.; Sibille, P.; Herzog, L.; Reine, F.; Jaumain, E.; Laude, H.; Rezaei, H.; Beringue, V. Quaternary structure of pathological prion protein as a determining factor of strain-specific prion replication dynamics. PLoS Pathog. 2013, 9, e1003702. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mahal, S.P.; Demczyk, C.A.; Weissmann, C. Mutability of prions. EMBO Rep. 2011, 12, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, J.; Mahler, J.; Beschorner, N.; Kaeser, S.A.; Hasler, L.M.; Baumann, F.; Nystrom, S.; Portelius, E.; Blennow, K.; Lashley, T.; et al. Amyloid polymorphisms constitute distinct clouds of conformational variants in different etiological subtypes of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2017, 114, 13018–13023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breydo, L.; Makarava, N.; Baskakov, I.V. Methods for conversion of prion protein into amyloid fibrils. Methods Mol. Biol. 2008, 459, 105–115. [Google Scholar] [PubMed]
- Torrent, J.; Font, J.; Herberhold, H.; Marchal, S.; Ribo, M.; Ruan, K.; Winter, R.; Vilanova, M.; Lange, R. The use of pressure-jump relaxation kinetics to study protein folding landscapes. Biochim. Biophys. Acta 2006, 1764, 489–496. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torrent, J.; Martin, D.; Igel-Egalon, A.; Béringue, V.; Rezaei, H. High-Pressure Response of Amyloid Folds. Viruses 2019, 11, 202. https://doi.org/10.3390/v11030202
Torrent J, Martin D, Igel-Egalon A, Béringue V, Rezaei H. High-Pressure Response of Amyloid Folds. Viruses. 2019; 11(3):202. https://doi.org/10.3390/v11030202
Chicago/Turabian StyleTorrent, Joan, Davy Martin, Angélique Igel-Egalon, Vincent Béringue, and Human Rezaei. 2019. "High-Pressure Response of Amyloid Folds" Viruses 11, no. 3: 202. https://doi.org/10.3390/v11030202
APA StyleTorrent, J., Martin, D., Igel-Egalon, A., Béringue, V., & Rezaei, H. (2019). High-Pressure Response of Amyloid Folds. Viruses, 11(3), 202. https://doi.org/10.3390/v11030202